
Diverse Galactooligosaccharides Differentially Reduce LPS-Induced Inflammation in Macrophages


Abstract
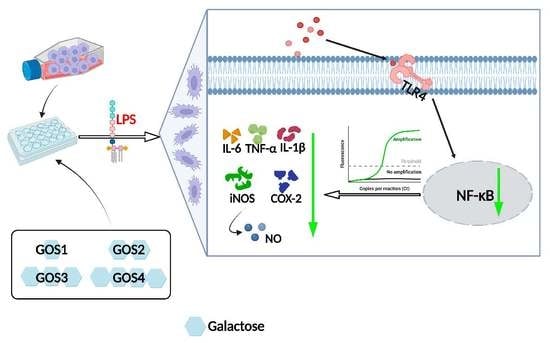
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Viability
2.4. Determination of NO, IL-1β, TNF-α, and IL-6 Levels
2.5. Isolation of RNA and RT-PCR
2.6. Western Blotting Analysis
2.7. Statistical Analysis
3. Results
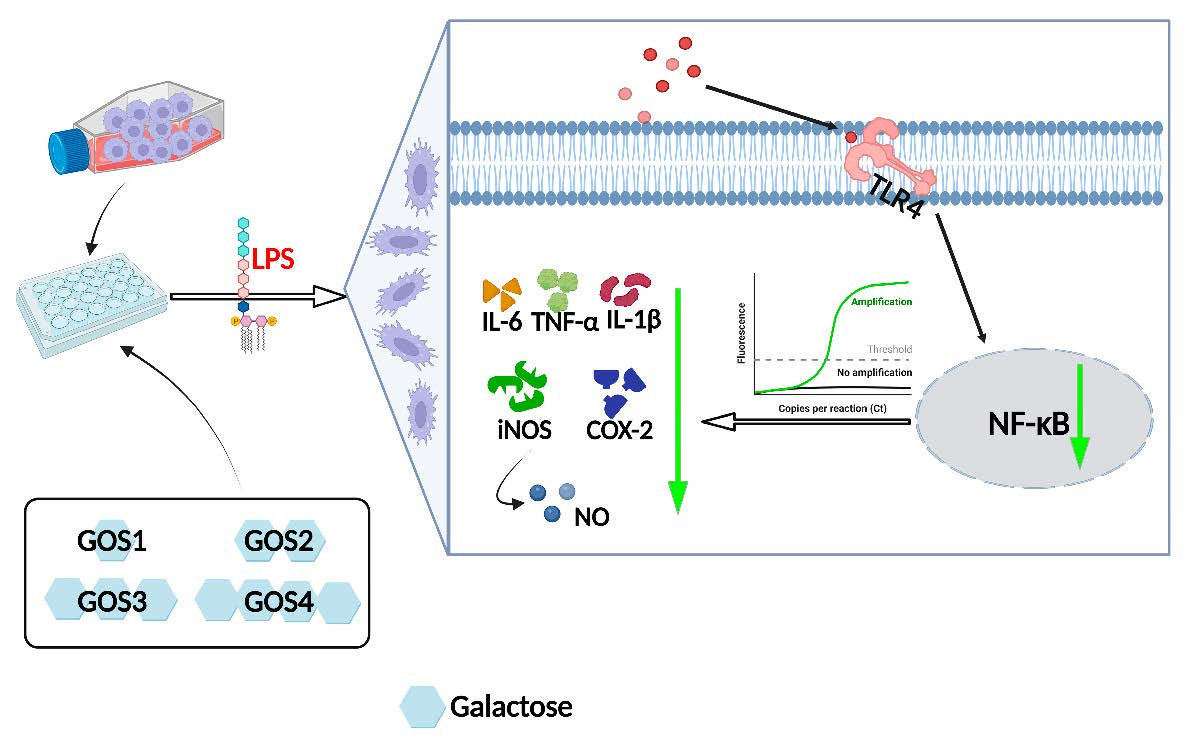
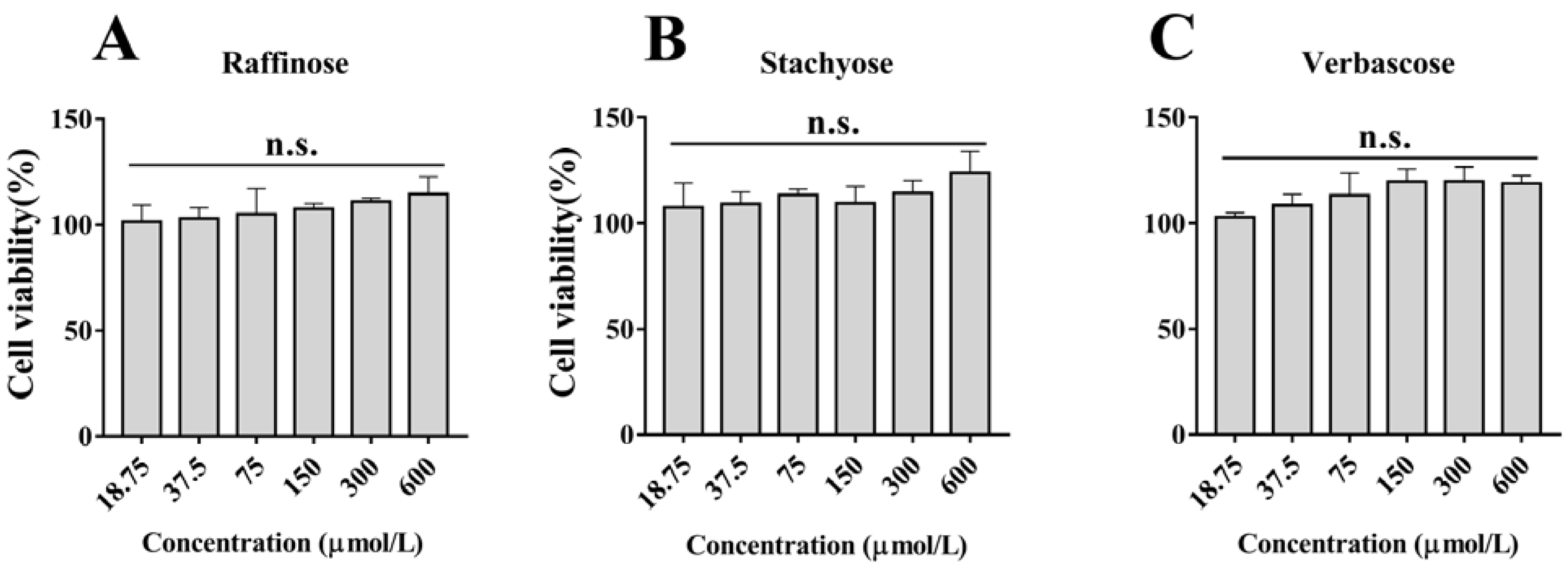
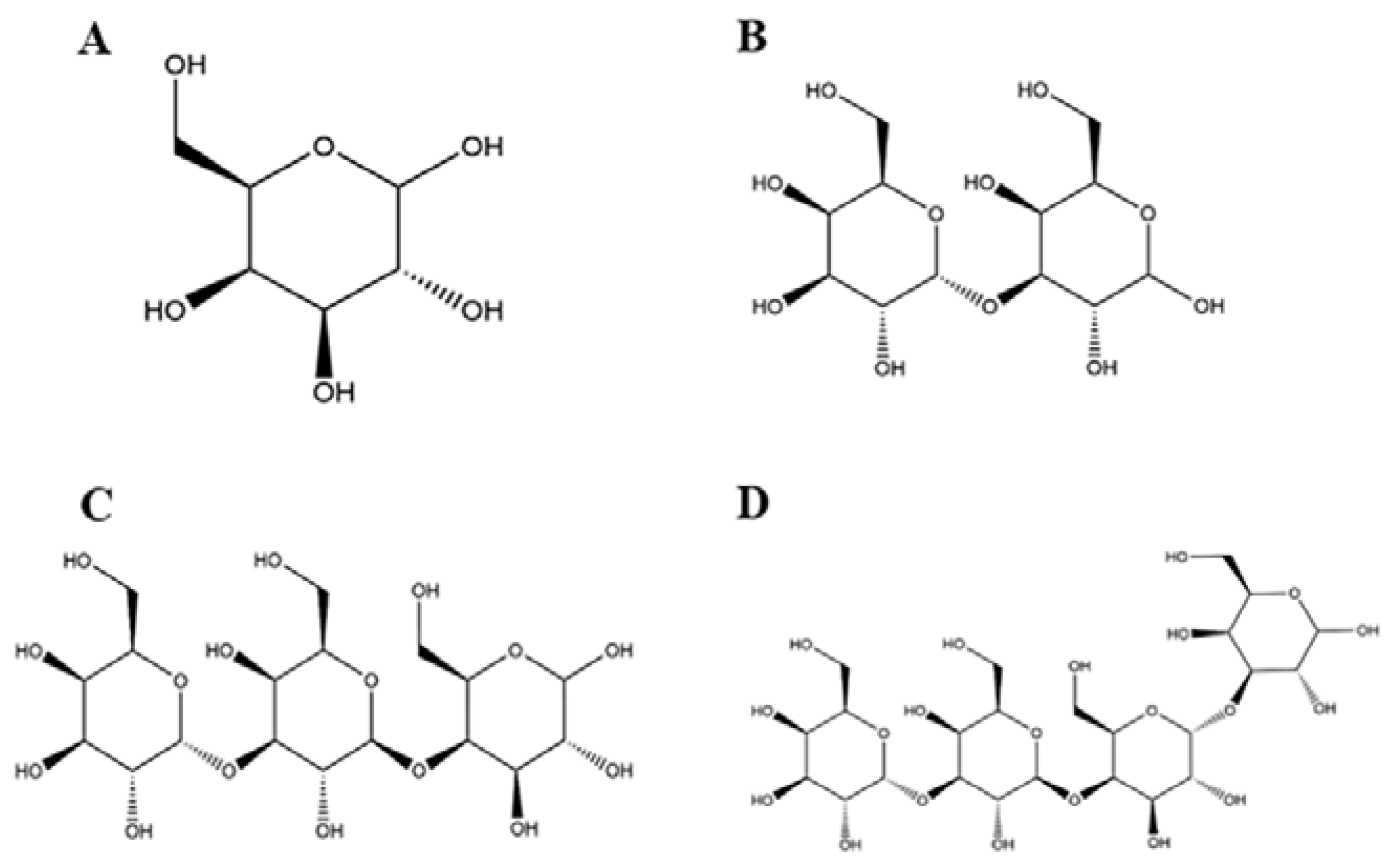
3.1. Effects of Natural GOS on Cell Viability
3.2. Effects of Natural GOS on NO, IL-1β, TNF-α, and IL-6 Secretion
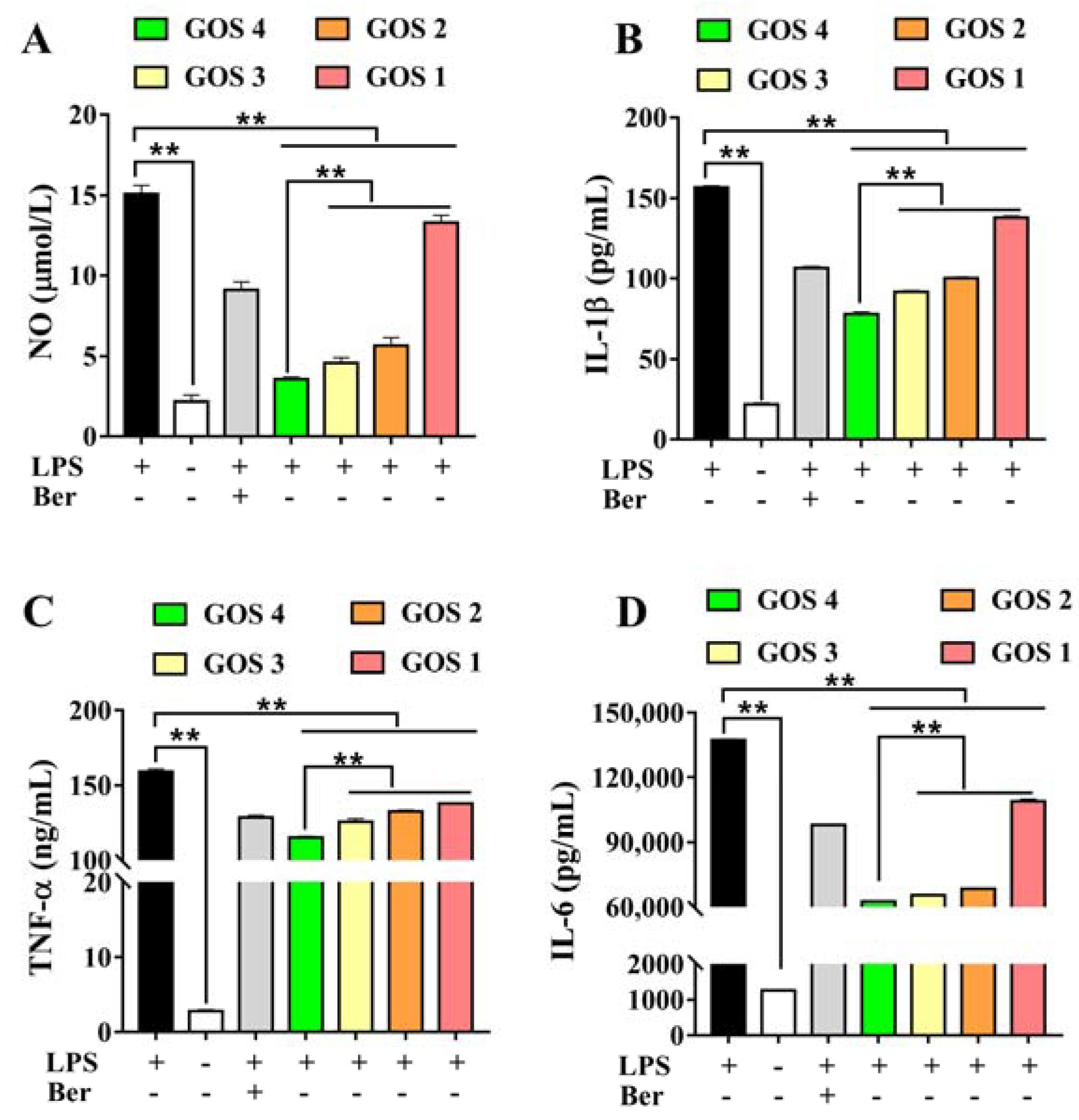
3.3. Effects of GOS1-4 on NO, IL-1β, TNF-α, and IL-6 levels
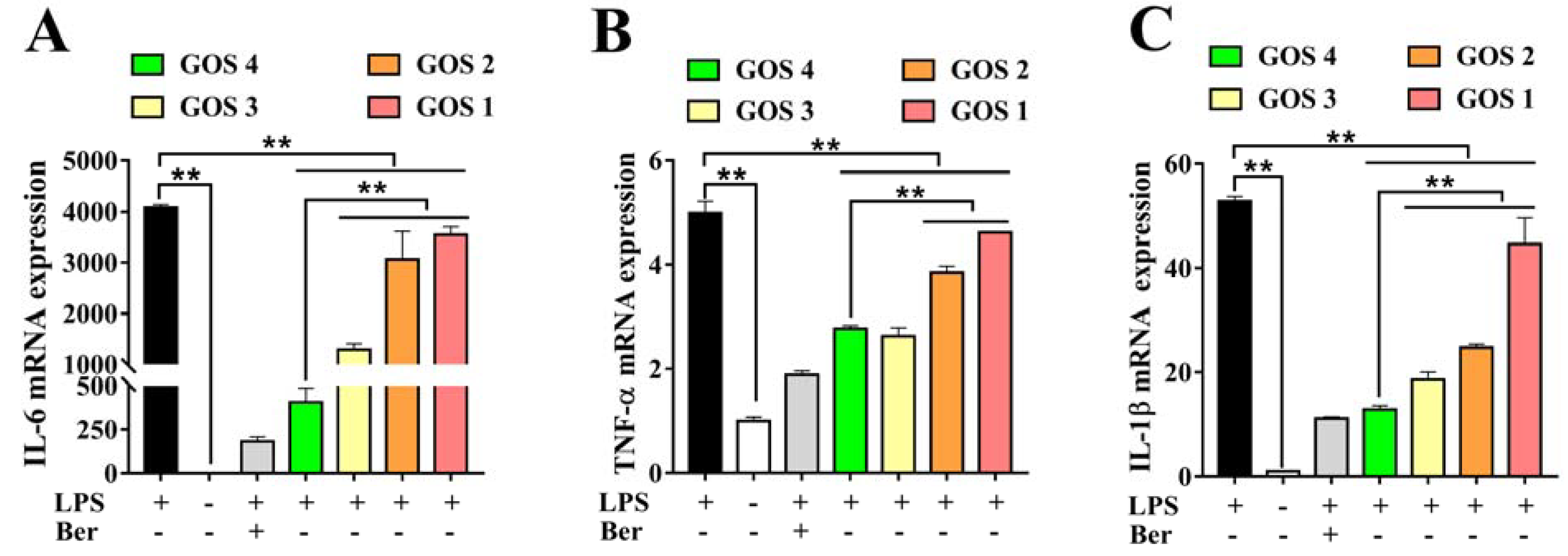
3.4. Effects of GOS1-4 on Pro-inflammatory Cytokine mRNA Expression in LPS-Induced RAW264.7 Macrophages
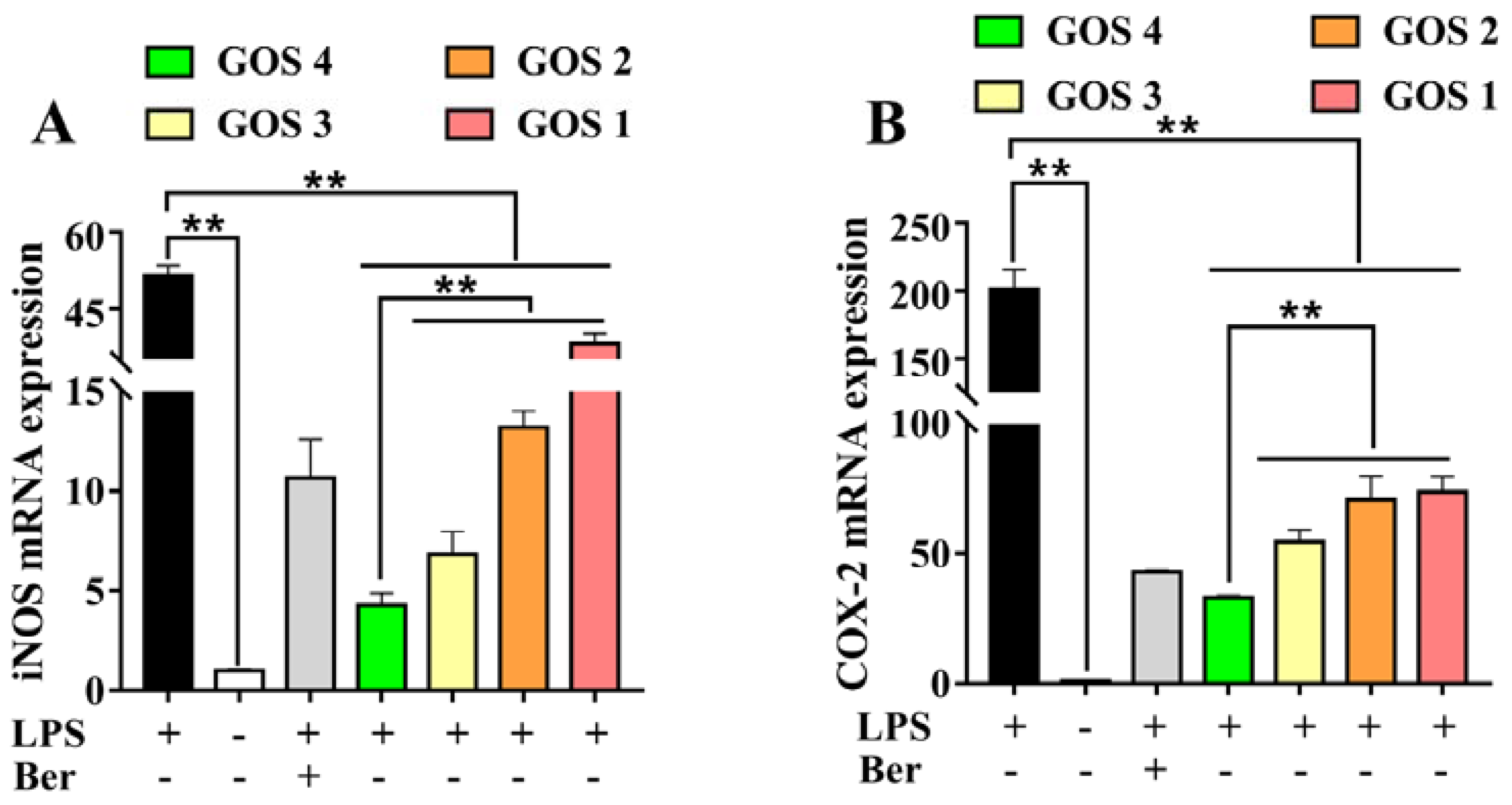
3.5. Effects of GOS1-4 on Pro-inflammatory Enzyme mRNA Expression in LPS-Induced RAW264.7 Macrophages
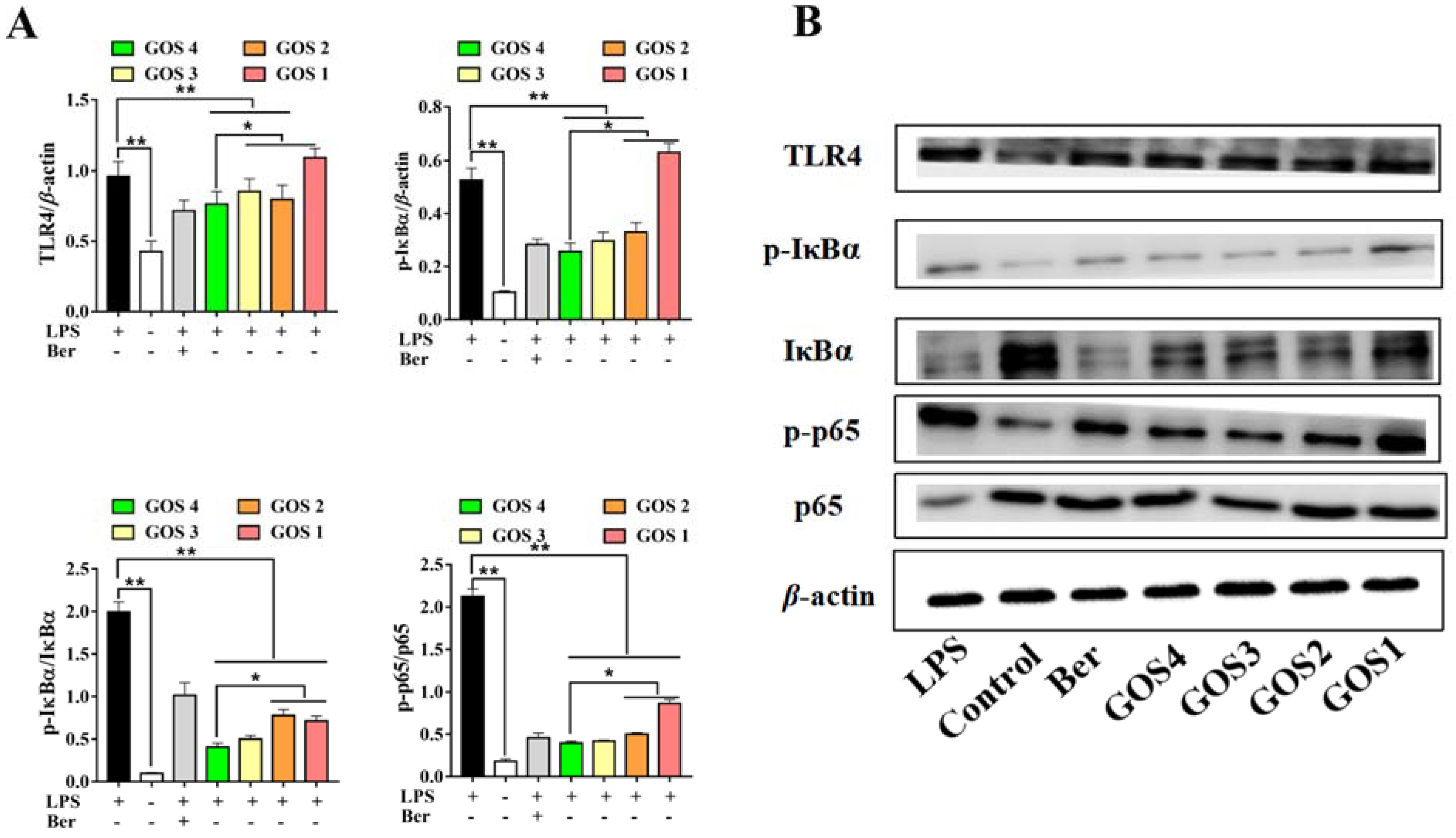
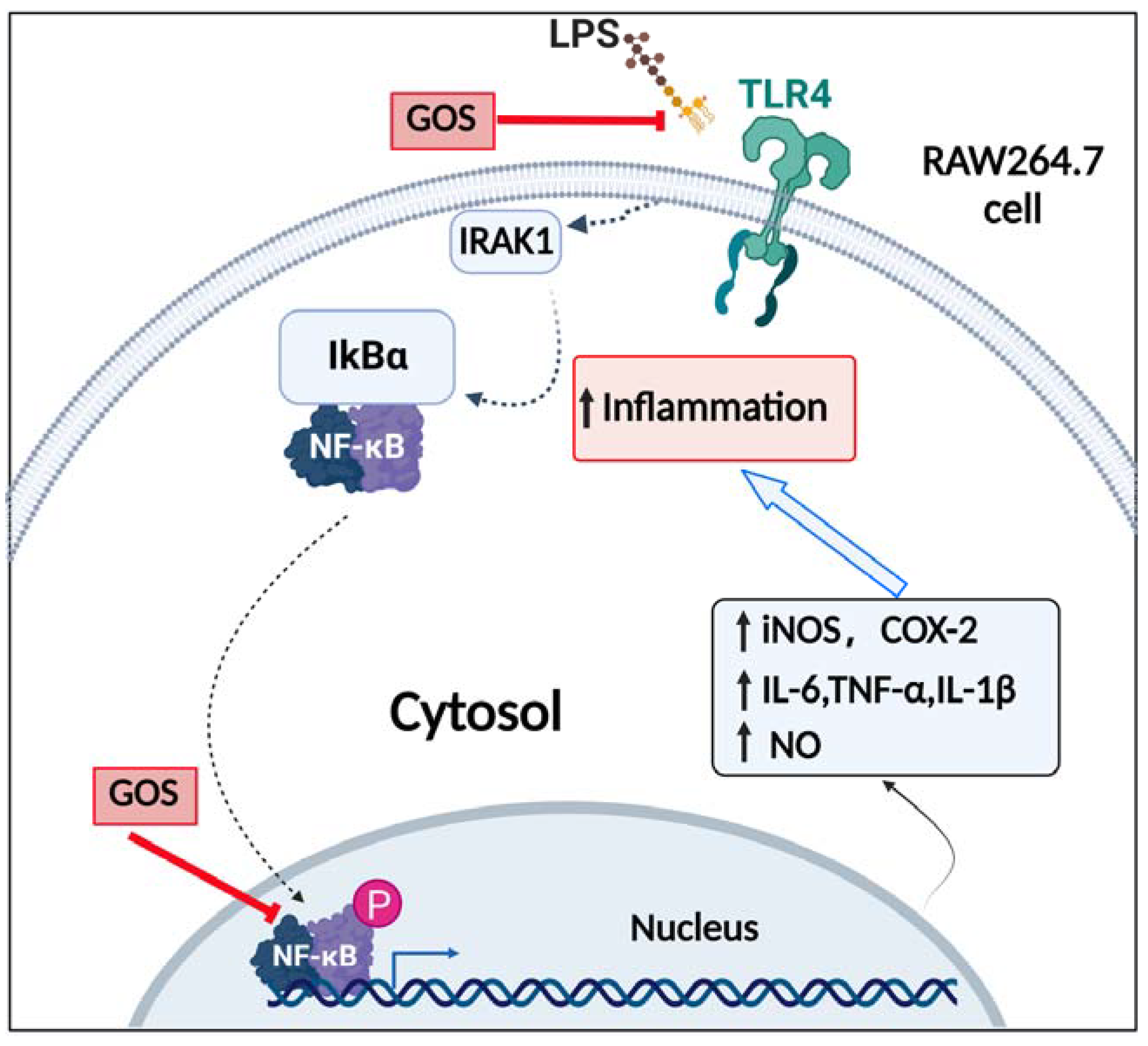
3.6. Potential of GOS to Modulate the TLR4/NF-κB Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Farias, D.; Araújo, F.; Neri-Numa, I.A.; Pastore, G.M. Prebiotics: Trends in food, health and technological applications—ScienceDirect. Trends Food Sci. Technol. 2019, 93, 23–35. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Q.; Goff, H.D.; Lapointe, G. Oligosaccharides: Structure, Function and Application. Encycl. Food Chem. 2019, 1, 202–207. [Google Scholar]
