

Gallic Acid Ameliorates the Inflammatory State of Periodontal Ligament Stem Cells and Promotes Pro-Osteodifferentiation Capabilities of Inflammatory Stem Cell-Derived Exosomes

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Methods
2.2.1. Primary Cell Extraction and Culture of PDLSCs
2.2.2. GA Induction Culture
2.2.3. MTT Cytotoxicity Assay
2.2.4. mRNA Sequencing
2.2.5. Quantification of DEG-Related Proteins by ELISA
2.2.6. Detection of Mitochondrial Membrane Potential
2.2.7. Detection of Oxidative Stress Level
2.2.8. Detection of Glycometabolism Level
2.2.9. Detection of Osteodifferentiation Level
2.2.10. Extraction of Exosomes by Differential Centrifugation
2.2.11. Exosomes Identification
2.2.12. Coculturing of i-PDLSCs and Exosomes
2.2.13. Detection of Osteodifferentiation Index
2.2.14. Statistical Analysis
3. Results
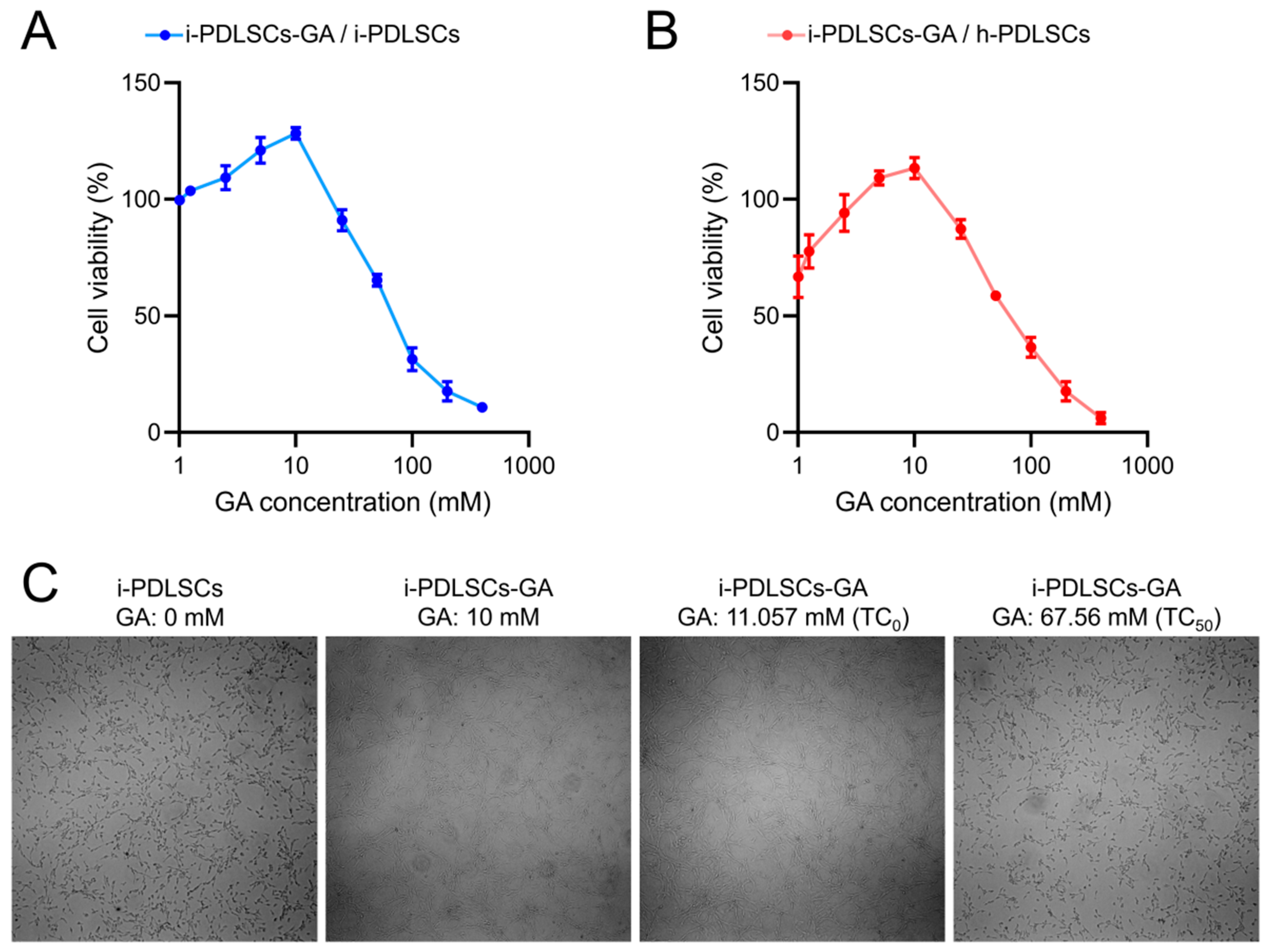
3.1. GA Promotes the Proliferation Activity of i-PDLSCs
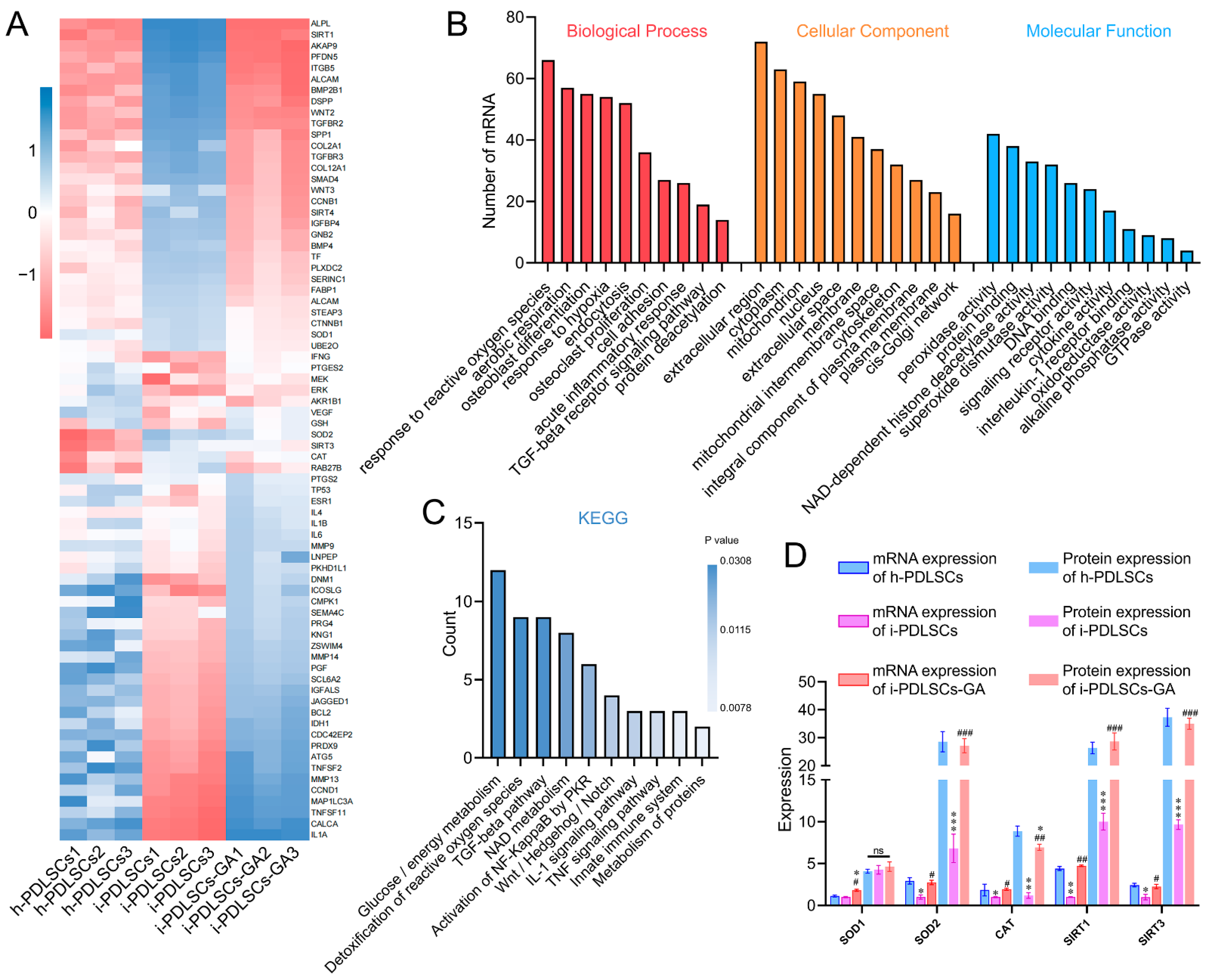
3.2. mRNA Sequencing Showed That the GA-Induced Culture Improved Oxidative Stress and Glycometabolism of i-PDLSCs
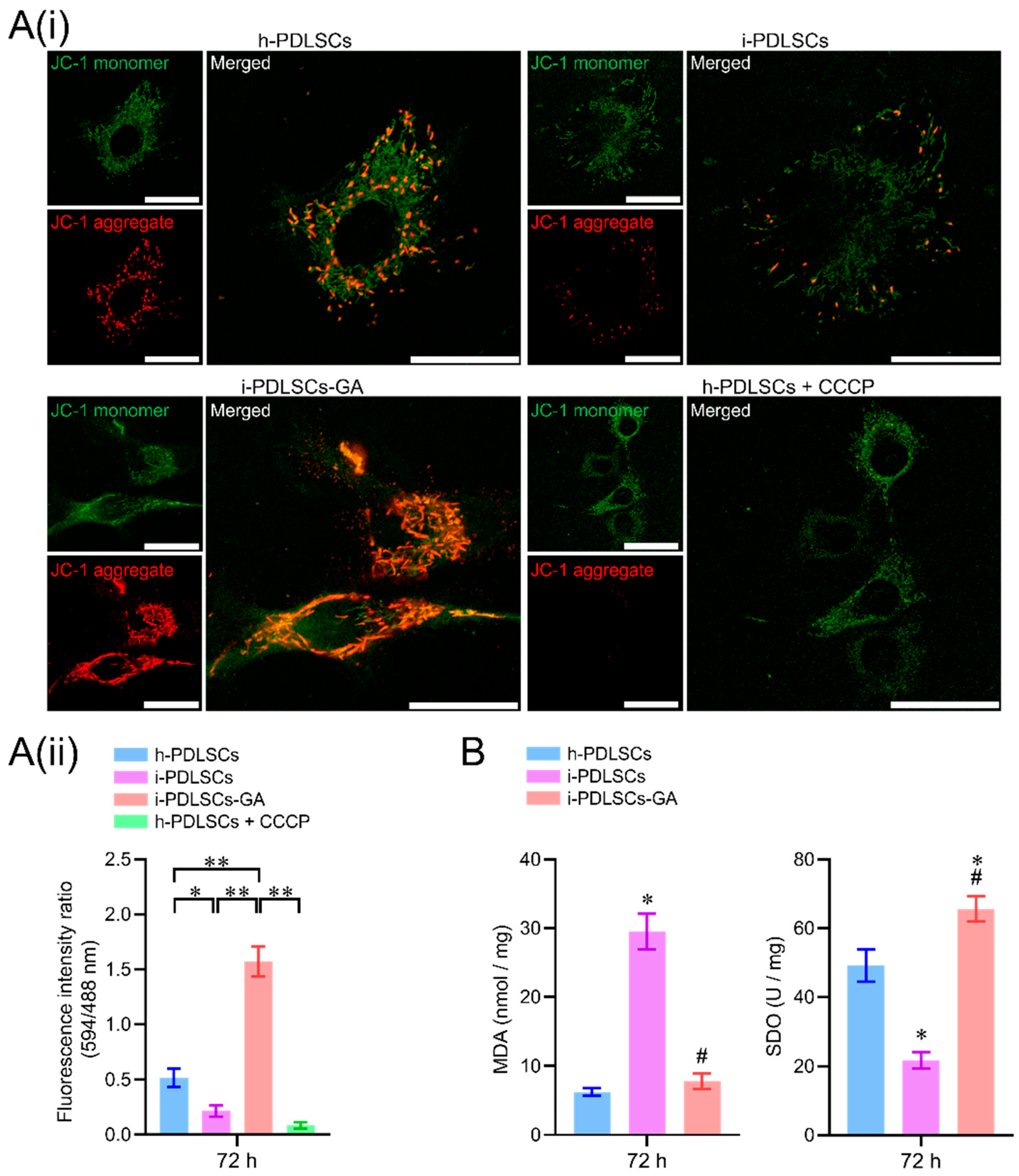
3.3. GA Treatment Alleviates Oxidative Stress in i-PDLSCs
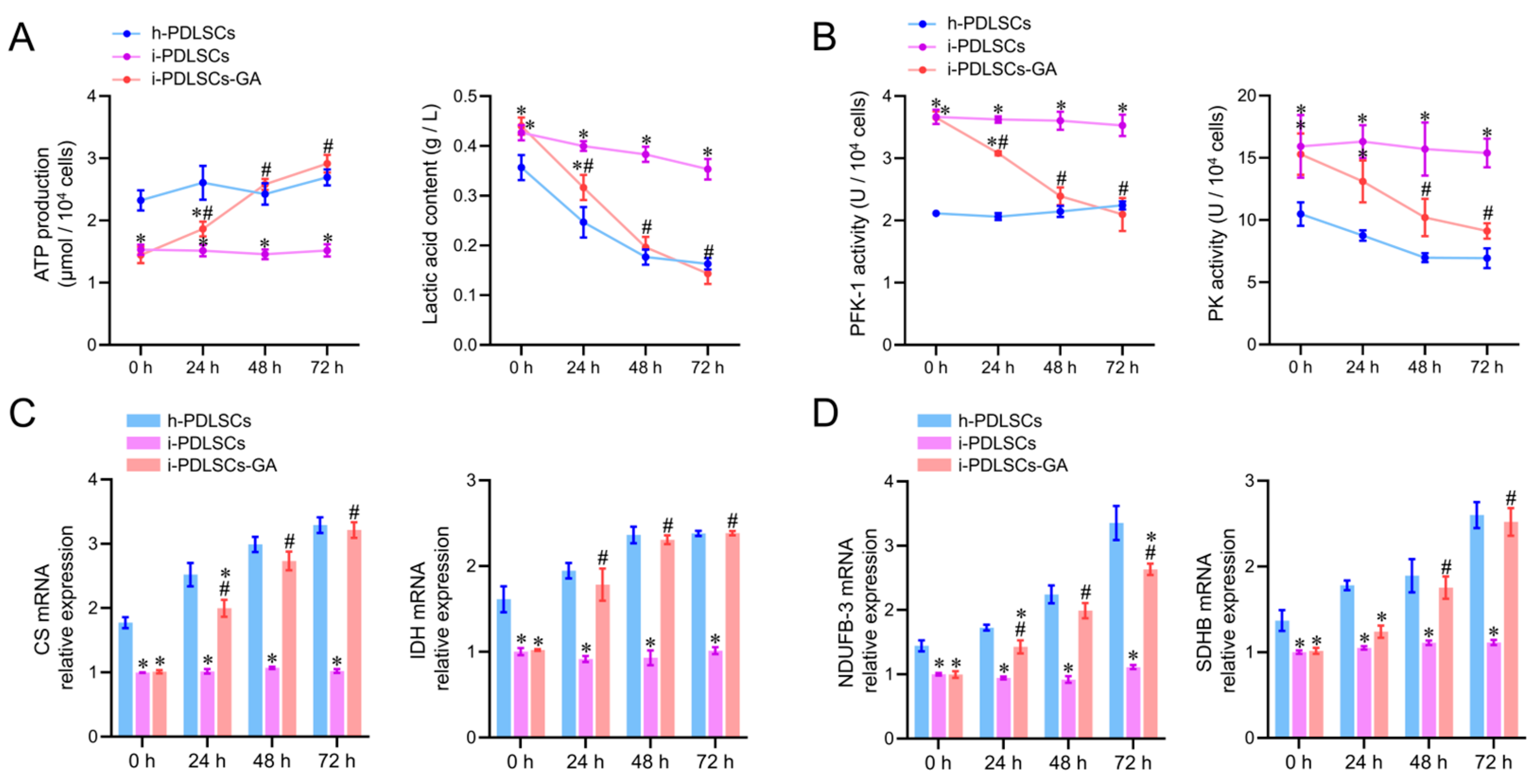
3.4. Enhancement of Glycometabolism in i-PDLSCs Induced by GA
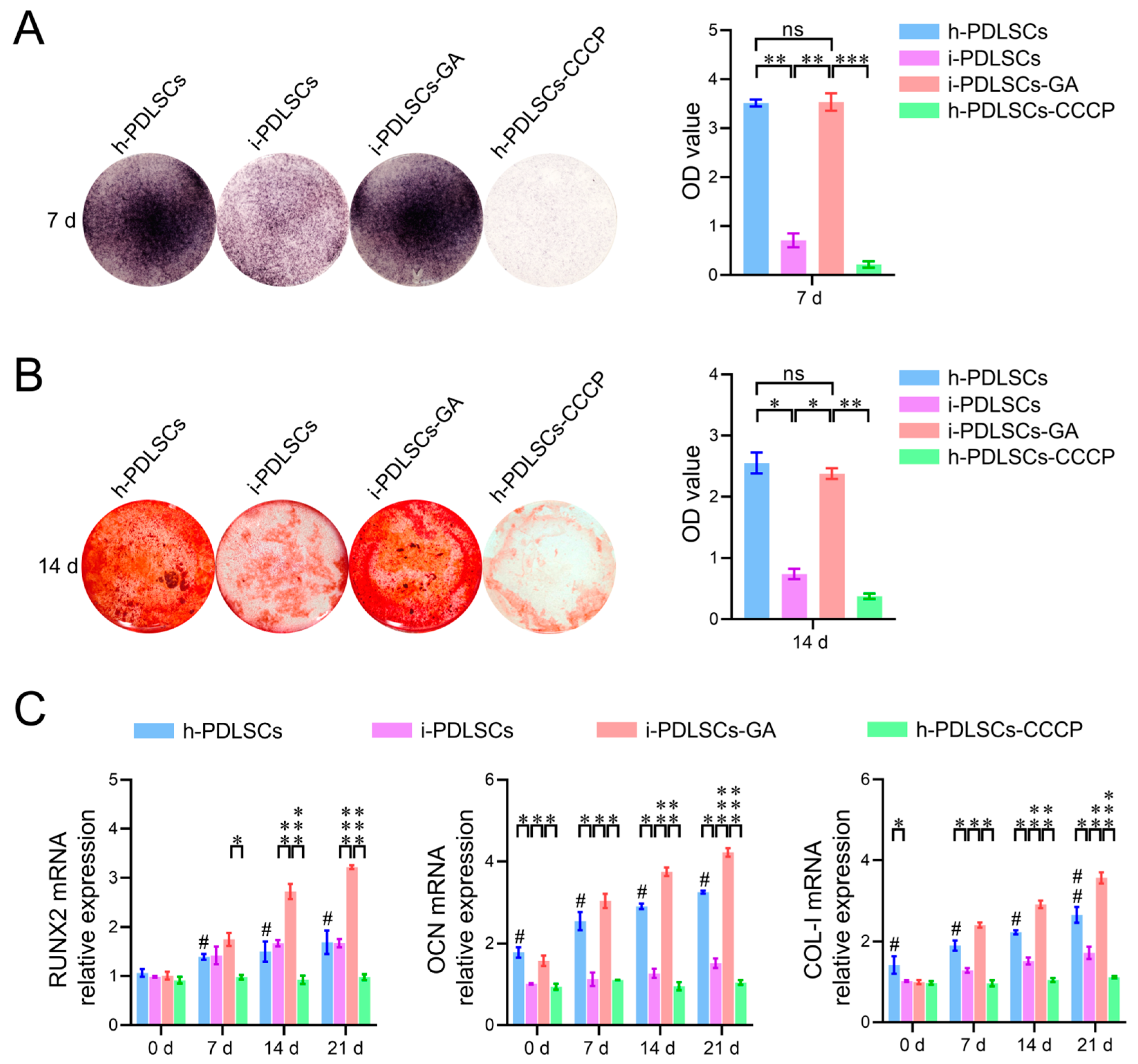
3.5. Effect of Oxidative Stress and Aerobic Glucose Metabolism on Osteodifferentiation of i-PDLSCs
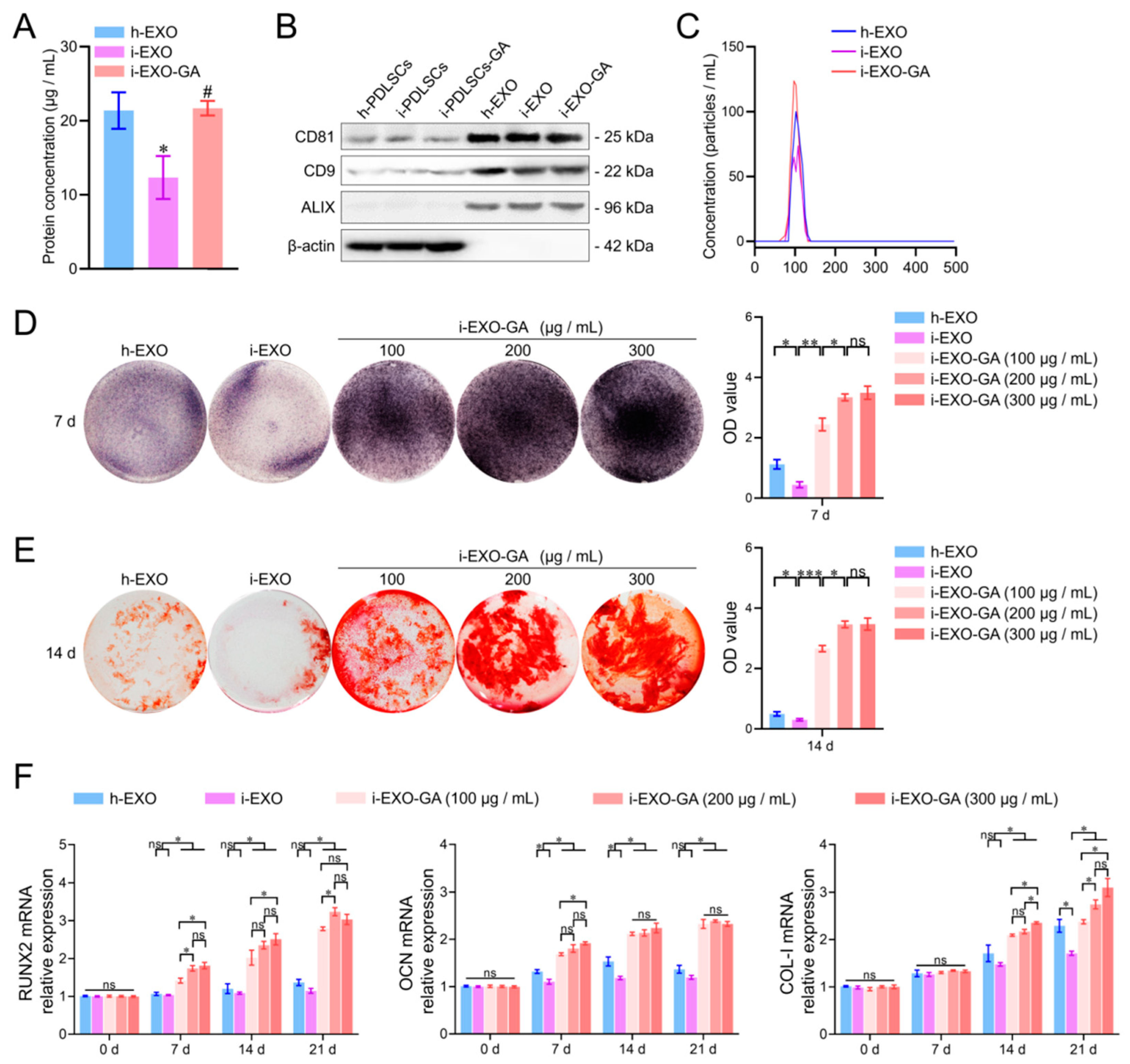
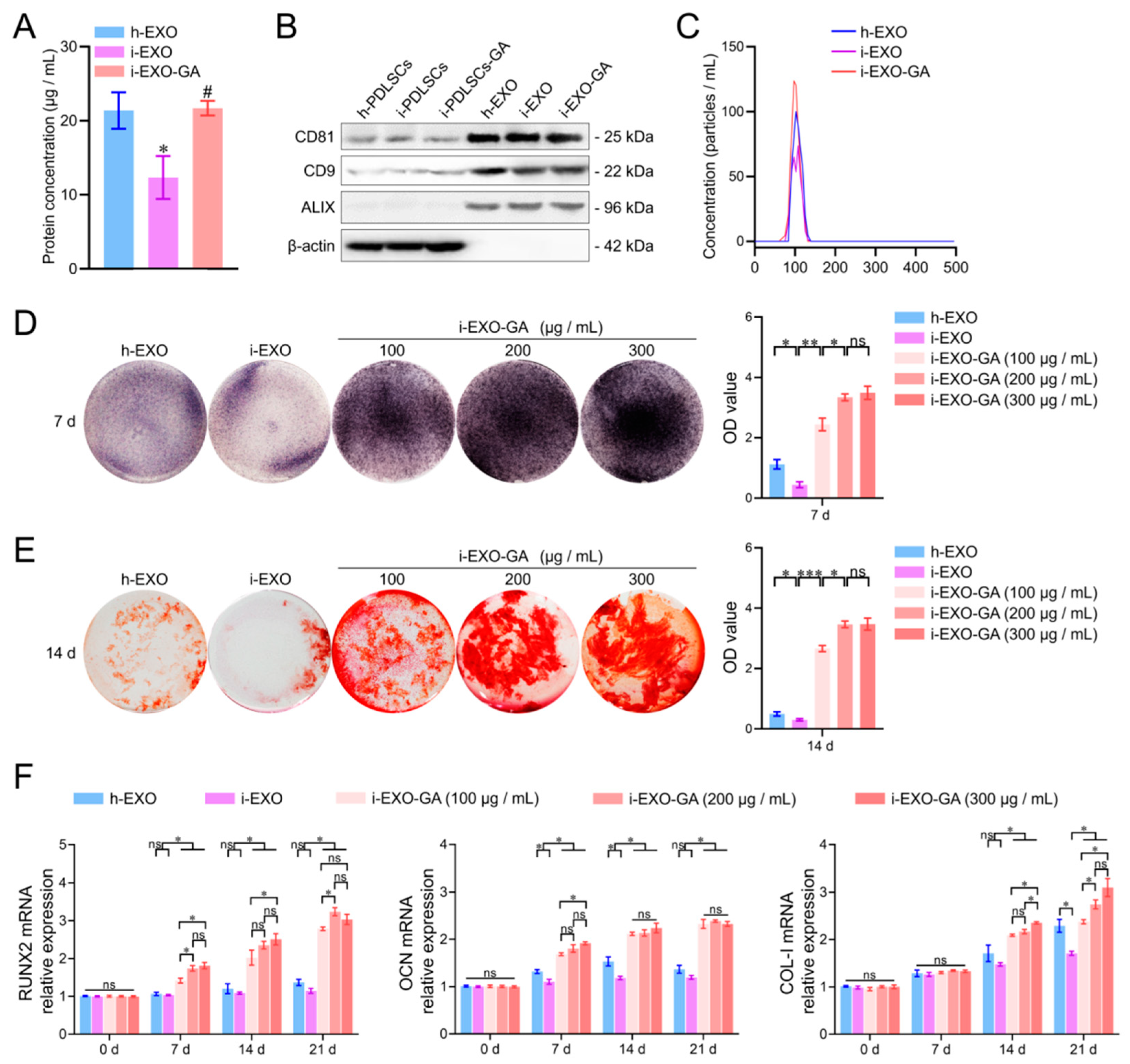
3.6. Identification of PDLSCs Exosomes
3.7. The Function of i-EXO-GA in Promoting Osteodifferentiation and Concentration Screening
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoare, A.; Soto, C.; Rojas-Celis, V.; Bravo, D. Chronic Inflammation as a Link between Periodontitis and Carcinogenesis. Mediat. Inflamm. 2019, 2019, 1029857. [Google Scholar] [CrossRef]
