The Impact of [C16Pyr][Amp] on the Aggressiveness in Breast and Prostate Cancer Cell Lines

 , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. [C16Pyr][Amp] Synthetic Compound Displayed Low Half-Maximal Inhibitory Concentration (IC50) Values
2.2. [C16Pyr][Amp] Exhibited Higher Selectivity toward Tumor Cells Compared to Cisplatin
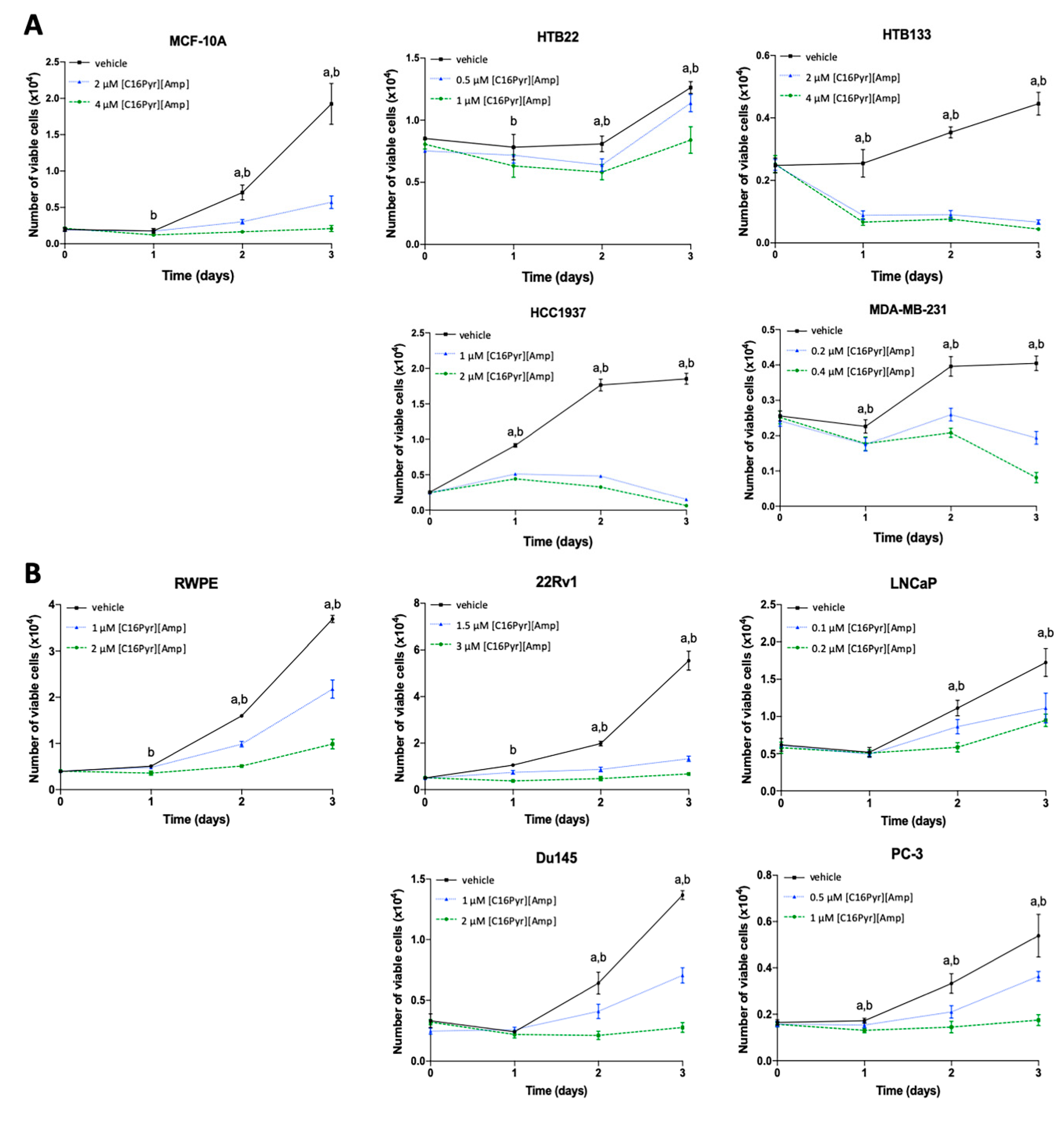
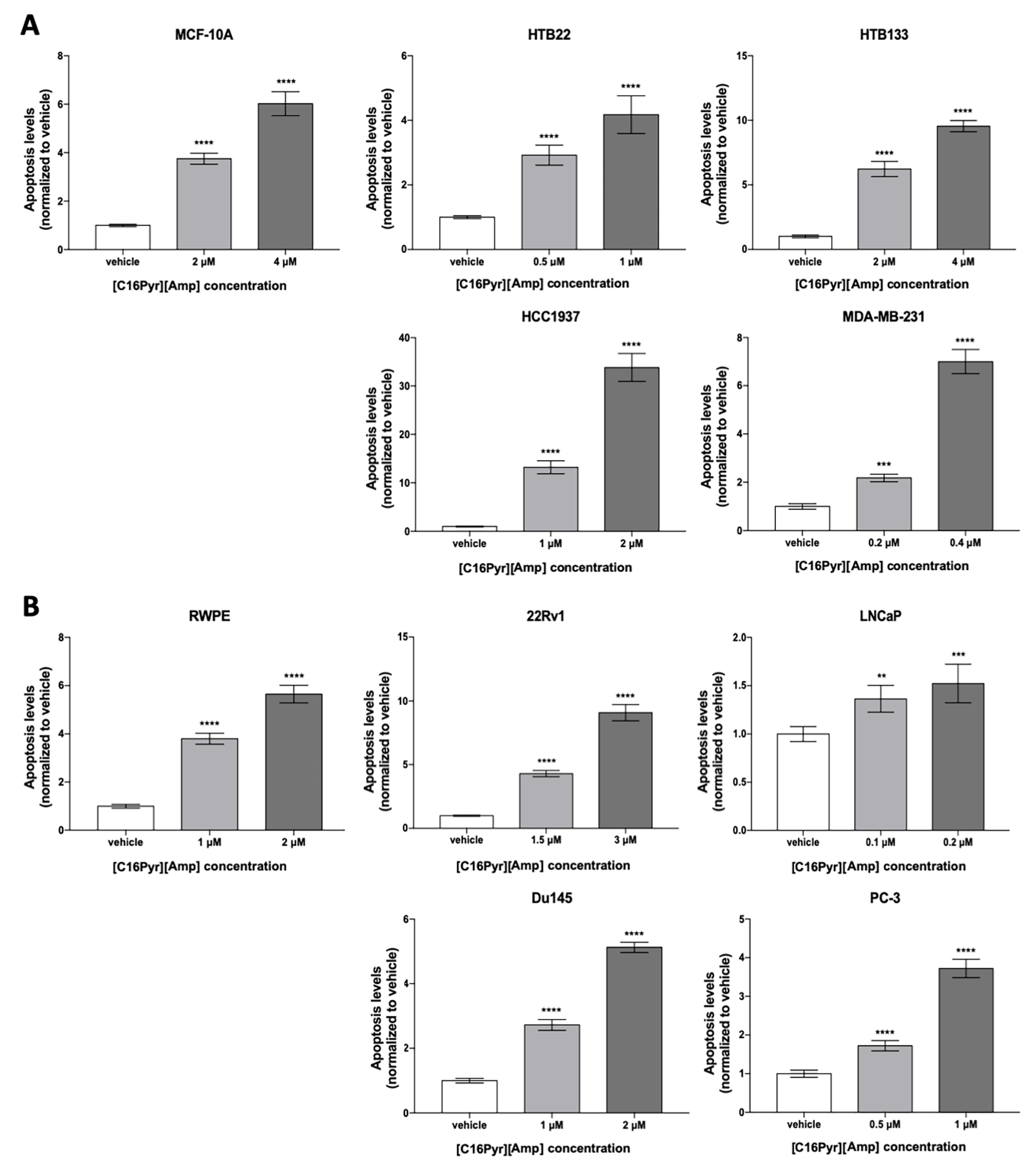
2.3. [C16Pyr][Amp] Decreased Tumor Cell Viability and Increased Apoptosis Levels
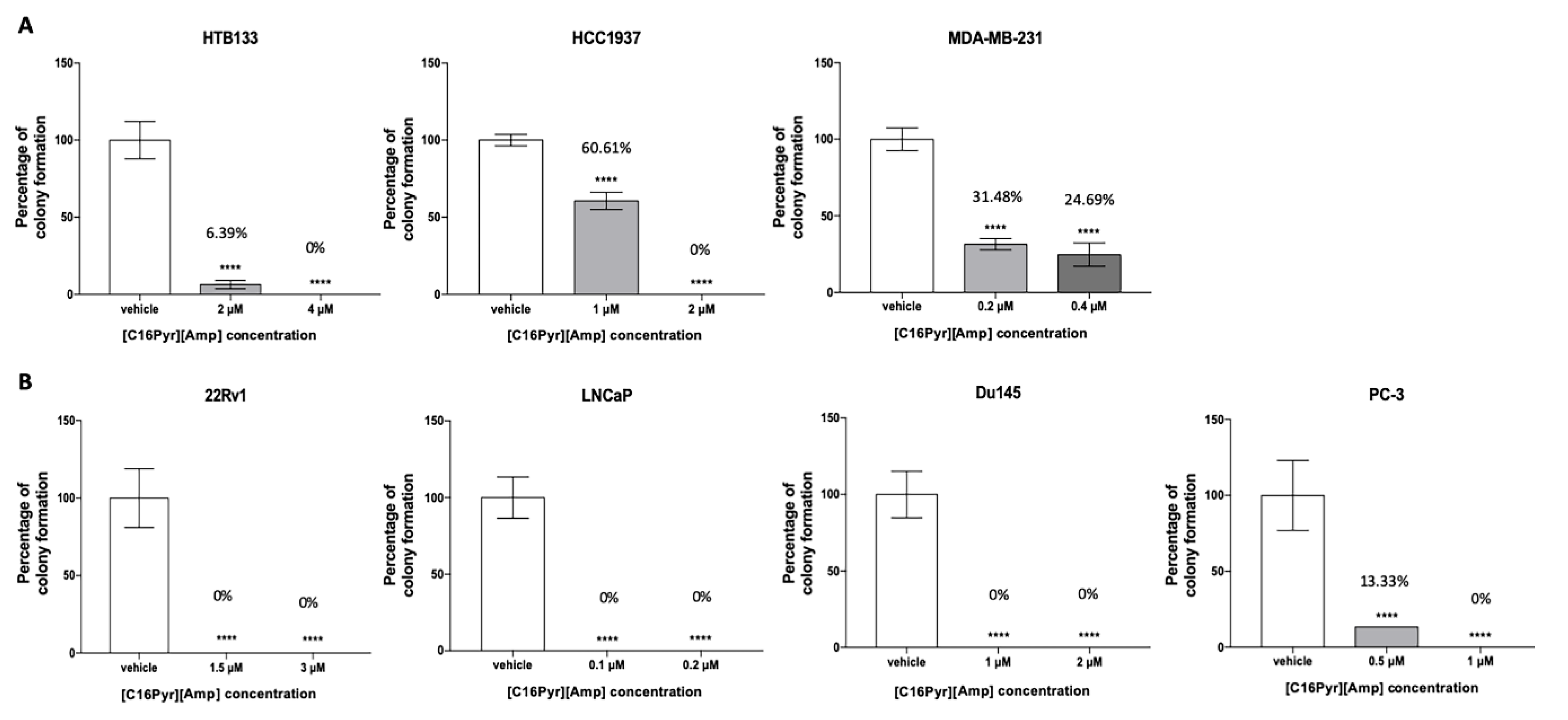
2.4. [C16Pyr][Amp] Reduced the Colony Formation Capacity of Tumor Cells
2.5. CPT2, LDHA, MCM,2 and SKP2 Gene Expression Is Downregulated in Tumor Cells
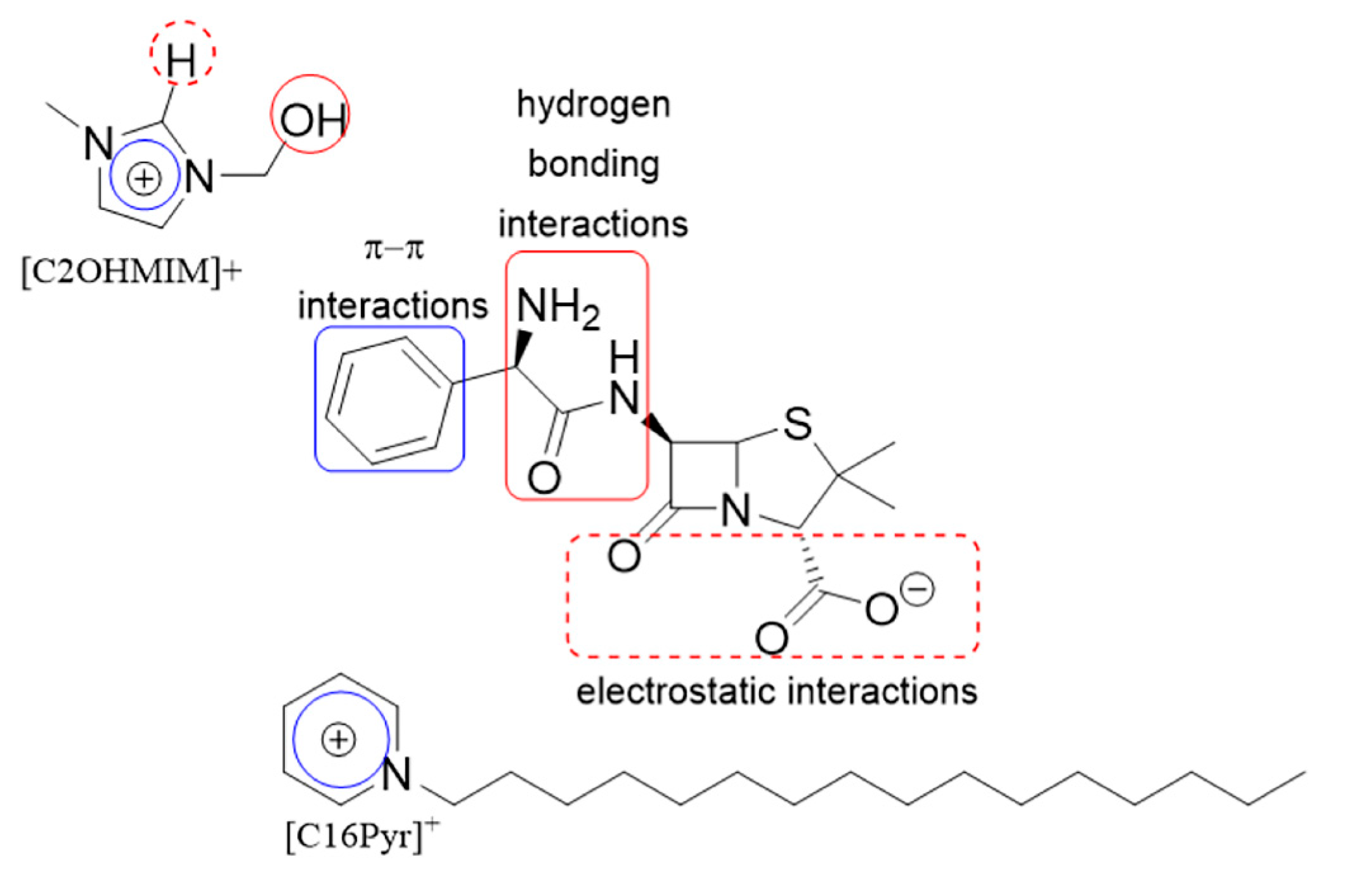
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Compound Formulation
4.3. Cell Viability Assay, IC50, and Selectivity Index Assessment
4.4. Apoptosis Assay
4.5. Cytotoxicity Assay
4.6. Colony Formation Assay
4.7. RNA Quantification, Reverse Transcription, and Real-Time PCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | One-way analysis of variance |
| BrCa | Breast cancer |
| cDNA | Complementary DNA |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl Sulfoxide |
| DSC | Differential scanning calorimetry |
| EGF | Epidermal Growth Factor |
| FAO | Fatty acid oxidation |
| FBS | Fetal bovine serum |
| GDC | DNA genomic contamination |
| IC50 | Half-maximal inhibitory concentration |
| IL | Ionic liquid |
| LDH | Lactate dehydrogenase |
| MEM | Minimum essential media |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium-bromide |
| OD | Optical Density |
| PCa | Prostate cancer |
| PPC | Real-time PCR efficiency |
| RTC | First strand synthesis |
| SFM | Serum-Free Medium |
| SI | Selectivity index |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuur, E.R.; DeAndrade, J.P. Breast Cancer: Molecular Mechanisms, Diagnosis, and Treatment. In International Manual of Oncology Practice; Springer: Berlin/Heidelberg, Germany, 2015; pp. 155–200. [Google Scholar]
