Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Food-Choice Treatments
2.3. Data Collection and Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimers
References
- Ng, W.-K.; Liew, F.-L.; Ang, L.-P.; Wong, K.-W. Potential of mealworm (Tenebrio molitor) as an alternative protein source in practical diets for African catfish, Clarias gariepinus. Aquac. Res. 2001, 32, 272–280. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañon, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Gasco, L.; Henry, M.; Piccolo, G.; Marono, S.; Gai, F.; Renna, M.; Lussiana, C.; Antonopoulou, E.; Mola, P.; Chatzifotis, S. Tenebrio molitor meal in diets for European sea bass (Dicentrarchus labrax L.) juveniles: Growth performance, whole body composition and in vivo apparent digestibility. Anim. Feed Sci. Technol. 2016, 220, 34–45. [Google Scholar] [CrossRef]
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal alternative protein sources for aquaculture. In Feeds for the Aquaculture Sector; Gasco, L., Gai, F., Maricchiolo, G., Genovese, L., Ragonese, S., Bottari, T., Caruso, G., Eds.; Springer Briefs in Molecular Science; Springer: Cham, Switzerland, 2018; pp. 1–28. [Google Scholar]
- Lock, E.J.; Biancarosa, I.; Gasco, L. Insects as raw materials in compound feed for aquaculture. In Edible Insects in Sustainable Food Systems; Halloran, A., Flore, R., Vantomme, P., Roos, N., Eds.; Springer: Cham, Switzerland, 2018; pp. 263–276. [Google Scholar]
- Ferrer, L.P.; Kallas, Z.; de Amores Gea, D. The use of insect meal as a sustainable feeding alternative in aquaculture: Current situation, Spanish consumers’ perceptions and willingness to pay. J. Clean. Prod. 2019, 229, 10–21. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J.; Avila Gonzalez, E.; Rocha Hernandez, A.; Pino, J.M. Use of Tenebrio molitor (Coleoptera, Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 2002, 95, 214–220. [Google Scholar] [CrossRef] [PubMed]
- van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects, Future Prospects for Food and Feed Security; FAO, ONU: Rome, Italy, 2013. [Google Scholar]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.; Bergero, D.; Katz, H.; Dabbou, S.; et al. Nutritional value of two insect larval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed Sci. Technol. 2015, 209, 211–218. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Barroso, F.G.; de Haro, C. Brief summary of insect usage as an industrial animal feed/feed ingredient. In Insects as Sustainable Food Ingredients–Production, Processing and Food Applications; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 273–309. [Google Scholar]
- Benzertiha, A.; Kierończyk, B.; Rawski, M.; Józefiak, A.; Kozłowski, K.; Jankowski, J.; Józefiak, D. Tenebrio molitor and Zophobas morio full-fat meals in broiler chicken diets: Effects on nutrients digestibility, digestive enzyme activities, and cecal microbiome. Animals 2019, 9, 1128. [Google Scholar] [CrossRef]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef]
- Dossey, A.T.; Tatum, J.T.; McGill, W.L. Modern insect-based food industry: Current status, insect processing technology, and recommendations moving forward. In Insects as Sustainable Food Ingredients–Production, Processing and Food Applications; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 113–152. [Google Scholar]
- van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Ins. Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, H.G.; Lee, K.Y.; Yoon, H.J.; Kim, N.J. Effects of brewer’s spent grain (BSG) on larval growth of mealworms, Tenebrio molitor (Coleoptera: Tenebrionidae). Int. J. Ind. Entomol. 2016, 32, 41–48. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Chen, G.; Qiao, L.; Li, J.; Liu, B.; Liu, Z.; Li, M.; Liu, X. Growth performance and nutritional profile of mealworms reared on corn stover, soybean meal, and distillers’ grains. Eur. Food Res. Technol. 2019, 245, 2631–2640. [Google Scholar] [CrossRef]