- Van der Velden, U. What exactly distinguishes aggressive from chronic periodontitis: Is it mainly a difference in the degree of bacterial invasiveness? Periodontology 2000 2017, 75, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Abu-Saleh, T. Interleukin-1 composite polymorphism as a risk factor for chronic periodontitis: A review. SADJ 2010, 65, 160–162, 164–166. [Google Scholar] [PubMed]
- Bottino, M.C.; Thomas, V.; Schmidt, G.; Vohra, Y.K.; Chu, T.-M.G.; Kowolik, M.J.; Janowski, G.M. Recent advances in the development of GTR/GBR membranes for periodontal regeneration—A materials perspective. Dent. Mater. 2012, 28, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Tomokiyo, A.; Wada, N.; Maeda, H. Periodontal Ligament Stem Cells: Regenerative Potency in Periodontium. Stem Cells Dev. 2019, 28, 974–985. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Li, X.; Wang, J.; He, X.-T.; Sun, H.-H.; Chen, F.-M. Concise Review: Periodontal Tissue Regeneration Using Stem Cells: Strategies and Translational Considerations. Stem Cells Transl. Med. 2019, 8, 392–403. [Google Scholar] [CrossRef]
- Hua, S.; Bartold, P.; Gulati, K.; Moran, C.; Ivanovski, S.; Han, P. Periodontal and Dental Pulp Cell-Derived Small Extracellular Vesicles: A Review of the Current Status. Nanomaterials 2021, 11, 1858. [Google Scholar] [CrossRef]
- Chew, J.; Chuah, S.J.; Teo, K.; Zhang, S.; Lai, R.C.; Fu, J.H.; Lim, L.P.; Toh, W.S. Mesenchymal stem cell exosomes enhance periodontal ligament cell functions and promote periodontal regeneration. Acta Biomater. 2019, 89, 252–264. [Google Scholar] [CrossRef]