- Lakhani, S.R. WHO Classification of Tumours of the Breast; International Agency for Research on Cancer: Lyon, France, 2012. [Google Scholar]
- Putti, T.C.; El-Rehim, D.M.; Rakha, E.A.; Paish, C.E.; Lee, A.H.; Pinder, S.E.; Ellis, I.O. Estrogen receptor-negative breast carcinomas: A review of morphology and immunophenotypical analysis. Mod. Pathol. 2005, 18, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Szostakowska, M.; Trebinska-Stryjewska, A.; Grzybowska, E.A.; Fabisiewicz, A. Resistance to endocrine therapy in breast cancer: Molecular mechanisms and future goals. Breast Cancer Res. Treat. 2019, 173, 489–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, F.; Costa, A.; Senkus, E.; Aapro, M.; Andre, F.; Barrios, C.H.; Bergh, J.; Bhattacharyya, G.; Biganzoli, L.; Cardoso, M.J.; et al. 3rd ESO-ESMO International Consensus Guidelines for Advanced Breast Cancer (ABC 3). Ann. Oncol. 2017, 28, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Jinno, H.; Murata, T.; Seki, T.; Takahashi, M.; Hayashida, T.; Kameyama, K.; Kitagawa, Y. Prognostic implications of receptor discordance between primary and recurrent breast cancer. Int. J. Clin. Oncol. 2015, 20, 701–708. [Google Scholar] [CrossRef]
- Heidenreich, A.; Bastian, P.J.; Bellmunt, J.; Bolla, M.; Joniau, S.; van der Kwast, T.; Mason, M.; Matveev, V.; Wiegel, T.; Zattoni, F.; et al. EAU guidelines on prostate cancer. Part II: Treatment of advanced, relapsing, and castration-resistant prostate cancer. Eur. Urol. 2014, 65, 467–479. [Google Scholar] [CrossRef]
- Watson, P.A.; Arora, V.K.; Sawyers, C.L. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat. Rev. Cancer 2015, 15, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Saad, F.; Miller, K. Treatment options in castration-resistant prostate cancer: Current therapies and emerging docetaxel-based regimens. Urol. Oncol. 2014, 32, 70–79. [Google Scholar] [CrossRef]