- Lamsal, B.P. Production, health aspects and potential food uses of dairy prebiotic galactooligosaccharides. J. Sci. Food Agric. 2012, 92, 2020–2028. [Google Scholar] [CrossRef]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Jennifer, C.; Krista, S.; Ali, K.; Jens, W. Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef]
- Mohajeri, M.H.; Brummer, R.; Rastall, R.A.; Weersma, R.K.; Harmsen, H.; Faas, M.; Eggersdorfer, M. The role of the microbiome for human health: From basic science to clinical applications. Eur. J. Nutr. 2018, 57, S1–S14. [Google Scholar] [CrossRef]
- Rigo-Adrover, M.; Knipping, K.; Garssen, J.; Van Limpt, K.; Knol, J.; Franch, À.; Castell, M.; Rodríguez-lagunas, M.; Pérez-Cano, F. Prevention of Rotavirus Diarrhea in Suckling Rats by a Specific Fermented Milk Concentrate with Prebiotic Mixture. Nutrients 2019, 11, 189. [Google Scholar] [CrossRef]
- Hirohisa, I.; Tatsuya, E.; Hirosuke, S.; Takeshi, M.; Eri, M.; Yuki, N.; Muneya, T.; Takashi, S.; Toshitaka, O.; Xiao, J.Z. The Combination of Bifidobacterium breve and Three Prebiotic Oligosaccharides Modifies Gut Immune and Endocrine Functions in Neonatal Mice. J. Nutr. 2019, 149, 344–353. [Google Scholar]
- Pan, P.; Oshima, K.; Huang, Y.W.; Agle, K.A.; Drobyski, W.R.; Chen, X.; Zhang, J.; Yearsley, M.M.; Yu, J.; Wang, L.S. Loss of FFAR2 promotes colon cancer by epigenetic dysregulation of inflammation suppressors. Int. J. Cancer 2018, 143, 886–896. [Google Scholar] [CrossRef]
- Lombó, F.; Fernández, J.; Moreno, F.J.; Olano, A.; Villar, C.J. A Galacto-Oligosaccharides Preparation Derived From Lactulose Protects Against Colorectal Cancer Development in an Animal Model. Front. Microbiol. 2018, 9, 2004. [Google Scholar]