- Liu, L.; Guo, S.; Shi, W.; Liu, Q.; Huo, F.; Wu, Y.; Tian, W. Bone Marrow Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Promote Periodontal Regeneration. Tissue Eng. Part A 2021, 27, 962–976. [Google Scholar] [CrossRef]
- Zhang, Z.; Shuai, Y.; Zhou, F.; Zhou, F.; Yin, J.; Hu, J.; Guo, S.; Wang, Y.; Liu, W. PDLSCs Regulate Angiogenesis of Periodontal Ligaments via VEGF Transferred by Exosomes in Periodontitis. Int. J. Med. Sci. 2020, 17, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, L.; He, Z.; Zhao, H.; Gai, Y.; Song, S.; Yu, Q. Effects of xipayi mouth rinse combined with minocycline on localized aggressive periodontitis’ therapeutic effect and the levels of CRP, TNF-α, IL-6 in serum. Ann. Palliat. Med. 2020, 9, 3410–3417. [Google Scholar] [CrossRef] [PubMed]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing Gallic Acid for Its Broad Spectrum Applications. Mini Rev. Med. Chem. 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Kosuru, R.Y.; Roy, A.; Das, S.K.; Bera, S. Gallic Acid and Gallates in Human Health and Disease: Do Mitochondria Hold the Key to Success. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Ahn, C.-B.; Marasinghe, M.; Je, J.-Y. Insertion of gallic acid onto chitosan promotes the differentiation of osteoblasts from murine bone marrow-derived mesenchymal stem cells. Int. J. Biol. Macromol. 2021, 183, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Liu, Y.; Wang, S. Stem cell-based tooth and periodontal regeneration. Oral. Dis. 2018, 24, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Kang, W.; Li, J.; Yu, L.; Ge, S. An in situ tissue engineering scaffold with growth factors combining angiogenesis and osteoimmunomodulatory functions for advanced periodontal bone regeneration. J. Nanobiotechnol. 2021, 19, 247. [Google Scholar] [CrossRef]