- Li, T.-H.; Che, P.-F.; Zhang, C.-R.; Zhang, B.; Ali, A.; Zang, L.-S. Recycling of spent mushroom substrate: Utilization as feed material for the larvae of the yellow mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). PLoS ONE 2020, 15, e0237259. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.A.; Rojas, M.G.; Dossey, A.T.; Berhow, M. Self-selection of food ingredients and agricultural by-products by the house cricket, Acheta domesticus (Orthoptera: Gryllidae): A Holistic approach to develop optimized diets. PLoS ONE 2020, 15, e0227400. [Google Scholar] [CrossRef] [PubMed]
- Waldbauer, G.P.; Friedman, S. Self-selection of optimal diets by insects. Annu Rev. Entomol. 1991, 36, 43–63. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Kelstrup, H.C.; Rojas, M.G.; Emery, V. Body mass increase induced by eight years of artificial selection in the yellow mealworm (Coleoptera: Tenebrionidae) and life history trade-offs. J. Ins. Sci. 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Kay, S.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Morphometric analysis of instar variation in Tenebrio molitor (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 2015, 108, 146–158. [Google Scholar] [CrossRef]
- Waldbauer, G.P. The consumption and utilization of food by insects. Adv. Ins. Physiol. 1968, 5, 229–288. [Google Scholar]
- Morales-Ramos, J.A.; Rojas, M.G.; Coudron, T.A. Artificial diet development for entomophagous arthropods. In Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; Morales-Ramos, J.A., Rojas, M.G., Shapiro-Ilan, D.I., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 203–240. [Google Scholar]
- National Research Council (NRC). United States-Canadian Tables of Feed Composition: Nutritional Data for United States and Canadian Feeds, 3rd ed.; National Academy Press: Washington, DC, USA, 1982; Available online: http://nap.edu/1713 (accessed on 29 August 2019).
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994; Available online: http://nap.edu/2114 (accessed on 30 August 2019).
- Wu, Y.V.; Stringfellow, A.C. Corn distillers’ dried grains with solubles and corn distillers’ dried grains: Dry fraction and composition. J. Food Sci. 1982, 47, 1155–1157. [Google Scholar] [CrossRef]
- Batajoo, K.K.; Shaver, K.D. In Situ dry matter, crude protein, and starch degradabilities of selected grains and by-products. Anim. Feed Sci Technol. 1998, 71, 165–176. [Google Scholar] [CrossRef]
- Spiehs, M.J.; Whitney, M.H.; Shurson, G.C. Nutrient database for distiller’s dried grains with solubles produced from new ethanol plants in Minnesota and South Dakota. J. Anim. Sci. 2002, 80, 2639–2645. [Google Scholar] [PubMed]
- Huzá, S.; Várhegyi, J.; Lehel, L.; Rózsa, L.; Kádár, M. Mineral content of grains, seeds and industrial by products. Állattenyésztés Takarm. 2003, 52, 277–283. [Google Scholar]
- Kim, Y.; Moiser, N.S.; Hendrickson, R.; Ezeji, T.; Blaschek, H.; Dien, B.; Cotta, M.; Dale, B.; Ladisch, M.R. Composition of corn dry-grain ethanol by-products: DDGS, wet cake, and thin stillage. Bioresour. Technol. 2008, 99, 5165–5176. [Google Scholar] [CrossRef] [PubMed]
- Kahlon, T.S. Rice bran: Production, composition, functionality and food applications, physiological benefits. In Fiber Ingredients: Food Applications and Health Benefits; Cho, S.S., Samuel, P., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2009; pp. 305–321. [Google Scholar]
- Newkirk, R. Canola Meal Feed Industry Guide, 4th ed.; Canadian International Grains Institute, Canola Council of Canada: Winnipeg, MB, Canada, 2009. [Google Scholar]
- U. S. Grains Council. A guide to Distiller’s Dried Grains with Solubles (DDGS), 3rd ed.; U. S. Grains Council: Washington, DC, USA, 2012. [Google Scholar]
- US Department of Agriculture (USDA). USDA National Nutrient Database for Standard Reference; US Department of Agriculture: Beltsville, MD, USA, 2015. Available online: http://www.ars.usda.gov/ba/bhnrc/ndl (accessed on 13 October 2015).
