
The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue



 and
and 
Abstract
1. Introduction
2. Results
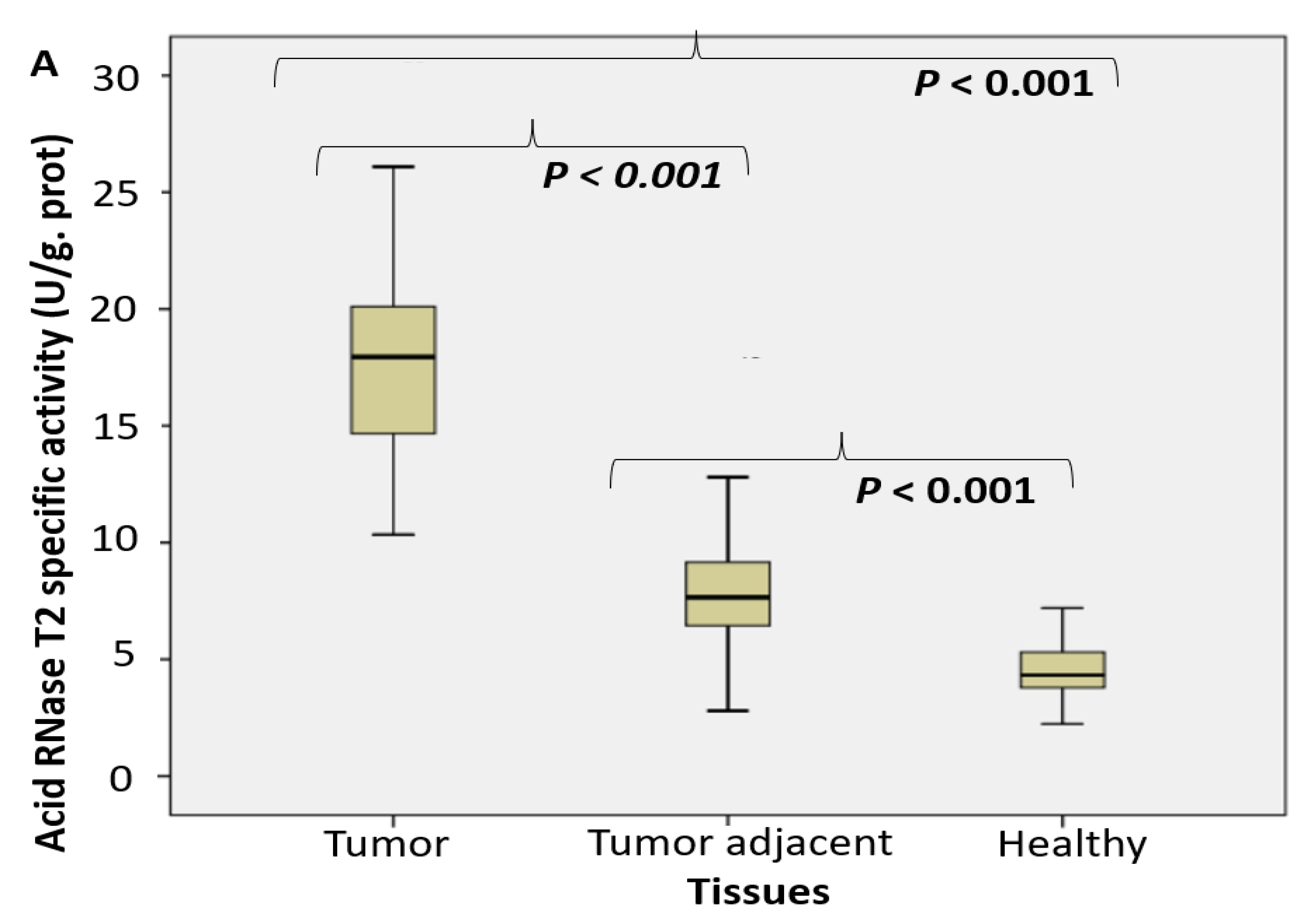
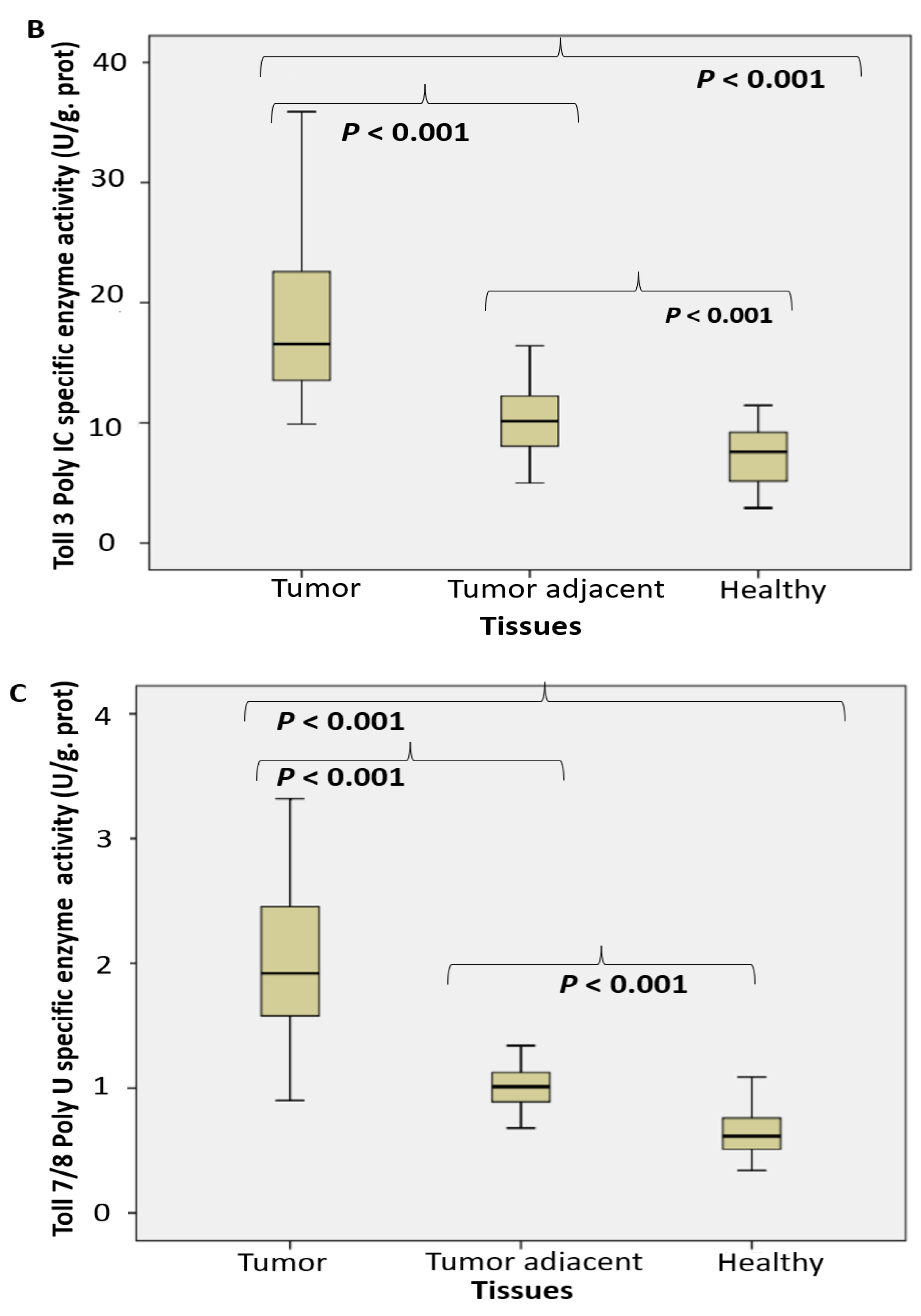
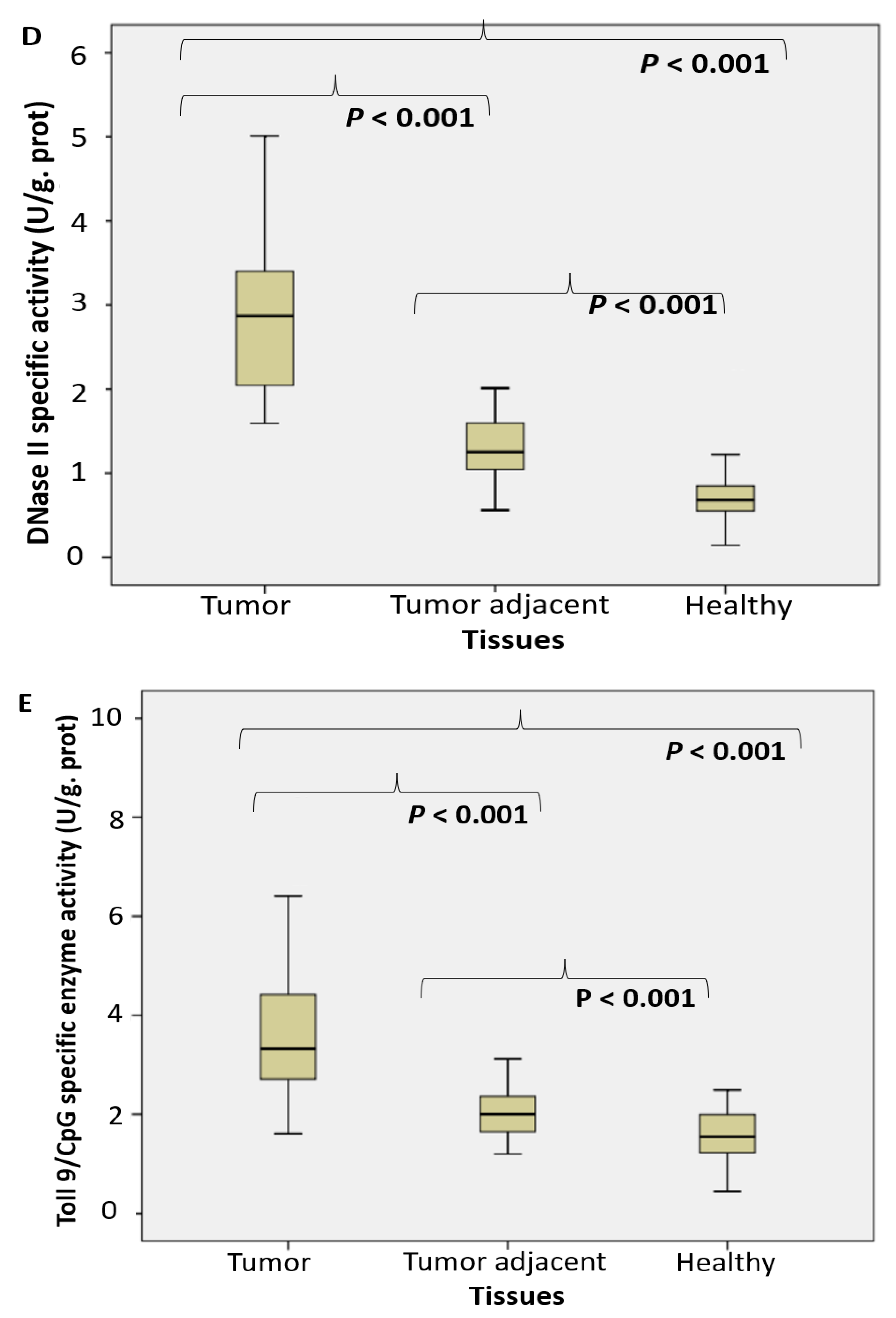
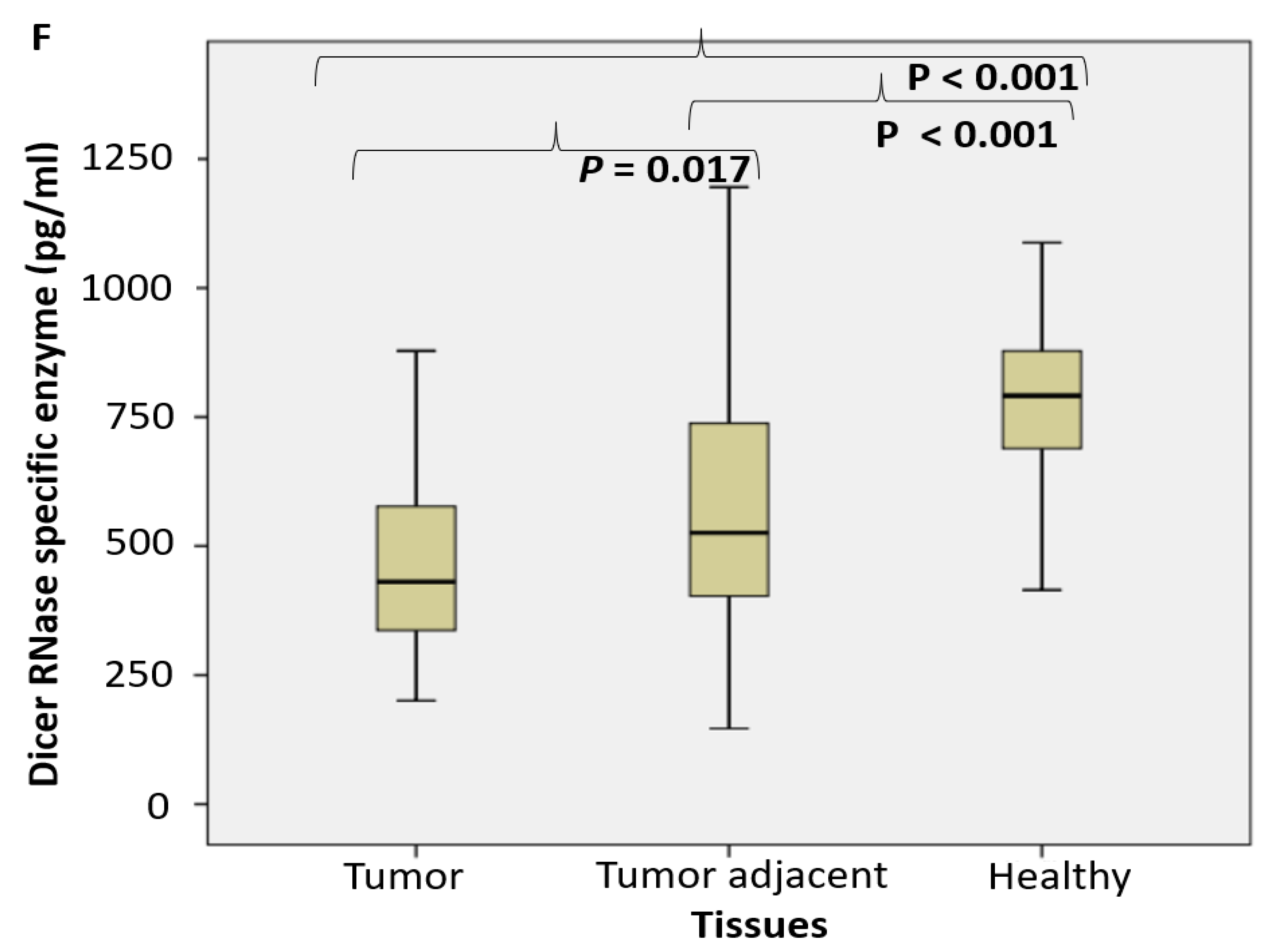
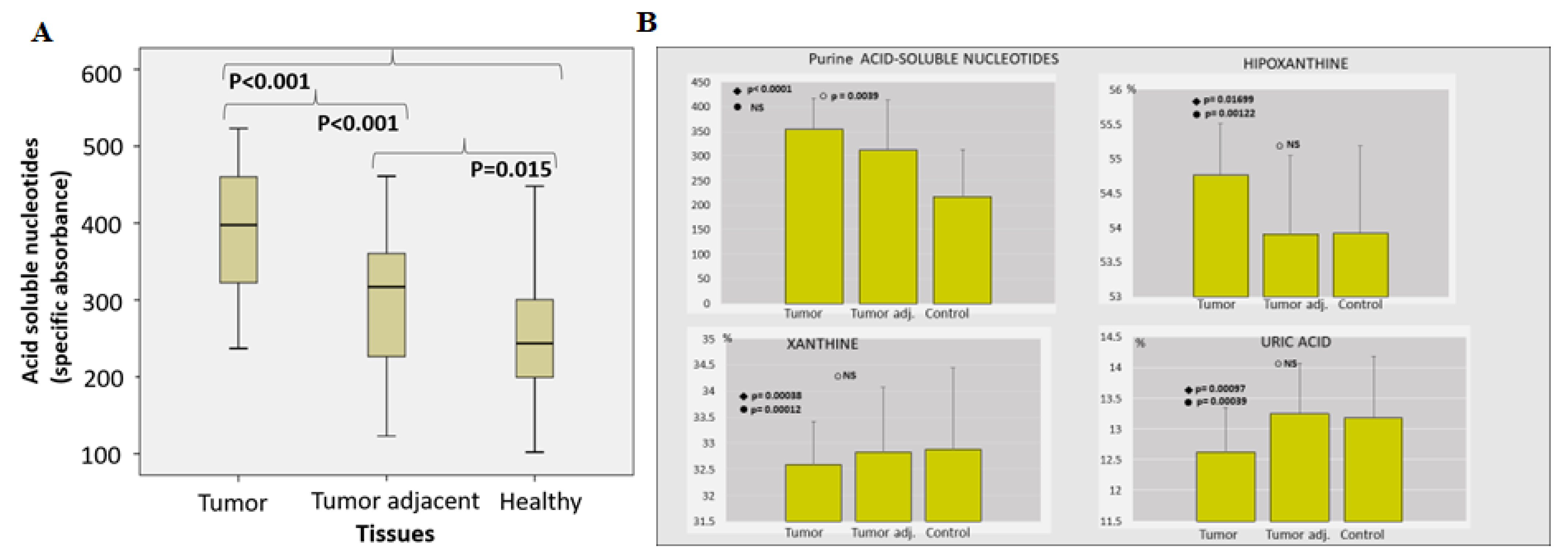
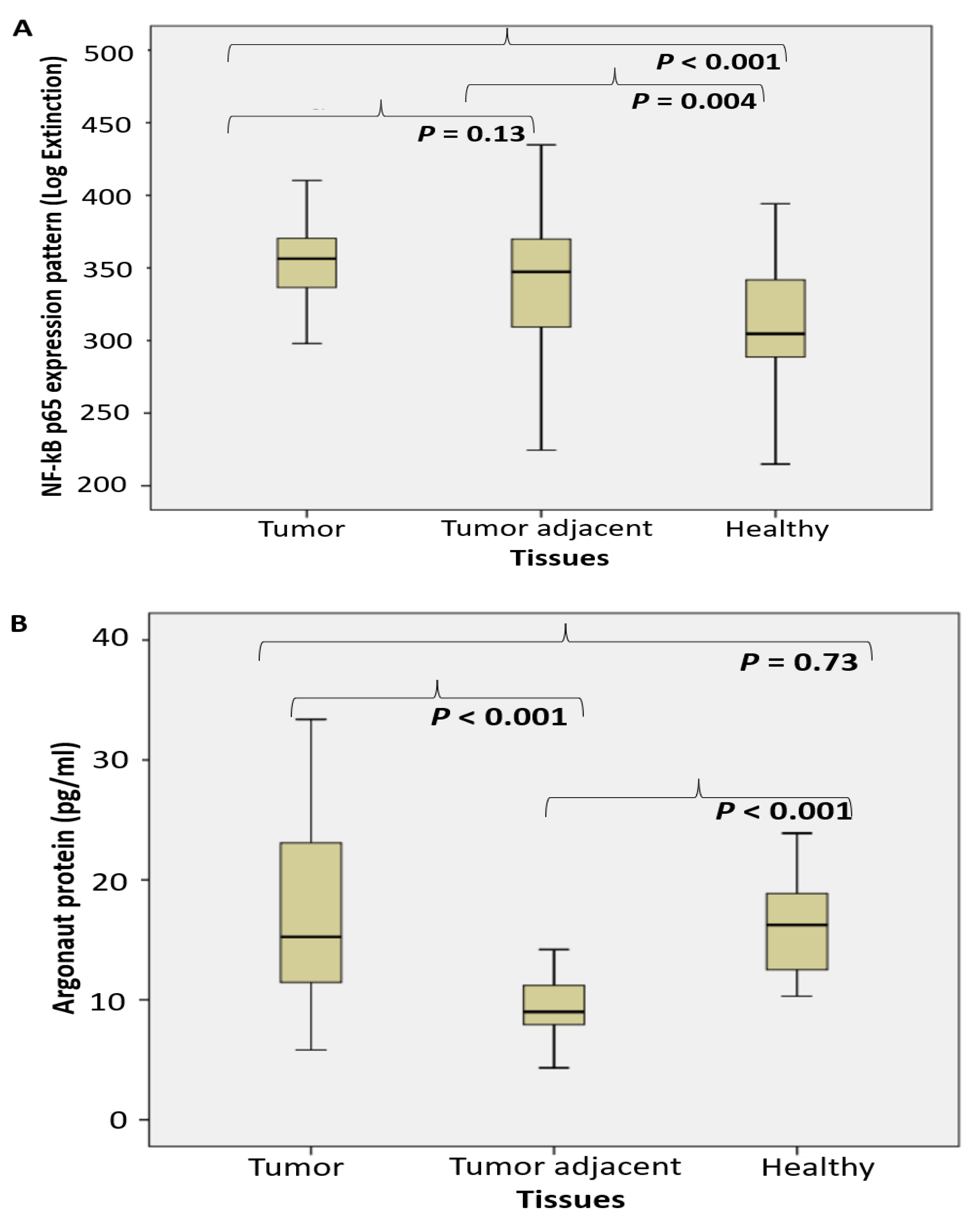
2.1. Enzyme Assays
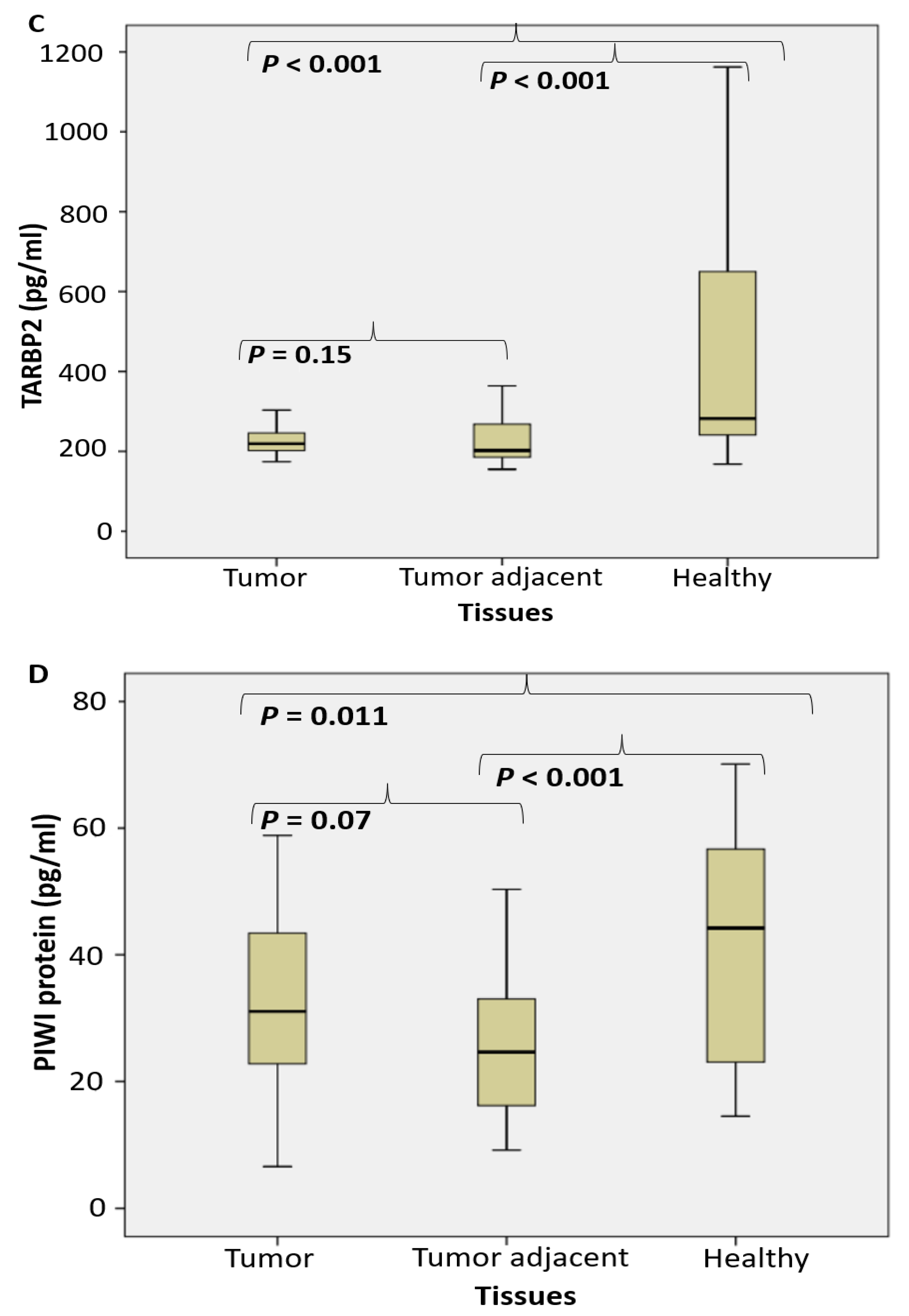
2.2. Proteomic Analysis
3. ROC (Receiver Operating Characteristic) Curves
4. Discussion
5. Materials and Methods
5.1. Patient Selection
5.2. Tissue Preparation
5.3. Enzyme Assays
5.4. Determination of NF-κB p65 in Prostate Tissue Samples
5.5. Statistical Analyses
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Zhao, S.; Zhang, Y.; Zhang, Q.; Wang, F.; Zhang, D. Toll-like receptors and prostate cancer. Front. Immunol. 2014, 5, 352. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Sayyid, R.K.; Benton, J.Z.; Reed, W.C.; Woodruff, P.; Terris, M.K.; Wallis, C.J.D.; Klaassen, Z. Prostate cancer mortality rates in low- and favorable intermediate-risk active surveillance patients: A population-based competing risks analysis. World J. Urol. 2022, 1–7. [Google Scholar] [CrossRef]