- Mijailovic, I.; Nikolic, N.; Djinic, A.; Carkic, J.; Milinkovic, I.; Peric, M.; Jankovic, S.; Milasin, J.; Aleksic, Z. The down-regulation of Notch 1 signaling contributes to the severity of bone loss in aggressive periodontitis. J. Periodontol. 2020, 91, 554–561. [Google Scholar] [CrossRef]
- Nuñez, J.; Vignoletti, F.; Caffesse, R.G.; Sanz, M. Cellular therapy in periodontal regeneration. Periodontology 2000 2019, 79, 107–116. [Google Scholar] [CrossRef]
- Queiroz, A.; Albuquerque-Souza, E.; Gasparoni, L.M.; de França, B.N.; Pelissari, C.; Trierveiler, M.; Holzhausen, M. Therapeutic potential of periodontal ligament stem cells. World J. Stem Cells 2021, 13, 605–618. [Google Scholar] [CrossRef]
- Lin, B.; Yue, X.; Ding, D.; Dingyu, D.; Yi, X. Advances in macrophage function and its anti-inflammatory and proresolving activity and role in periodontitis development. West China J. Stomatol. 2017, 35, 427–432. [Google Scholar]
- Dai, Z.; Zhen, W.; Li, S. Receptor activator of nuclear factor-κB ligand and tumor necrosis factor-α promotes osteoclast differentiation through the exosomes of inflammatory periodontal ligament stem cells. West China J. Stomatol. 2022, 40, 377–385. [Google Scholar]
- Lu, Y.; Jiang, F.; Jiang, H.; Wu, K.; Zheng, X.; Cai, Y.; Katakowski, M.; Chopp, M.; To, S.-S.T. Gallic acid suppresses cell viability, proliferation, invasion and angiogenesis in human glioma cells. Eur. J. Pharmacol. 2010, 641, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, M.; Velmurugan, B.; Rajamanickam, S.; Agarwal, R.; Agarwal, C. Gallic Acid, an Active Constituent of Grape Seed Extract, Exhibits Anti-proliferative, Pro-apoptotic and Anti-tumorigenic Effects Against Prostate Carcinoma Xenograft Growth in Nude Mice. Pharm. Res. 2009, 26, 2133–2140. [Google Scholar] [CrossRef] [PubMed]