- Wang, Z.Y.; Zeng, M.M.; Wang, Z.J.; Qin, F.; Chen, J.; He, Z.Y. Dietary Polyphenols to Combat Nonalcoholic Fatty Liver Disease via the Gut-Brain-Liver Axis: A Review of Possible Mechanisms. J. Agric. Food Chem. 2021, 69, 3585–3600. [Google Scholar] [CrossRef]
- Kim, Y.A.; Keogh, J.B.; Clifton, P.M. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr. Res. Rev. 2018, 31, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Chen, X.; Zhao, Z.J.; Liao, Y.; Zhou, T.; Xiang, Q. A potential link between plasma short-chain fatty acids, TNF-alpha level and disease progression in non-alcoholic fatty liver disease: A retrospective study. Exp. Ther. Med. 2022, 24, 598. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.E.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of Diet With Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology 2017, 153, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Tao, X.; Sun, Z.; Hao, W.; Wei, X. Galactooligosaccharides protects against DSS-induced murine colitis through regulating intestinal flora and inhibiting NF-κB pathway. Life Sci. 2019, 242, 117220. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z. Enzymatic Synthesis of Alpha—Galactooligosaccharides and Evaluation of Its Physiological Function and Structure-Activity Relationship. Master’s thesis, Nanjing Agricultural University, Nanjing, China, 2017. [Google Scholar]
- Panesar, P.S.; Kaur, R.; Singh, R.S.; Kennedy, J.F. Biocatalytic strategies in the production of galacto-oligosaccharides and its global status. Int. J. Biol. Macromol. 2018, 111, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P.K.; Sullivan, P.A.; Smart, J.B. Utilisation of galacto-oligosaccharides as selective substrates for growth by lactic acid bacteria including Bifidobacterium lactis DR10 and Lactobacillus rhamnosus DR20. Int. Dairy J. 2001, 11, 19–25. [Google Scholar] [CrossRef]