- Petrylak, D.P.; Tangen, C.M.; Hussain, M.H.; Lara, P.N., Jr.; Jones, J.A.; Taplin, M.E.; Burch, P.A.; Berry, D.; Moinpour, C.; Kohli, M.; et al. Docetaxel and estramustine compared with mitoxantrone and prednisone for advanced refractory prostate cancer. N. Engl. J. Med. 2004, 351, 1513–1520. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, R.; Costa-Rodrigues, J.; Fernandes, M.H.; Santos, M.M.; Marrucho, I.M.; Rebelo, L.P.; Prudencio, C.; Noronha, J.P.; Petrovski, Z.; Branco, L.C. Antitumor Activity of Ionic Liquids Based on Ampicillin. ChemMedChem 2015, 10, 1480–1483. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.; Pinheiro, C.; Fernandes, R.; Noronha, J.P.; Prudencio, C. Antimicrobial activity of quinoxaline 1,4-dioxide with 2- and 3-substituted derivatives. Microbiol. Res. 2014, 169, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Egorova, K.S.; Gordeev, E.G.; Ananikov, V.P. Biological Activity of Ionic Liquids and Their Application in Pharmaceutics and Medicine. Chem. Rev. 2017, 117, 7132–7189. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, B.F. Antimicrobial ionic liquids. In Ionic Liquids: Applications and Perspectives; Kokorin, P.A., Ed.; InTech: London, UK, 2011; pp. 587–604. [Google Scholar]
- Ferraz, R.; Branco, L.C.; Prudencio, C.; Noronha, J.P.; Petrovski, Z. Ionic liquids as active pharmaceutical ingredients. Chem. Med. Chem. 2011, 6, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Marrucho, I.M.; Branco, L.C.; Rebelo, L.P. Ionic liquids in pharmaceutical applications. Ann. Rev. Chem. Biomol. Eng. 2014, 5, 527–546. [Google Scholar] [CrossRef]
- Smiglak, M.; Pringle, J.M.; Lu, X.; Han, L.; Zhang, S.; Gao, H.; MacFarlane, D.R.; Rogers, R.D. Ionic liquids for energy, materials, and medicine. Chem. Commun. 2014, 50, 9228–9250. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, S.V. Ionic Liquid Applications: Pharmaceuticals, Therapeutics, and Biotechnology; American Chemical Society: Washington, DC, USA, 2010; pp. i–v. [Google Scholar] [CrossRef] [Green Version]