- Beatty, G.L.; Gladney, W.L. Immune escape mechanisms as a guide for cancer immunotherapy. Clin. Cancer Res. 2015, 21, 687–692. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, Q.; Yu, L.; Zhu, D.; Li, Y.; Xue, Z.; Hua, Z.; Luo, X.; Song, Z.; Lu, C.; et al. The role of miRNA in tumor immune escape and miRNA-based therapeutic strategies. Front. Immunol. 2022, 12, 807895. [Google Scholar] [CrossRef] [PubMed]
- Lind, N.A.; Rael, V.E.; Pestal, K.; Liu, B.; Barton, G.M. Regulation of the nucleic acid-sensing Toll-like receptors. Nat. Rev. Immunol. 2021, 22, 224–235. [Google Scholar] [CrossRef]
- Blasius, A.L.; Beutler, B. Intracellular toll-like receptors. Immunity 2010, 32, 305–315. [Google Scholar] [CrossRef]
- Krieg, A.M. Development of TLR9 agonists for cancer therapy. J. Clin. Investig. 2007, 117, 1184–1194. [Google Scholar] [CrossRef]
- Väisänen, M.R.; Jukkola-Vuorinen, A.; Vuopala, K.S.; Selander, K.S.; Vaarala, M.H. Expression of Toll-like receptor-9 is associated with poor progression-free survival in prostate cancer. Oncol. Lett. 2013, 5, 1659–1663. [Google Scholar] [CrossRef]
- Cherfils-Vicini, J.; Platonova, S.; Gillard, M.; Laurans, L.; Validire, P.; Caliandro, R.; Magdeleinat, P.; Mami-Chouaib, F.; Dieu-Nosjean, M.-C.; Fridman, W.H.; et al. Triggering of TLR7 and TLR8 expressed by human lung cancer cells induces cell survival and chemoresistance. J. Clin. Investig. 2010, 120, 1285–1297. [Google Scholar] [CrossRef]
- Miyake, K.; Saitoh, S.I.; Fukui, R.; Shibata, T.; Sato, R.; Murakami, Y. Dynamic control of nucleic-acid-sensing Toll-like receptors by the endosomal compartment. Int. Immunol. 2021, 33, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Greulich, W.; Wagner, M.; Gaidt, M.M.; Stafford, C.; Cheng, Y.; Linder, A.; Carell, T.; Hornung, V. TLR8 Is a Sensor of RNase T2 Degradation Products. Cell 2019, 179, 1264–1275.e13. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K.; Shibata, T.; Fukui, R.; Sato, R.; Saitoh, S.I.; Murakami, Y. Nucleic Acid Sensing by Toll-Like Receptors in the Endosomal Compartment. Front. Immunol. 2022, 13, 941931. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.; Zhang, Q.; Hossain, D.M.; Nechaev, S.; Li, H.; Kowolik, C.M.; D’Apuzzo, M.; Forman, S.; Jones, J.; Pal, S.K.; et al. TLR9 signaling through NF-κB/RELA and STAT3 promotes tumor-propagating potential of prostate cancer cells. Oncotarget 2015, 6, 17302–17313. [Google Scholar] [CrossRef]
- Tinari, A.; Matarrese, P.; Minetti, M.; Malorni, W. Hyperphagia by self- and xeno-cannibalism: Cell death by indigestion? A reminiscence of the Phedrus Fabula “Rana Rupta et Bos”? Autophagy 2008, 4, 128–130. [Google Scholar] [CrossRef][Green Version]
- Degenhardt, K.; Mathew, R.; Beaudoin, B.; Bray, K.; Anderson, D.; Chen, G.; Mukherjee, C.; Shi, Y.; Gélinas, C.; Fan, Y.; et al. Autophagy promotes tumor cell survival and restricts necrosis, inflammation, and tumorigenesis. Cancer Cell 2006, 10, 51–64. [Google Scholar] [CrossRef]