- Barboza, M.; Sela, D.A.; Pirim, C.; Locascio, R.G.; Lebrilla, C.B. Glycoprofiling Bifidobacterial Consumption of Galacto-Oligosaccharides by Mass Spectrometry Reveals Strain-Specific, Preferential Consumption of Glycans. Appl. Environ. Microbiol. 2009, 75, 7319–7325. [Google Scholar] [CrossRef]
- Wei, R.; Rong, C.; Xie, Q.; Wu, S.; Lin, T. Neuroprotective Effect of Optimized Yinxieling Formula in 6-OHDA-Induced Chronic Model of Parkinson’s Disease through the Inflammation Pathway. Evid. -Based Complement. Altern. Med. 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Xie, C.; Wang, S.; Cao, M.; Xiong, W.; Wu, L. (E)-9-Octadecenoic Acid Ethyl Ester Derived from Lotus Seedpod Ameliorates Inflammatory Responses by Regulating MAPKs and NF-κB Signalling Pathways in LPS-Induced RAW264.7 Macrophages. Evid. -Based Complement. Altern. Med. 2022, 2022, 6731360. [Google Scholar] [CrossRef]
- Li, L.-C.; Pan, Z.-H.; Ning, D.-S.; Fu, Y.-X. Anti-Inflammatory Effect of Simonsinol on Lipopolysaccharide Stimulated RAW264.7 Cells through Inactivation of NF-κB Signaling Pathway. Molecules 2020, 25, 3573. [Google Scholar] [CrossRef]
- Zeng, J.; Xu, Y.; Shi, Y.; Jiang, C. Inflammation Role in Sensory Neuropathy in Chinese Patients with Diabetes/Prediabetes. Clin. Neurol. Neurosurg. 2018, 166, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Angelotti, F.; Parma, A.; Cafaro, G.; Capecchi, R.; Puxeddu, I. One year in review 2017: Pathogenesis of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 368–378. [Google Scholar] [PubMed]