- Amorim, M.; Pereira, J.O.; Gomes, D.; Pereira, C.D.; Pinheiro, H.; Pintado, M. Nutritional ingredients from spent brewer’s yeast obtained by hydrolysis and selective membrane filtration integrated in a pilot process. J. Food Eng. 2016, 185, 42–47. [Google Scholar] [CrossRef]
- SAS Institute. JMP Statistical Discovery from SAS, Version 14: Fitting Linear Models; SAS Institute: Cary, NC, USA, 2018. [Google Scholar]
- Myers, R.H. Classical and Modern Regression with Applications; Duxbury Press: Boston, MA, USA, 1986. [Google Scholar]
- Freund, R.; Littell, R.; Creighton, L. Regression Using JMP®; SAS Institute Inc.: Cary, NC, USA, 2003. [Google Scholar]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Self-selection of two diet components by Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and its impact on fitness. Environ. Entomol. 2011, 40, 1285–1294. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro Ilan, D.I.; Tedders, W.L. Use of nutrient self-selection as a diet refining tool in Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, P. Geometric analysis of nutrient balancing in the mealworm beetle, Tenebrio molitor L. (Coleoptera: Tenebrionidae). J. Ins. Physiol. 2014, 71, 37–45. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, P. Nutrient-specific food selection buffers the effect of nutritional imbalance in the mealworm beetle, Tenebrio molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2015, 112, 251–258. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, P. Balanced intake of protein and carbohydrate maximizes lifetime reproductive success in the mealworm beetle, Tenebrio molitor (Coleoptera: Tenebrionidae). J. Ins. Physiol. 2016, 91–92, 93–99. [Google Scholar] [CrossRef]
- Cortes Ortiz, J.A.; Ruiz, A.T.; Morales-Ramos, J.A.; Thomas, M.; Rojas, M.G.; Tomberlin, J.K.; Yi, L.; Han, R.; Giroud, L.; Jullien, R.L. Insect mass production technologies. In Insects as Sustainable Food Ingredients–Production, Processing and Food Applications; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 154–201. [Google Scholar]
- Li, L.; Stasiak, M.; Li, L.; Xie, B.; Fu, Y.; Gidzinski, D.; Dixon, M.; Liu, H. Rearing Tenebrio molitor in BLSS: Dietary fiber affects larval growth, development, and respiration characteristics. Acta Astronaut. 2015, 118, 130–136. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