- Plechkova, N.V.; Seddon, K.R. Applications of ionic liquids in the chemical industry. Chem. Soc. Rev. 2008, 37, 123–150. [Google Scholar] [CrossRef]
- Pereira, J.A.; Pessoa, A.M.; Cordeiro, M.N.; Fernandes, R.; Prudencio, C.; Noronha, J.P.; Vieira, M. Quinoxaline, its derivatives and applications: A State of the Art review. Eur. J. Med. Chem. 2015, 97, 664–672. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.; Coelho, P.; Soares, R.; Prudencio, C.; Vieira, M. Quinoxaline-1,4-dioxide derivatives inhibitory action in melanoma and brain tumor cells. Future Med. Chem. 2019, 11, 645–657. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573. [Google Scholar] [CrossRef]
- Rabik, C.A.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat Rev. 2007, 33, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, P.; Carmo-Fonseca, M. Molecular mechanisms involved in cisplatin cytotoxicity. Cell Mol. Life Sci. 2000, 57, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Caparica, R.; Lambertini, M.; de Azambuja, E. How I treat metastatic triple-negative breast cancer. ESMO Open. 2019, 4, e000504. [Google Scholar] [CrossRef] [Green Version]
- Gronwald, J.; Byrski, T.; Lubinski, J.; Narod, S.A. Cisplatin in breast cancer treatment in BRCA1 carriers. Hered Cancer Clin. Pract. 2012, 10, A17. [Google Scholar] [CrossRef] [Green Version]
- Petrelli, F.; Barni, S.; Bregni, G.; de Braud, F.; Di Cosimo, S. Platinum salts in advanced breast cancer: A systematic review and meta-analysis of randomized clinical trials. Breast Cancer Res. Treat 2016, 160, 425–437. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Cisplatin in Castration Resistant Prostate Cancer (NCT03275857). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT03275857 (accessed on 9 May 2020).