- Lugini, L.; Matarrese, P.; Tinari, A.; Lozupone, F.; Federici, C.; Iessi, E.; Gentile, M.; Luciani, F.; Parmiani, G.; Rivoltini, L.; et al. Cannibalism of live lymphocytes by human metastatic but not primary melanoma cells. Cancer Res. 2006, 66, 3629–3638. [Google Scholar] [CrossRef]
- Matarrese, P.; Ciarlo, L.; Tinari, A.; Piacentini, M.; Malorni, W. Xeno-cannibalism as an exacerbation of self-cannibalism: A possible fruitful survival strategy for cancer cells. Curr. Pharm. Des. 2008, 14, 245–252. [Google Scholar]
- Fujiwara, Y.; Wada, K.; Kabuta, T. Lysosomal degradation of intracellular nucleic acids—Multiple autophagic pathways. J. Biochem. 2017, 161, 145–154. [Google Scholar] [CrossRef]
- Wu, L.; Xu, Y.; Zhao, H.; Li, Y. RNase T2 in inflammation and cancer: Immunological and biological views. Front. Immunol. 2020, 11, 1554. [Google Scholar] [CrossRef]
- Lauková, L.; Konečná, B.; Janovičová, Ľ.; Vlková, B.; Celec, P. Deoxyribonucleases and their applications in biomedicine. Biomolecules 2020, 10, 1036. [Google Scholar] [CrossRef] [PubMed]
- Thomson, T.; Lin, H. The biogenesis and function PIWI proteins and piRNAs: Progress and prospect. Annu. Rev. Cell Dev. Biol. 2009, 25, 355–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Chen, H.; Zhang, L.; Dakhova, O.; Zhang, Y.; Lewis, M.T.; Creighton, C.J.; Ittmann, M.M.; Xin, L. A dosage-dependent pleiotropic role of Dicer in prostate cancer growth and metastasis. Oncogene 2014, 33, 3099–3108. [Google Scholar] [CrossRef] [PubMed]
- Nowak, I.; Sarshad, A.A. Argonaute proteins take center stage in cancers. Cancers 2021, 13, 788. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, Z. The role of TARBP2 in the development and progression of cancers. Tumor Biol. 2016, 37, 57–60. [Google Scholar] [CrossRef]
- Liu, Y. MicroRNAs and PIWI-interacting RNAs in oncology. Oncol. Lett. 2016, 12, 2289–2292. [Google Scholar] [CrossRef]
- FDA-NIH Biomarker Working Group. BEST (Biomarkers, EndpointS, and other Tools) Resource; Food and Drug Administration (US): Silver Spring, MD, USA; National Institutes of Health (US): Bethesda, MD, USA, 2016.