- Ren, Z.Y.; Huo, Y.L.; Zhang, Q.M.; Chen, S.F.; Lv, H.H.; Peng, L.L.; Wei, H.; Wan, C.X. Protective Effect of Lactiplantibacillus plantarum 1201 Combined with Galactooligosaccharide on Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Nutrients 2021, 13, 4441. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Wang, L.K.; Wu, H.Y.; Jiao, L. Effects of prebiotic galacto-oligosaccharide on postoperative cognitive dysfunction and neuroinflammation through targeting of the gut-brain axis. Bmc Anesthesiol. 2018, 18, 177. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Zhang, Y.H.; Li, W.H.; Yin, J.; Zhang, B.W.; Wang, J.; Wang, S. Differential responses on gut microbiota and microbial metabolome of 2’-fucosyllactose and galactooligosaccharide against DSS-induced colitis. Food Res. Int. 2022, 162, 112072. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Sang, Y.; Liu, W.; Yu, W.; Wang, X. Anti-Inflammatory Effects of Gingerol on Lipopolysaccharide-Stimulated RAW 264.7 Cells by Inhibiting NF-κB Signaling Pathway. Inflammation 2018, 41, 835–845. [Google Scholar] [PubMed]
- Zhang, T.; Cui, B.; Zhang, F. Progress of cytokines in treatment of inflammatory bowel disease. Chin. J. Gastroenterol. Hepatol. 2016, 25, 724–728. [Google Scholar]
- Chen, J.; Li, D.L.; Xie, L.N.; Ma, Y.R.; Liu, X. Synergistic anti-inflammatory effects of silibinin and thymol combination on LPS-induced RAW264.7 cells by inhibition of NF-κB and MAPK activation. Phytomedicine 2020, 78, 153309. [Google Scholar] [CrossRef]
- Li, M.; Dong, L.; Du, H.; Bao, Z.; Lin, S. Potential mechanisms underlying the protective effects of Tricholoma matsutake Singer peptides against LPS-induced inflammation in RAW264.7 macrophages. Food Chem. 2021, 353, 129452. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in LPS- Induced RAW264.7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef]