- ClinicalTrials.gov. Gemcitabine Hydrochloride and Cisplatin With or Without Bevacizumab in Treating Patients With Advanced Urinary Tract Cancer (NCT00942331). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT00942331 (accessed on 9 May 2020).
- Qazi, R.; Khandekar, J. Phase II study of cisplatin for metastatic prostatic carcinoma. An Eastern Cooperative Oncology Group study. Am. J. Clin. Oncol. 1983, 6, 203–205. [Google Scholar] [CrossRef]
- Loening, S.A.; Beckley, S.; Brady, M.F.; Chu, T.M.; deKernion, J.B.; Dhabuwala, C.; Gaeta, J.F.; Gibbons, R.P.; McKiel, C.F.; McLeod, D.G.; et al. Comparison of estramustine phosphate, methotrexate and cis-platinum in patients with advanced, hormone refractory prostate cancer. J. Urol. 1983, 129, 1001–1006. [Google Scholar] [CrossRef]
- Yagoda, A.; Petrylak, D. Cytotoxic chemotherapy for advanced hormone-resistant prostate cancer. Cancer 1993, 71, 1098–1109. [Google Scholar] [CrossRef]
- Buonerba, C.; Federico, P.; D’Aniello, C.; Rescigno, P.; Cavaliere, C.; Puglia, L.; Ferro, M.; Altieri, V.; Perdonà, S.; De Placido, S.; et al. Phase II trial of cisplatin plus prednisone in docetaxel-refractory castration-resistant prostate cancer patients. Cancer Chemother. Pharmacol. 2011, 67, 1455–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ClinicalTrials.gov. Combination Chemotherapy Plus Peripheral Stem Cell Transplantation in Treating Patients With Metastatic Prostate Cancer (NCT00003400). 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT00003400 (accessed on 9 May 2020).
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostova, I. Platinum complexes as anticancer agents. Recent Patents Anti Cancer Drug Discov. 2006, 1, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, U.; Hamilton, G. A better platinum-based anticancer drug yet to come? Anti Cancer Agents Med. Chem. Form. Curr. Med. Chem. Anti Cancer Agents 2010, 10, 293–301. [Google Scholar] [CrossRef]
- Chan, F.K.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. Methods Mol. Biol. 2013, 979, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The soft agar colony formation assay. J. Vis. Exp. 2014, e51998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.R.; Costa-Rodrigues, J.; Fernandes, M.H.; Ferraz, R.; Prudêncio, C. The anticancer potential of ionic liquids. ChemMedChem 2017, 12, 11–18. [Google Scholar] [CrossRef]
- Jovanovic-Santa, S.; Kojic, V.; Atlagic, K.; Tot, A.; Vranes, M.; Gadzuric, S.; Karaman, M. Anticancer and antimicrobial properties of imidazolium based ionic liquids with salicylate anion. J. Serb. Chem. Soc. 2020, 85, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, S.V.; Kumar, V. A profile of the in vitro anti-tumor activity of imidazolium-based ionic liquids. Bioorg. Med. Chem. Lett. 2010, 20, 581–585. [Google Scholar] [CrossRef]
- Galluzzi, L.; Aaronson, S.A.; Abrams, J.; Alnemri, E.S.; Andrews, D.W.; Baehrecke, E.H.; Bazan, N.G.; Blagosklonny, M.V.; Blomgren, K.; Borner, C.; et al. Guidelines for the use and interpretation of assays for monitoring cell death in higher eukaryotes. Cell Death Differ. 2009, 16, 1093–1107. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.R.; Moshikur, R.M.; Wakabayashi, R.; Tahara, Y.; Kamiya, N.; Moniruzzaman, M.; Goto, M. Ionic-Liquid-Based Paclitaxel Preparation: A New Potential Formulation for Cancer Treatment. Mol. Pharm. 2018, 15, 2484–2488. [Google Scholar] [CrossRef]
- Rahimi, M.; Shafiei-Irannejad, V.; Safa, K.D.; Salehi, R. Multi-branched ionic liquid-chitosan as a smart and biocompatible nano-vehicle for combination chemotherapy with stealth and targeted properties. Carbohydr. Polym. 2018, 196, 299–312. [Google Scholar] [CrossRef]
- Tang, W.; Liu, B.; Wang, S.; Liu, T.; Fu, C.; Ren, X.; Tan, L.; Duan, W.; Meng, X. Doxorubicin-loaded ionic liquid–polydopamine nanoparticles for combined chemotherapy and microwave thermal therapy of cancer. RSC Adv. 2016, 6, 32434–32440. [Google Scholar] [CrossRef]
- Heger, J.I.; Froehlich, K.; Pastuschek, J.; Schmidt, A.; Baer, C.; Mrowka, R.; Backsch, C.; Schleußner, E.; Markert, U.R.; Schmidt, A. Human serum alters cell culture behavior and improves spheroid formation in comparison to fetal bovine serum. Exp. Cell Res. 2018, 365, 57–65. [Google Scholar] [CrossRef]
- Ledur, P.F.; Onzi, G.R.; Zong, H.; Lenz, G. Culture conditions defining glioblastoma cells behavior: What is the impact for novel discoveries? Oncotarget 2017, 8, 69185–69197. [Google Scholar] [CrossRef] [Green Version]
- Ivankovic, T.; Hrenovic, J. Surfactants in the environment. Arh. Hig. Rada Toksikol. 2010, 61, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Bubalo, M.C.; Radosevic, K.; Redovnikovic, I.R.; Halambek, J.; Srcek, V.G. A brief overview of the potential environmental hazards of ionic liquids. Ecotoxicol. Environ. Saf. 2014, 99, 1–12. [Google Scholar] [CrossRef]