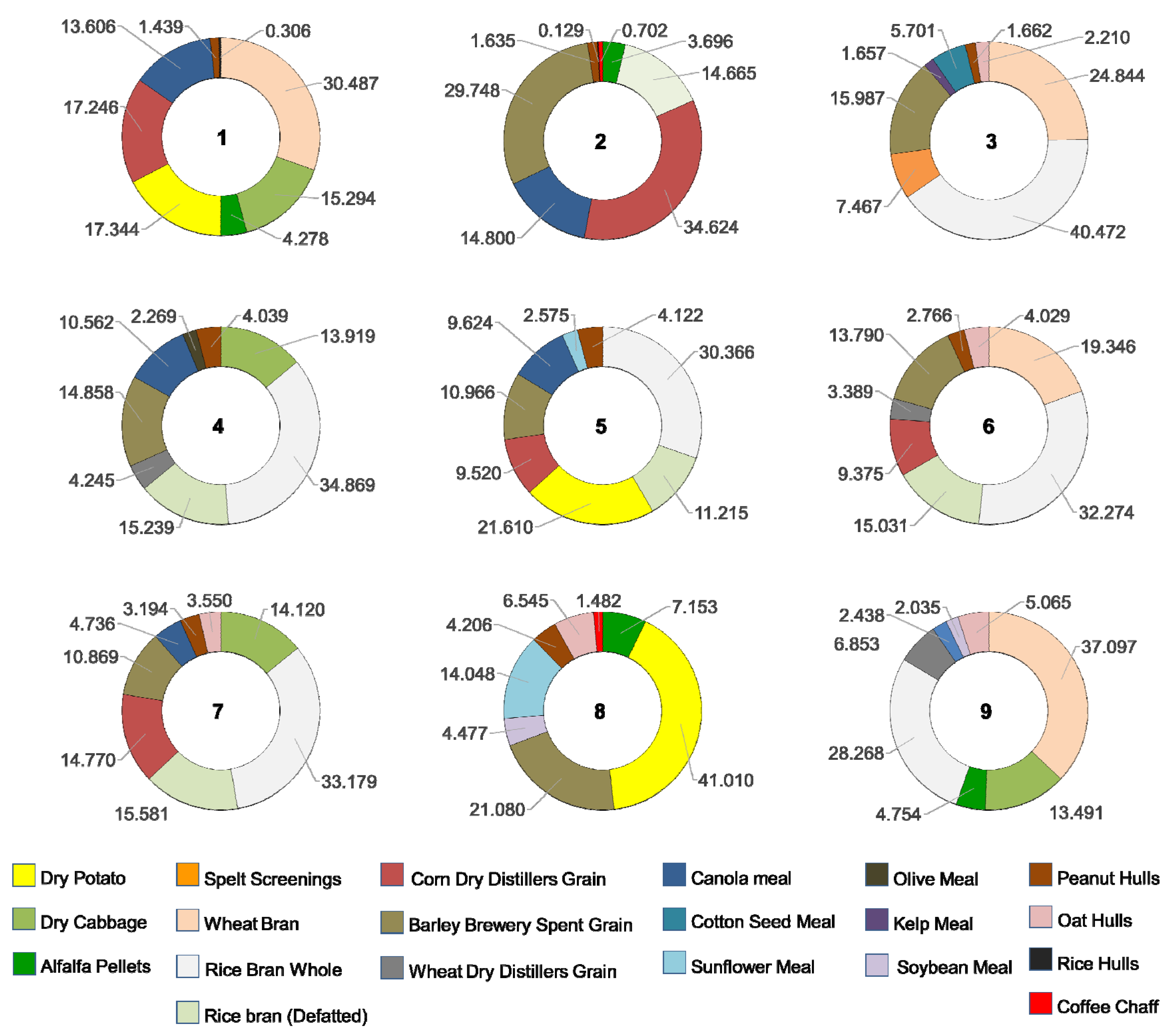
| Ingredient | Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| White Cabbage | X | X | X | X | |||||
| Potato | X | X | X | ||||||
| Alfalfa pellets | X | X | X | X | |||||
| Wheat Bran | X | X | X | X | |||||
| Rice bran whole | X | X | X | X | X | X | |||
| Rice bran defatted | X | X | X | X | X | ||||
| Spelt Screenings | X | ||||||||
| Corn dry distiller’s grain | X | X | X | X | X | ||||
| Wheat dry distiller’s grain | X | X | X | ||||||
| Barley brewery spent grain | X | X | X | X | X | X | X | ||
| Canola meal | X | X | X | X | X | X | |||
| Soy Meal | X | X | |||||||
| Sunflower meal | X | X | |||||||
| Olive meal | X | ||||||||
| Cotton seed meal | X | ||||||||
| Kelp meal | X | ||||||||
| Oat hulls | X | X | X | X | X | ||||
| Peanut hulls | X | X | X | X | X | X | X | X | |
| Rice hulls | X | X | |||||||
| Coffee chaff | X | X | |||||||
| Ingredient | Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| White Cabbage | 4.49 ± 0.91 | 3.85 ± 0.35 | 4.16 ± 0.61 | 4.14 ± 0.45 | |||||