- Kocić, G.; Hadzi-Djokić, J.; Veljković, A.; Roumeliotis, S.; Janković-Veličković, L.; Šmelcerović, A. Template-independent poly(A)-tail decay and RNASEL as potential cellular biomarkers for prostate cancer development. Cancers 2022, 14, 2239. [Google Scholar] [CrossRef]
- Adekoya, T.O.; Richardson, R.M. Cytokines and chemokines as mediators of prostate cancer metastasis. Int. J. Mol. Sci. 2020, 21, 4449. [Google Scholar] [CrossRef]
- Shankaran, H.; Wiley, H.S.; Resat, H. Receptor downregulation and desensitization enhance the information processing ability of signalling receptors. BMC Syst. Biol. 2007, 1, 48. [Google Scholar] [CrossRef]
- Maelfait, J.; Liverpool, L.; Rehwinkel, J. Nucleic acid sensors and programmed cell death. J. Mol. Biol. 2020, 432, 552–568. [Google Scholar] [CrossRef]
- Sharma, S.; Fitzgerald, K.A.; Cancro, M.P.; Marshak-Rothstein, A. Nucleic acid–sensing receptors: Rheostats of autoimmunity and autoinflammation. J. Immunol. 2015, 195, 3507–3512. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ren, W.; Huang, X.; Deng, J.; Li, T.; Yin, Y. Potential mechanisms connecting purine metabolism and cancer therapy. Front. Immunol. 2018, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Calì, B.; Molon, B.; Viola, A. Tuning cancer fate: The unremitting role of host immunity. Open Biol. 2017, 7, 170006. [Google Scholar] [CrossRef]
- Medler, T.; Patel, J.M.; Alice, A.; Baird, J.R.; Hu, H.M.; Gough, M.J. Activating the nucleic acid-sensing machinery for anticancer immunity. Int. Rev. Cell Mol. Biol. 2019, 344, 173–214. [Google Scholar]
- Karapetyan, L.; Luke, J.J.; Davar, D. Toll-like receptor 9 agonists in cancer. OncoTargets Ther. 2020, 13, 10039. [Google Scholar] [CrossRef] [PubMed]
- Lubaroff, D.M.; Karan, D. CpG oligonucleotide as an adjuvant for the treatment of prostate cancer. Adv. Drug Deliv. Rev. 2009, 61, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Gosu, V.; Basith, S.; Kwon, O.P.; Choi, S. Therapeutic applications of nucleic acids and their analogues in Toll-like receptor signaling. Molecules 2012, 17, 13503–13529. [Google Scholar] [CrossRef]
- McWhirter, S.M.; Jefferies, C.A. Nucleic acid sensors as therapeutic targets for human disease. Immunity 2020, 52, 78–97. [Google Scholar] [CrossRef]
- Chen, M.; Hu, S.; Li, Y.; TJiang, T.; Jin, H.; Feng, L. Targeting nuclear acid-mediated immunity in cancer immune checkpoint inhibitor therapies. Signal Transduct. Target. Ther. 2020, 5, 270. [Google Scholar]
- Kolarevic, A.; Yancheva, D.; Kocic, G.; Smelcerovic, A. Deoxyribonuclease inhibitors. Eur. J. Med. Chem. 2014, 88, 101–111. [Google Scholar] [CrossRef]
- Piasentin, N.; Milotti, E.; Chignola, R. The control of acidity in tumor cells: A biophysical model. Sci. Rep. 2020, 10, 13613. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, R.E.; Narui, R.; Wada, H. Effects of alkalization therapy on chemotherapy outcomes in metastatic or recurrent pancreatic cancer. Anticancer Res. 2020, 40, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Staal, J.; Beyaert, R. Inflammation and NF-κB signaling in prostate cancer: Mechanisms and clinical implications. Cells 2018, 7, 122. [Google Scholar] [CrossRef] [PubMed]
- Zelivianski, S.; Glowacki, R.; Lin, M.F. Transcriptional activation of the human prostatic acid phosphatase gene by NF-kappaB via a novel hexanucleotide-binding site. Nucleic Acids Res. 2004, 32, 3566–3580. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Zhu, S.; Deng, S.; Zou, S.S.; Gao, B.; Zang, G.; Wu, J.; Jiang, Y.; Liu, Y.-J.; Chen, J. Human cancer cells sense cytosolic nucleic acids through the RIG-I–MAVS pathway and cGAS–STING pathway. Front. Cell Dev. Biol. 2021, 8, 606001. [Google Scholar] [CrossRef]
- Bayraktar, R.; Bertilaccio, M.T.; Calin, G.A. The interaction between two worlds: MicroRNAs and toll-like receptors. Front. Immunol. 2019, 10, 1053. [Google Scholar] [CrossRef] [PubMed]
- Forsbach, A.; Nemorin, J.G.; Montino, C.; Müller, C.; Samulowitz, U.; Vicari, A.P.; Jurk, M.; Mutwiri, G.K.; Krieg, A.M.; Lipford, G.B.; et al. Identification of RNA sequence motifs stimulating sequence-specific TLR8-dependent immune responses. J. Immunol. 2008, 180, 3729–3738. [Google Scholar] [CrossRef]
- Völler, D.; Linck, L.; Bruckmann, A.; Hauptmann, J.; Deutzmann, R.; Meister, G.; Bosserhoff, A.K. Argonaute family protein expression in normal tissue and cancer entities. PLoS ONE 2016, 11, e0161165. [Google Scholar] [CrossRef]
- Li, L.; Yu, C.; Gao, H.; Li, Y. Argonaute proteins: Potential biomarkers for human colon cancer. BMC Cancer 2010, 10, 38. [Google Scholar] [CrossRef]