- Garcia, M.T.; Campos, E.; Ribosa, I. Biodegradability and ecotoxicity of amine oxide based surfactants. Chemosphere 2007, 69, 1574–1578. [Google Scholar] [CrossRef]
- Kumar, V.; Malhotra, S.V. Study on the potential anti-cancer activity of phosphonium and ammonium-based ionic liquids. Bioorg. Med. Chem. Lett. 2009, 19, 4643–4646. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Smith, S.M.; Wunder, M.B.; Norris, D.A.; Shellman, Y.G. A simple protocol for using a LDH-based cytotoxicity assay to assess the effects of death and growth inhibition at the same time. PLoS ONE 2011, 6, e26908. [Google Scholar] [CrossRef]
- Kabakov, A.E.; Gabai, V.L. Cell Death and Survival Assays. Methods Mol. Biol. 2018, 1709, 107–127. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, N.K.; Attri, P.; Kaushik, N.; Choi, E.H. Synthesis and antiproliferative activity of ammonium and imidazolium ionic liquids against T98G brain cancer cells. Molecules 2012, 17, 13727–13739. [Google Scholar] [CrossRef]
- Bonnefont, J.P.; Djouadi, F.; Prip-Buus, C.; Gobin, S.; Munnich, A.; Bastin, J. Carnitine palmitoyltransferases 1 and 2: Biochemical, molecular and medical aspects. Mol. Asp. Med. 2004, 25, 495–520. [Google Scholar] [CrossRef]
- Melone, M.A.B.; Valentino, A.; Margarucci, S.; Galderisi, U.; Giordano, A.; Peluso, G. The carnitine system and cancer metabolic plasticity. Cell Death Dis. 2018, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Viale, A.; Pettazzoni, P.; Lyssiotis, C.A.; Ying, H.; Sánchez, N.; Marchesini, M.; Carugo, A.; Green, T.; Seth, S.; Giuliani, V.; et al. Oncogene ablation-resistant pancreatic cancer cells depend on mitochondrial function. Nature 2014, 514, 628–632. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Daniels, G.; Lee, P.; Monaco, M.E. Lipid metabolism in prostate cancer. Am. J. Clin. Exp. Urol. 2014, 2, 111–120. [Google Scholar]
- Zhao, D.; Xiong, Y.; Lei, Q.-Y.; Guan, K.-L. LDH-A acetylation: Implication in cancer. Oncotarget 2013, 4, 802–803. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Le, A.; Cooper, C.R.; Gouw, A.M.; Dinavahi, R.; Maitra, A.; Deck, L.M.; Royer, R.E.; Vander Jagt, D.L.; Semenza, G.L.; Dang, C.V. Inhibition of lactate dehydrogenase A induces oxidative stress and inhibits tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 2037–2042. [Google Scholar] [CrossRef] [Green Version]
- Sheng, S.L.; Liu, J.J.; Dai, Y.H.; Sun, X.G.; Xiong, X.P.; Huang, G. Knockdown of lactate dehydrogenase A suppresses tumor growth and metastasis of human hepatocellular carcinoma. FEBS J. 2012, 279, 3898–3910. [Google Scholar] [CrossRef]
- Xian, Z.Y.; Liu, J.M.; Chen, Q.K.; Chen, H.Z.; Ye, C.J.; Xue, J.; Yang, H.Q.; Li, J.L.; Liu, X.F.; Kuang, S.J. Inhibition of LDHA suppresses tumor progression in prostate cancer. Tumour Biol. 2015, 36, 8093–8100. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Chun, J.; Pan, C.; Alesi, G.N.; Li, D.; Magliocca, K.R.; Kang, Y.; Chen, Z.G.; Shin, D.M.; Khuri, F.R.; et al. Phosphorylation-mediated activation of LDHA promotes cancer cell invasion and tumour metastasis. Oncogene 2017, 36, 3797–3806. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate dehydrogenase A: A key player in carcinogenesis and potential target in cancer therapy. Cancer Med. 2018, 7, 6124–6136. [Google Scholar] [CrossRef] [Green Version]
- Fantin, V.R.; St-Pierre, J.; Leder, P. Attenuation of LDH-A expression uncovers a link between glycolysis, mitochondrial physiology, and tumor maintenance. Cancer Cell 2006, 9, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Gao, D.; Fukushima, H.; Inuzuka, H.; Liu, P.; Wan, L.; Sarkar, F.H.; Wei, W. Skp2: A novel potential therapeutic target for prostate cancer. Biochim. Biophys. Acta 2012, 1825, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, D.; Banerjee, D. Lactate Dehydrogenases as Metabolic Links between Tumor and Stroma in the Tumor Microenvironment. Cancers 2019, 11, 750. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Zhao, Y.; Ding, Y.; Liu, H.; Liu, Z.; Fodstad, O.; Riker, A.I.; Kamarajugadda, S.; Lu, J.; Owen, L.B.; et al. Warburg effect in chemosensitivity: Targeting lactate dehydrogenase-A re-sensitizes taxol-resistant cancer cells to taxol. Mol. Cancer 2010, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Duzdevich, D.; Warner, M.D.; Ticau, S.; Ivica, N.A.; Bell, S.P.; Greene, E.C. The dynamics of eukaryotic replication initiation: Origin specificity, licensing, and firing at the single-molecule level. Mol. Cell 2015, 58, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Evrin, C.; Fernández-Cid, A.; Riera, A.; Zech, J.; Clarke, P.; Herrera, M.C.; Tognetti, S.; Lurz, R.; Speck, C. The ORC/Cdc6/MCM2-7 complex facilitates MCM2-7 dimerization during prereplicative complex formation. Nucleic Acids Res. 2014, 42, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Kwok, H.F.; Zhang, S.-D.; McCrudden, C.M.; Yuen, H.-F.; Ting, K.-P.; Wen, Q.; Khoo, U.-S.; Chan, K.Y.-K. Prognostic significance of minichromosome maintenance proteins in breast cancer. Am. J. Cancer Res. 2014, 5, 52–71. [Google Scholar] [PubMed]