| Potato | 5.09 ± 0.63 | 6.09 ± 0.55 | 10.33 ± 0.93 | ||||||
| Alfalfa pellets | 1.26 ± 0.22 | 1.07 ± 0.24 | 1.8 ± 0.33 | 1.46 ± 0.24 | |||||
| Wheat Bran | 8.95 ± 1.1 | 6.99 ± 0.98 | 5.81 ± 0.79 | 11.37 ± 0.81 | |||||
| Rice bran whole | 11.38 ± 0.98 | 9.64 ± 0.73 | 8.55 ± 0.88 | 9.69 ± 0.51 | 9.78 ± 2.09 | 8.67 ± 1.7 | |||
| Rice bran defatted | 4.23 ± 1.19 | 4.21 ± 0.72 | 3.16 ± 1.03 | 4.51 ± 0.89 | 4.59 ± 1.84 | ||||
| Spelt Screenings | 2.1 ± 0.35 | ||||||||
| Corn DDGS | 5.06 ± 1.09 | 9.99 ± 1.23 | 2.68 ± 0.65 | 2.82 ± 0.64 | 4.35 ± 0.82 | ||||
| Wheat DDGS | 1.17 ± 0.26 | 1.02 ± 0.16 | 2.1 ± 0.75 | ||||||
| Barley brewery spent grain | 8.58 ± 0.59 | 4.5 ± 0.59 | 4.11 ± 0.52 | 3.09 ± 0.89 | 4.14 ± 0.4 | 3.2 ± 0.43 | 5.31 ± 0.43 | ||
| Canola meal | 3.99 ± 0.9 | 4.27 ± 0.85 | 2.92 ± 0.31 | 2.71 ± 0.3 | 1.4 ± 0.46 | 0.75 ± 0.35 | |||
| Soybean Meal | 1.13 ± 0.14 | 0.62 ± 0.07 | |||||||
| Sunflower meal | 0.73 ± 0.16 | 3.54 ± 1.03 | |||||||
| Olive meal | 0.63 ± 0.11 | ||||||||
| Cotton seed meal | 1.6 ± 0.35 | ||||||||
| Kelp meal | 0.47 ± 0.05 | ||||||||