- Ko, W.; Kim, K.-W.; Quang, T.H.; Yoon, C.-S.; Kim, N.; Lee, H.; Kim, S.-C.; Woo, E.-R.; Kim, Y.-C.; Oh, H.; et al. Cudraflavanone B Isolated from the Root Bark of Cudrania tricuspidata Alleviates Lipopolysaccharide-Induced Inflammatory Responses by Downregulating NF-κB and ERK MAPK Signaling Pathways in RAW264.7 Macrophages and BV2 Microglia. Inflammation 2021, 44, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dao, T.T.; Hien, T.T.; Zhao, Y.M.; Shi, Y.P. Further sesquiterpenoids from the rhizomes of Homalomena occulta and their anti-inflammatory activity. Bioorganic Med. Chem. Lett. 2019, 29, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, P.; Hao, C.; Wu, L.; Wan, W.; Mao, X. Neoagaro-oligosaccharide monomers inhibit inflammation in LPS-stimulated macrophages through suppression of MAPK and NF-κB pathways. Sci. Rep. 2017, 7, 44252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.N.; Yin, L.Q.; Zhang, L.R.; Jiang, D.L.; Liu, L.; Ji, H. Chitoheptaose Promotes Heart Rehabilitation in a Rat Myocarditis Model by Improving Antioxidant, Anti-Inflammatory, and Antiapoptotic Properties. Oxidative Med. Cell. Longev. 2020, 2020, 2394704. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, C.; Zhang, L.; Wang, Y.; Chen, G.; Fan, J.; Jia, Y.; Yan, F.; Ning, C. Pectin oligosaccharides from fruit of Actinidia arguta: Structure-activity relationship of prebiotic and antiglycation potentials. Carbohydr. Polym. 2019, 217, 90–97. [Google Scholar] [CrossRef]