- Issac, M.S.M.; Yousef, E.; Tahir, M.R.; Gaboury, L.A. MCM2, MCM4, and MCM6 in Breast Cancer: Clinical Utility in Diagnosis and Prognosis. Neoplasia 2019, 21, 1015–1035. [Google Scholar] [CrossRef]
- Meng, M.V.; Grossfeld, G.D.; Williams, G.H.; Dilworth, S.; Stoeber, K.; Mulley, T.W.; Weinberg, V.; Carroll, P.R.; Tlsty, T.D. Minichromosome maintenance protein 2 expression in prostate: Characterization and association with outcome after therapy for cancer. Clin. Cancer Res. 2001, 7, 2712–2718. [Google Scholar]
- Toubaji, A.; Sutcliffe, S.; Chaux, A.; Lecksell, K.; Hicks, J.; De Marzo, A.M.; Platz, E.A.; Netto, G.J. Immunohistochemical expression of minichromosome maintenance complex protein 2 predicts biochemical recurrence in prostate cancer: A tissue microarray and digital imaging analysis-based study of 428 cases. Hum. Pathol. 2012, 43, 1852–1865. [Google Scholar] [CrossRef] [Green Version]
- Yousef, E.M.; Furrer, D.; Laperriere, D.L.; Tahir, M.R.; Mader, S.; Diorio, C.; Gaboury, L.A. MCM2: An alternative to Ki-67 for measuring breast cancer cell proliferation. Mod. Pathol. 2017, 30, 682–697. [Google Scholar] [CrossRef]
- Zhong, K.; Yang, F.; Han, Q.; Chen, J.; Wang, J. Skp2 expression has different clinicopathological and prognostic implications in lung adenocarcinoma and squamous cell carcinoma. Oncol. Lett. 2018, 16, 2873–2880. [Google Scholar] [CrossRef] [Green Version]
- Carrano, A.C.; Eytan, E.; Hershko, A.; Pagano, M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat. Cell Biol. 1999, 1, 193–199. [Google Scholar] [CrossRef]
- Bornstein, G.; Bloom, J.; Sitry-Shevah, D.; Nakayama, K.; Pagano, M.; Hershko, A. Role of the SCFSkp2 ubiquitin ligase in the degradation of p21Cip1 in S phase. J. Biol. Chem. 2003, 278, 25752–25757. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Cao, L.; Sun, Z.; Xu, J.; Tang, L.; Chen, W.; Luo, J.; Yang, F.; Wang, Y.; Guan, X. Skp2 is over-expressed in breast cancer and promotes breast cancer cell proliferation. Cell Cycl. 2016, 15, 1344–1351. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Du, L.; Ren, Y.; Liu, X.; Jiao, Q.; Cui, D.; Wen, M.; Wang, C.; Wei, G.; Wang, Y.; et al. SKP2 promotes breast cancer tumorigenesis and radiation tolerance through PDCD4 ubiquitination. J. Exp. Clin. Cancer Res. 2019, 38, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimečková, Š.; Kahounová, Z.; Fedr, R.; Remšík, J.; Slabáková, E.; Suchánková, T.; Procházková, J.; Bouchal, J.; Kharaishvili, G.; Král, M.; et al. High Skp2 expression is associated with a mesenchymal phenotype and increased tumorigenic potential of prostate cancer cells. Sci. Rep. 2019, 9, 5695. [Google Scholar] [CrossRef]
- Yang, Y.; Lu, Y.; Wang, L.; Mizokami, A.; Keller, E.T.; Zhang, J.; Fu, J. Skp2 is associated with paclitaxel resistance in prostate cancer cells. Oncol. Rep. 2016, 36, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, D.; He, J.; Li, C.F.; Lee, H.J.; Liu, J.; Lin, H.K.; Chan, C.H. Skp2 deficiency restricts the progression and stem cell features of castration-resistant prostate cancer by destabilizing Twist. Oncogene 2017, 36, 4299–4310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-On, O.; Shapira, M.; Hershko, D.D. Differential effects of doxorubicin treatment on cell cycle arrest and Skp2 expression in breast cancer cells. Anticancer Drugs 2007, 18, 1113–1121. [Google Scholar] [CrossRef]
- Acree, W.E.; Powell, J.R.; Tucker, S.A.; Ribeiro da Silva, M.D.M.C.; Matos, M.A.R.; Gonçalves, J.M.; Santos, L.M.N.B.F.; Morais, V.M.F.; Pilcher, G. Thermochemical and Theoretical Study of Some Quinoxaline 1,4-Dioxides and of Pyrazine 1,4-Dioxide. J. Organ. Chem. 1997, 62, 3722–3726. [Google Scholar] [CrossRef]
- Ribeiro da Silva, M.D.M.C.; Gomes, J.R.B.; Gonçalves, J.M.; Sousa, E.A.; Pandey, S.; Acree, W.E. Thermodynamic Properties of Quinoxaline-1,4-Dioxide Derivatives: A Combined Experimental and Computational Study. J. Organ. Chem. 2004, 69, 2785–2792. [Google Scholar] [CrossRef]