- Choińska, R.; Dąbrowska, K.; Świsłocka, R.; Lewandowski, W.; Świergiel, A.H. Antimicrobial Properties of Mandelic Acid, Gallic Acid and their Derivatives. Mini-Rev. Med. Chem. 2021, 21, 2544–2550. [Google Scholar] [CrossRef]
- Zahrani, N.A.A.; El-Shishtawy, R.M.; Asiri, A.M. Recent developments of gallic acid derivatives and their hybrids in medicinal chemistry: A review. Eur. J. Med. Chem. 2020, 204, 112609. [Google Scholar] [CrossRef]
- Hamulati, W.; Aibibai, Y.; Zhong, L.J. Inhibitory effect of xipayi mouth rinse on the secretion of IL-6 from human gingival fibroblast induced by lipopolysaccharide. J. Cent. South Univ. Technol. 2006, 31, 326–328. [Google Scholar]
- Dehghani, M.A.; Shakiba, M.N.; Moghimipour, E.; Khorsandi, L.; Khah, M.A.; Mahdavinia, M. Protective effect of gallic acid and gallic acid-loaded Eudragit-RS 100 nanoparticles on cisplatin-induced mitochondrial dysfunction and inflammation in rat kidney. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165911. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cai, W.; Zhao, S.; Shi, L.; Chen, Y.; Li, X.; Sun, X.; Mao, Y.; He, B.; Hou, Y.; et al. Oxidative stress-related biomarkers in saliva and gingival crevicular fluid associated with chronic periodontitis: A systematic review and meta-analysis. J. Clin. Periodontol. 2019, 46, 608–622. [Google Scholar] [CrossRef]
- Sczepanik, F.S.C.; Grossi, M.L.; Casati, M.; Goldberg, M.; Glogauer, M.; Fine, N.; Tenenbaum, H.C. Periodontitis is an inflammatory disease of oxidative stress: We should treat it that way. Periodontol. 2000 2020, 84, 45–68. [Google Scholar] [CrossRef]