- Sheeba, M. Toll-Like Receptor 4 in Inflammation and Angiogenesis: A Double-Edged Sword. Front. Immunol. 2014, 5, 313. [Google Scholar]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J.; Jung, H.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-κB and MAPK signaling pathways in LPS-induced RAW264.7 macrophage cells. J. Ethnopharmacol. 2018, 231, 10–18. [Google Scholar] [CrossRef]
- Joh, E.H.; Kim, D.H. Kalopanaxsaponin A ameliorates experimental colitis in mice by inhibiting IRAK-1 activation in the NF-κB and MAPK pathways. Br. J. Pharmacol. 2011, 162, 1731–1742. [Google Scholar] [CrossRef]
- Jin, S.; Wang, J.; Chen, S.; Jiang, A.; Jiang, M.; Su, Y.; Yan, W.; Xu, Y.; Gong, G. A novel limonin derivate modulates inflammatory response by suppressing the TLR4/NF-κB signalling pathway. Biomed. Pharmacother. 2018, 100, 501–508. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.J.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of iNOS and COX-2 expression by flavokawain A via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 58, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.X.; Gao, H.W.; Sun, W.; Yu, J.; Hu, H.; Xu, Q.M.; Chen, X.P.; Nepetoidin, B. A Natural Product, Inhibits LPS-stimulated Nitric Oxide Production via Modulation of iNOS Mediated by NF-kappa B/MKP-5 Pathways. Phytother. Res. 2017, 31, 1072–1077. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| iNOS | CAGCGGAGTGACGGCAAACAT | GCAAGACCAGAGGCAGCACATC |
| IL-1β | ACCTGGGCTGTCCTGATGAGAG | TGTTGATGTGCTGCTGCGAGAT |
| COX-2 | CTGGTGCCTGGTCTGATGATGTATG | TCTCCTATGAGTATGAGTCTGCTGGTT |
| TNF-α | TGGAACTGGCAGAAGAGGCACT | AGAGGCTGAGACATAGGCACCG |
| IL-6 | GTTCTCTGGGAAATCGTGGA | GGAAATTGGGGTAGGAAGGA |