- Ferraz, R.; Branco, L.C.; Marrucho, I.M.; Araújo, J.M.M.; Rebelo, L.P.N.; da Ponte, M.N.; Prudêncio, C.; Noronha, J.P.; Petrovski, Ž. Development of novel ionic liquids based on ampicillin. MedChemComm 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.L.; Wen, X.M.; Lai, Q.H.; Li, J.; Wang, X.W. Lenalidomide improvement of cisplatin antitumor efficacy on triple-negative breast cancer cells in vitro. Oncol. Lett. 2018, 15, 6469–6474. [Google Scholar] [CrossRef] [Green Version]
- Pan, T.; Mao, T.; Yang, H.; Wang, H.; Wang, Y. Silencing of TGIF sensitizes MDA-MB-231 human breast cancer cells to cisplatin-induced apoptosis. Exp. Ther. Med. 2018, 15, 2978–2984. [Google Scholar] [CrossRef] [Green Version]
- Dunne, M.; Dou, Y.N.; Drake, D.M.; Spence, T.; Gontijo, S.M.; Wells, P.G.; Allen, C. Hyperthermia-mediated drug delivery induces biological effects at the tumor and molecular levels that improve cisplatin efficacy in triple negative breast cancer. J. Control. Release 2018, 282, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; Turkekul, K.; Serttas, R.; Erdogan, Z. The natural flavonoid apigenin sensitizes human CD44+ prostate cancer stem cells to cisplatin therapy. Biomed. Pharmacother. 2017, 88, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yue, C.; Li, J.; Wu, J.; Wang, S.; Sun, D.; Guo, Y.; Lin, Z.; Zhang, D.; Wang, R. Enhancement of cisplatin cytotoxicity by Retigeric acid B involves blocking DNA repair and activating DR5 in prostate cancer cells. Oncol. Lett. 2018, 15, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Li, Y.; Zeng, J.; Wang, B.; Ji, K.; Tang, Y.; Sun, Q. Knockdown of HIF-1α by siRNA-expressing plasmid delivered by attenuated salmonella enhances the antitumor effects of cisplatin on prostate cancer. Sci. Rep. 2017, 7, 7546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prayong, P.; Barusrux, S.; Weerapreeyakul, N. Cytotoxic activity screening of some indigenous Thai plants. Fitoterapia 2008, 79, 598–601. [Google Scholar] [CrossRef]
- Vieira-Silva, T.S.; Monteiro-Reis, S.; Barros-Silva, D.; Ramalho-Carvalho, J.; Graca, I.; Carneiro, I.; Martins, A.T.; Oliveira, J.; Antunes, L.; Hurtado-Bages, S.; et al. Histone variant MacroH2A1 is downregulated in prostate cancer and influences malignant cell phenotype. Cancer Cell Int. 2019, 19, 112. [Google Scholar] [CrossRef] [Green Version]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Cell Lines | IC50 Value ± SD (µM) | |
|---|---|---|---|
| [C16Pyr][Amp] | Cisplatin | ||
| Breast | MCF-10A | 2.1 ± 0.27 | 4.9 ± 0.42 |
| HTB22 | 0.6 ± 0.08 | 10.9 ± 0.62 | |
| HTB133 | 2.0 ± 0.55 | 8.2 ± 0.69 | |
| HCC1937 | 0.8 ± 0.01 | 3.5 ± 0.25 | |
| MDA-MB-231 | 0.2 ± 0.01 | 1.1 ± 0.10 | |
| Prostate | RWPE | 0.9 ± 0.13 | 4.9 ± 0.06 |
| 22Rv1 | 1.4 ± 0.16 | 5.5 ± 0.18 | |
| LNCaP | 0.1 ± 0.02 | 7.4 ± 0.33 | |
| Du145 | 0.9 ± 0.08 | 0.6 ± 0.01 | |
| PC-3 | 0.3 ± 0.00 | 5.1 ± 0.67 | |
| Tumor Model | Cell Lines | Selectivity Index | |
|---|---|---|---|
| [C16Pyr][Amp] | Cisplatin | ||
| Breast cancer | HTB22 | 3.73 | 0.45 |
| HTB133 | 1.05 | 0.60 | |
| HCC1937 | 2.66 | 1.41 | |
| MDA-MB-231 | 11.00 | 4.37 | |
| Prostate cancer | 22Rv1 | 0.66 | 0.89 |
| LNCaP | 7.16 | 0.66 | |
| Du145 | 1.01 | 8.20 | |
| PC-3 | 2.63 | 0.96 | |
| Genes | HTB133 | MDA-MB-231 | 22Rv1 | Du145 | ||||
|---|---|---|---|---|---|---|---|---|
| Fold Change | p-Value | Fold Change | p-Value | Fold Change | p-Value | Fold Change | p-Value | |
| CPT2 | −1.51 | 0.160 | −1.22 | 0.360 | −10.65 | 0.006 | −1.73 | 0.004 |
| LDHA | −5.21 | <0.001 | −3.68 | 0.0001 | −1.04 | 0.840 | −1.15 | 0.203 |
| MCM2 | −2.78 | <0.0001 | −1.99 | 0.003 | −1.55 | 0.030 | −1.18 | 0.163 |
| SKP2 | −2.90 | <0.0001 | −2.86 | <0.0001 | −2.36 | 0.030 | −1.96 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, F.Q.; Marques-Magalhães, Â.; Miranda-Gonçalves, V.; Ferraz, R.; Vieira, M.; Prudêncio, C.; Jerónimo, C.; Silva, R.A. The Impact of [C16Pyr][Amp] on the Aggressiveness in Breast and Prostate Cancer Cell Lines. Int. J. Mol. Sci. 2020, 21, 9584. https://doi.org/10.3390/ijms21249584
Vieira FQ, Marques-Magalhães Â, Miranda-Gonçalves V, Ferraz R, Vieira M, Prudêncio C, Jerónimo C, Silva RA. The Impact of [C16Pyr][Amp] on the Aggressiveness in Breast and Prostate Cancer Cell Lines. International Journal of Molecular Sciences. 2020; 21(24):9584. https://doi.org/10.3390/ijms21249584
Chicago/Turabian StyleVieira, Filipa Quintela, Ângela Marques-Magalhães, Vera Miranda-Gonçalves, Ricardo Ferraz, Mónica Vieira, Cristina Prudêncio, Carmen Jerónimo, and Regina Augusta Silva. 2020. "The Impact of [C16Pyr][Amp] on the Aggressiveness in Breast and Prostate Cancer Cell Lines" International Journal of Molecular Sciences 21, no. 24: 9584. https://doi.org/10.3390/ijms21249584