- Gao, J.; Feng, Z.; Wang, X.; Zeng, M.; Liu, J.; Han, S.; Xu, J.; Chen, L.; Cao, L.; Long, J.; et al. SIRT3/SOD2 maintains osteoblast differentiation and bone formation by regulating mitochondrial stress. Cell Death Differ. 2018, 25, 229–240. [Google Scholar] [CrossRef]
- Tanaka, M.; Kishimoto, Y.; Sasaki, M.; Sato, A.; Kamiya, T.; Kondo, K.; Iida, K. Terminalia bellirica (Gaertn.) Roxb.Extract and Gallic Acid Attenuate LPS-Induced Inflammation and Oxidative Stress via MAPK/NF-κB and Akt/AMPK/Nrf2 Pathways. Oxid. Med. Cell. Longev. 2018, 2018, 9364364. [Google Scholar] [CrossRef]
- Sokolov, S.S.; Balakireva, A.V.; Markova, O.V.; Severin, F.F. Negative Feedback of Glycolysis and Oxidative Phosphorylation: Mechanisms of and Reasons for It. Biochemistry 2015, 80, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. The investigation of energy metabolism in osteoblasts and osteoclasts. West China J. Stomatol. 2021, 39, 501–509. [Google Scholar]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome Theranostics: Biology and Translational Medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef]
- Gupta, D.; Zickler, A.M.; El Andaloussi, S. Dosing extracellular vesicles. Adv. Drug Deliv. Rev. 2021, 178, 113961. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, H.S. Extracellular Vesicles in Regenerative Medicine: Potentials and Challenges. Tissue Eng. Regen. Med. 2021, 18, 479–484. [Google Scholar] [CrossRef]
- Hwang, H.S.; Kim, H.; Han, G.; Lee, J.W.; Kim, K.; Kwon, I.C.; Yang, Y.; Kim, S.H. Extracellular Vesicles as Potential Therapeutics for Inflammatory Diseases. Int. J. Mol. Sci. 2021, 22, 5478. [Google Scholar] [CrossRef]