- Alshalalfa, M.; Bader, G.D.; Bismar, T.A.; Alhajj, R. Coordinate microRNA-mediated regulation of protein complexes in prostate cancer. PLoS ONE 2013, 8, e84261. [Google Scholar] [CrossRef]
- Humphrey, P.A.; Moch, H.; Cubilla, A.L.; Ulbright, T.M.; Reuter, V.E. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs-Part B: Prostate and Bladder Tumours. Eur. Urol. 2016, 70, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Oesterling, J.E.; Jacobsen, S.J.; Chute, C.G.; Guess, H.A.; Girman, C.J.; Panser, L.A.; Lieber, M.M. Serum prostate-specific antigen in a community-based population of healthy men. Establishment of age-specific reference ranges. JAMA 1993, 270, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Kocic, G.; Bjelakovic, G.; Pavlovic, D.; Jevtovic, T.; Pavlovic, V.; Sokolovic, D.; Basic, J.; Cekic, S.; Cvetkovic, T.; Kocic, R.; et al. Protective effect of interferon-alpha on the DNA- and RNA-degrading pathway in anti-Fas-antibody induced apoptosis. Hepatol. Res. 2007, 37, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Kocic, G.; Bjelakovic, G.; Saranac Lj Kocic, R.; Jevtovic, T.; Sokolovic, D.; Nikolic, G.; Pavlovic, D.; Stojanovic, S. Altered degradation of circulating nucleic acids and oligonucleotides in diabetic patients. Diabetes Res. Clin. Pract. 2008, 79, 204–213. [Google Scholar] [CrossRef]
- Akagi, K.; Yamanaka, M.; Murai, K.; Omae, T. Purification and properties of acid ribonucleases in human serum and leukocytes. Cancer Res. 1978, 38, 2163–2167. [Google Scholar] [PubMed]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the pholin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Dudzinska, W.; Lubkowska, A. Changes in the concentration of purine and pyridine as a response to single whole-body cryostimulation. Front. Physiol. 2021, 12, 634816. [Google Scholar] [CrossRef]
- Kocic, G.; Veljkovic, A.; Kocic, H.; Colic, M.; Mihajlovic, D.; Tomovic, K.; Stojanovic, S.; Smelcerovic, A. Depurinized milk downregulates rat thymus MyD88/Akt/p38 function, NF-κB-mediated inflammation, caspase-1 activity but not the endonuclease pathway: In vitro/in vivo study. Sci. Rep. 2017, 7, 41971. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Investigated Parameters | X ± SD |
|---|---|
| Age | 67.05 ± 4.35 (range 26–73 years) |
| tPSA (ng/mL) | 17.26 ± 9.88 (range 5.42–36.45) |
| Gleason score | 6.69 ± 0.55 (range 6–8) |
| Tumor stage (TNM) | 2.33 ± 0.48 (range 2–3) |
| Marker | AUC | 95% CI | p | Cut-Off Value | Sensitivity (%) | Specificity (%) |
|---|---|---|---|---|---|---|
| RNase T2 specific enzyme activity | 0.99 | 0.98–1.00 | <0.0001 | 10.3 | 100 | 92.5 |
| DNase II specific enzyme activity | 0.98 | 0.96–1.00 | <0.0001 | 1.65 | 97.5 | 88.8 |
| TLR9 (CpG) specific enzyme activity | 0.92 | 0.86–0.97 | <0.0001 | 2.49 | 80 | 91.3 |
| TLR3 (Poly I:C) specific enzyme activity | 0.95 | 0.92–0.99 | <0.0001 | 12.8 | 85 | 91.3 |
| TLR7/8 (Poly U) specific enzyme activity | 0.97 | 0.95–1.00 | <0.0001 | 0.98 | 97.5 | 68.8 |
| Argonaut AGO2 protein | 0.66 | 0.56–0.78 | 0.004 | 22.8 | 27.5 | 96.3 |
| NF-κB p65 expression pattern | 0.69 | 0.60–0.79 | 0.001 | 326 | 87.5 | 48.8 |
| Dicer RNase specific enzyme | 0.76 | 0.67–0.84 | <0.0001 | 695 | 90 | 51.3 |
| Acid soluble nucleotides (specific absorbance) | 0.81 | 0.73–0.89 | <0.0001 | 314 | 82.5 | 66.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocic, G.; Hadzi-Djokic, J.; Colic, M.; Veljkovic, A.; Tomovic, K.; Roumeliotis, S.; Smelcerovic, A.; Liakopoulos, V. The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue. Int. J. Mol. Sci. 2023, 24, 509. https://doi.org/10.3390/ijms24010509
Kocic G, Hadzi-Djokic J, Colic M, Veljkovic A, Tomovic K, Roumeliotis S, Smelcerovic A, Liakopoulos V. The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue. International Journal of Molecular Sciences. 2023; 24(1):509. https://doi.org/10.3390/ijms24010509
Chicago/Turabian StyleKocic, Gordana, Jovan Hadzi-Djokic, Miodrag Colic, Andrej Veljkovic, Katarina Tomovic, Stefanos Roumeliotis, Andrija Smelcerovic, and Vassilios Liakopoulos. 2023. "The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue" International Journal of Molecular Sciences 24, no. 1: 509. https://doi.org/10.3390/ijms24010509
APA StyleKocic, G., Hadzi-Djokic, J., Colic, M., Veljkovic, A., Tomovic, K., Roumeliotis, S., Smelcerovic, A., & Liakopoulos, V. (2023). The Role of Nucleases Cleaving TLR3, TLR7/8 and TLR9 Ligands, Dicer RNase and miRNA/piRNA Proteins in Functional Adaptation to the Immune Escape and Xenophagy of Prostate Cancer Tissue. International Journal of Molecular Sciences, 24(1), 509. https://doi.org/10.3390/ijms24010509