| GAPDH | ACTCCACTCACGGCAAATTC | GTCATGAGCCCTTCCACAAT |
| Groups | NO (μmol/L) | IL-1β (pg/mL) | TNF-α (ng/mL) | IL-6 (ng/mL) |
|---|---|---|---|---|
| Control | 1.18 ± 0.61 f | 24.04 ± 0.20 f | 1.49 ± 0.12 f | 0.24 ± 0.01 f |
| LPS | 14.14 ± 0.19 a | 88.09 ± 0.26 a | 121.64 ± 1.29 a | 71.48 ± 0.12 a |
| Ber | 12.91 ± 0.47 b | 65.66 ± 0.18 b | 85.83 ± 0.28 b | 55.84 ± 0.21 b |
| Raffinose | 10.83 ± 0.12 c | 65.17 ± 0.29 c | 97.18 ± 0.29 c | 67.16 ± 0.23 c |
| Stachyose | 10.30 ± 0.18 d | 64.13 ± 0.11 d | 93.90 ± 0.24 d | 60.90 ± 0.13 d |
| Verbascose | 7.21 ± 0.08 e | 55.32 ± 0.29 e | 92.50 ± 1.02 e | 56.80 ± 0.22 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Hao, B.; Pang, D.; Li, Q.; Li, E.; Yang, Q.; Zou, Y.; Liao, S.; Liu, F. Diverse Galactooligosaccharides Differentially Reduce LPS-Induced Inflammation in Macrophages. Foods 2022, 11, 3973. https://doi.org/10.3390/foods11243973
Sun C, Hao B, Pang D, Li Q, Li E, Yang Q, Zou Y, Liao S, Liu F. Diverse Galactooligosaccharides Differentially Reduce LPS-Induced Inflammation in Macrophages. Foods. 2022; 11(24):3973. https://doi.org/10.3390/foods11243973
Chicago/Turabian StyleSun, Congcong, Bifang Hao, Daorui Pang, Qian Li, Erna Li, Qiong Yang, Yuxiao Zou, Sentai Liao, and Fan Liu. 2022. "Diverse Galactooligosaccharides Differentially Reduce LPS-Induced Inflammation in Macrophages" Foods 11, no. 24: 3973. https://doi.org/10.3390/foods11243973
APA StyleSun, C., Hao, B., Pang, D., Li, Q., Li, E., Yang, Q., Zou, Y., Liao, S., & Liu, F. (2022). Diverse Galactooligosaccharides Differentially Reduce LPS-Induced Inflammation in Macrophages. Foods, 11(24), 3973. https://doi.org/10.3390/foods11243973