- Languino, L.R.; Hooper, D.C. Activated Extracellular Vesicles as New Therapeutic Targets? Trends Cell Biol. 2019, 29, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, H.; Ghorbani, F.; Derakhshani, M.; Movassaghpour, A.; Yousefi, M. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles: A novel therapeutic paradigm. J. Cell. Physiol. 2020, 235, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Lei, F.; Li, M.; Lin, T.; Zhou, H.; Wang, F.; Su, X. Treatment of inflammatory bone loss in periodontitis by stem cell-derived exosomes. Acta Biomater. 2022, 141, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, Z.; Liu, P.; Hu, Y.; Li, T.; Yang, J.; Gao, P.; Xu, Q. Exosomes Derived From Human Gingival Mesenchymal Stem Cells Attenuate the Inflammatory Response in Periodontal Ligament Stem Cells. Front. Chem. 2022, 10, 863364. [Google Scholar] [CrossRef]
- Zheng, Y.; Dong, C.; Yang, J.; Jin, Y.; Zheng, W.; Zhou, Q.; Liang, Y.; Bao, L.; Feng, G.; Ji, J.; et al. Exosomal microRNA-155-5p from PDLSCs regulated Th17/Treg balance by targeting sirtuin-1 in chronic periodontitis. J. Cell. Physiol. 2019, 234, 20662–20674. [Google Scholar] [CrossRef]
- Novello, S.; Tricot-Doleux, S.; Novella, A.; Pellen-Mussi, P.; Jeanne, S. Influence of Periodontal Ligament Stem Cell-Derived Conditioned Medium on Osteoblasts. Pharmaceutics 2022, 14, 729. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Ye, Y.; He, S.; Song, J. SHED-derived conditioned exosomes enhance the osteogenic differentiation of PDLSCs via Wnt and BMP signaling in vitro. Differentiation 2020, 111, 1–11. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer Sequences |
|---|---|---|
| NDUFB-3 | Forward | TCAGATTGCTGTCAGACATGG |
| Reverse | TGGTGTCCCTTCTATCTTCCA | |
| SDHB | Forward | AAATGTGGCCCCATGGTATTG |
| Reverse | AGAGCCACAGATGCCTTCTCT | |
| β-tubulin | Forward | CCCAACAATGTGAAGACGG |
| Reverse | GCCTCGGTGAACTCCATCT | |
| CS | Forward | CGGCTACCACATCCAAGGAA |
| Reverse | GCTGGAATTACCGCGGCT | |
| IDH | Forward | TCACCAAATGGCACCATACGA |
| Reverse | GCCAACATGACTTACTTGATCCC |
| Gene | Direction | Primer Sequences |
|---|---|---|
| OCN | Forward | GACCCTCTCTCTGCTCACT |
| Reverse | CACCTTACTGCCCTCCTGC | |
| RUNX2 | Forward | GTAGAGAGCAGGGAAGAC |
| Reverse | GCTTGGATTAGGGAGTCAC | |
| CoL-I | Forward | TTCTCCTGGCAAAGACGGAC |
| Reverse | TTGCTGTTGAAGTCGCAGGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Z.; Li, Z.; Zheng, W.; Yan, Z.; Zhang, L.; Yang, J.; Xiao, J.; Sun, H.; Li, S.; Huang, W. Gallic Acid Ameliorates the Inflammatory State of Periodontal Ligament Stem Cells and Promotes Pro-Osteodifferentiation Capabilities of Inflammatory Stem Cell-Derived Exosomes. Life 2022, 12, 1392. https://doi.org/10.3390/life12091392
Dai Z, Li Z, Zheng W, Yan Z, Zhang L, Yang J, Xiao J, Sun H, Li S, Huang W. Gallic Acid Ameliorates the Inflammatory State of Periodontal Ligament Stem Cells and Promotes Pro-Osteodifferentiation Capabilities of Inflammatory Stem Cell-Derived Exosomes. Life. 2022; 12(9):1392. https://doi.org/10.3390/life12091392
Chicago/Turabian StyleDai, Zhenning, Ziyue Li, Weihan Zheng, Zi Yan, Lijun Zhang, Jiaxin Yang, Jing Xiao, Hanxiao Sun, Shiyu Li, and Wenhua Huang. 2022. "Gallic Acid Ameliorates the Inflammatory State of Periodontal Ligament Stem Cells and Promotes Pro-Osteodifferentiation Capabilities of Inflammatory Stem Cell-Derived Exosomes" Life 12, no. 9: 1392. https://doi.org/10.3390/life12091392
APA StyleDai, Z., Li, Z., Zheng, W., Yan, Z., Zhang, L., Yang, J., Xiao, J., Sun, H., Li, S., & Huang, W. (2022). Gallic Acid Ameliorates the Inflammatory State of Periodontal Ligament Stem Cells and Promotes Pro-Osteodifferentiation Capabilities of Inflammatory Stem Cell-Derived Exosomes. Life, 12(9), 1392. https://doi.org/10.3390/life12091392