| Oat hulls | 0.62 ± 0.29 | 1.21 ± 0.22 | 1.05 ± 0.19 | 1.65 ± 0.36 | 1.55 ± 0.39 | ||||
| Peanut hulls | 0.42 ± 0.19 | 0.47 ± 0.12 | 0.47 ± 0.18 | 1.12 ± 0.21 | 1.16 ± 0.31 | 0.83 ± 0.18 | 0.94 ± 0.38 | 1.06 ± 0.54 | |
| Rice hulls | 0.09 ± 0.03 | 0.04 ± 0.02 | |||||||
| Coffee shaft | 0.2 ± 0.09 | 0.37 ± 0.14 | |||||||
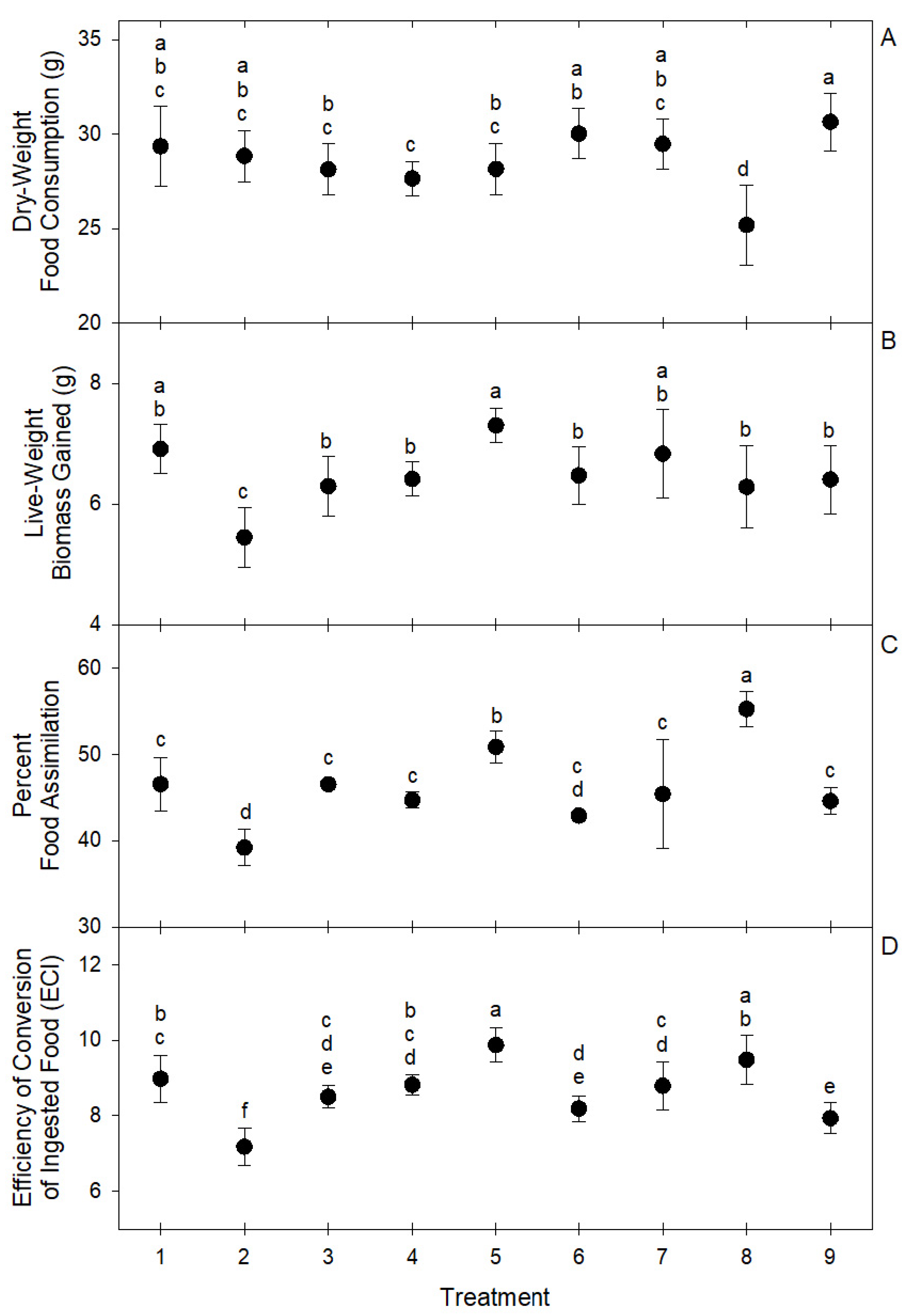
| Total Consumption | 29.36 ± 2.13 | 28.84 ± 1.36 | 28.13 ± 1.36 | 27.06 ± 0.9 | 28.16 ± 1.35 | 30.03 ± 1.32 | 29.48 ± 1.35 | 25.18 ± 2.12 | 30.66 ± 1.53 |
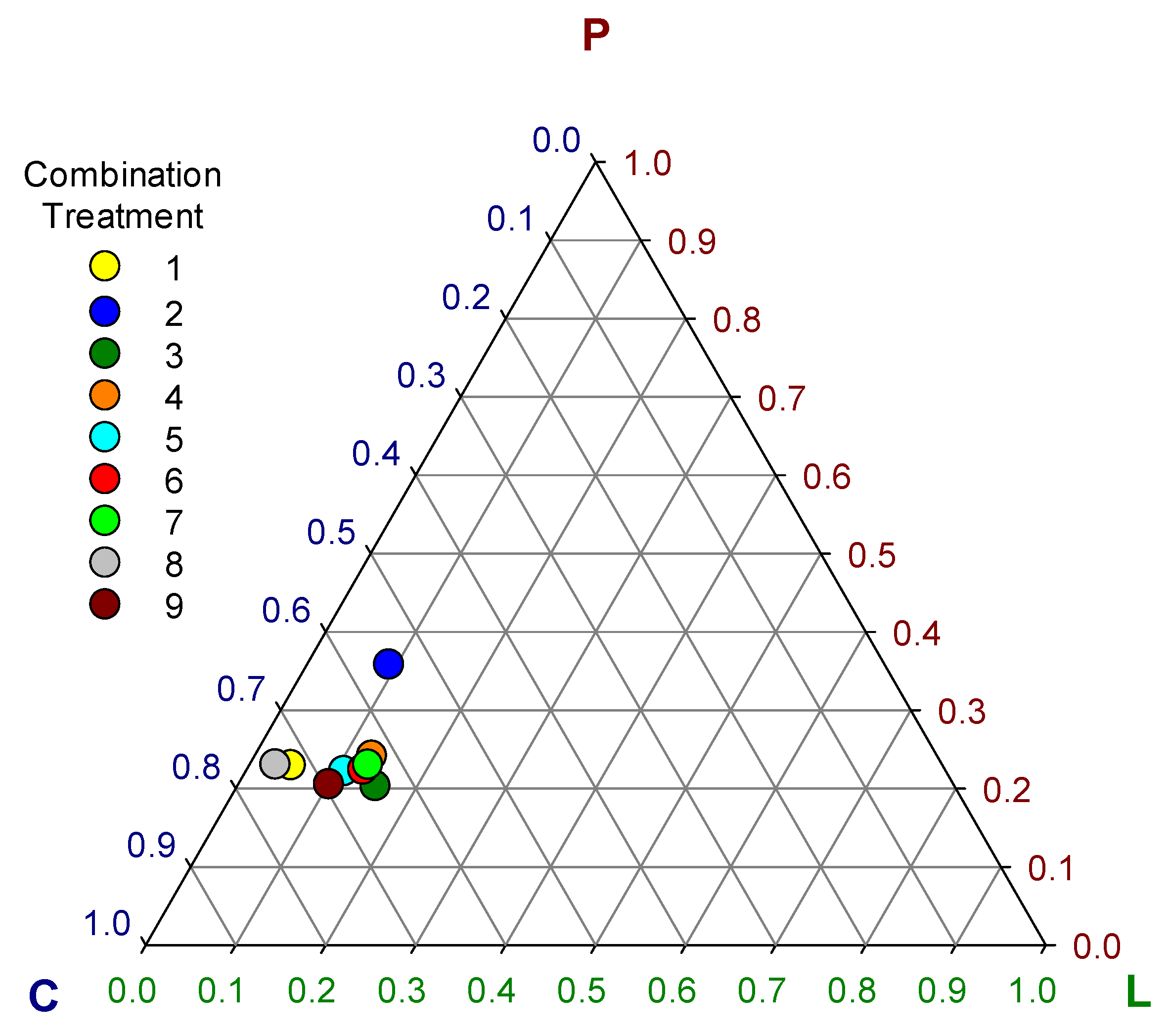
| Treatment | Lipid | Protein | Carbohydrate |
|---|---|---|---|
| 1 | 0.047 ± 0.003 e | 0.23 ± 0.007 c | 0.723 ± 0.008 b |
| 2 | 0.092 ± 0.003 d | 0.359 ± 0.006 a | 0.549 ± 0.008 g |
| 3 | 0.157 ± 0.006 a | 0.206 ± 0.005 d | 0.637 ± 0.003 e |
| 4 | 0.132 ± 0.006 b | 0.244 ± 0.005 b | 0.624 ± 0.006 f |
| 5 | 0.112 ± 0.005 c | 0.22 ± 0.012 c | 0.668 ± 0.01 d |
| 6 | 0.134 ± 0.004 b | 0.23 ± 0.004 c | 0.636 ± 0.005 e |
| 7 | 0.132 ± 0.017 b | 0.23 ± 0.008 c | 0.638 ± 0.014 e |
| 8 | 0.029 ± 0.002 f | 0.23 ± 0.008 c | 0.741 ± 0.009 a |
| 9 | 0.101 ± 0.011 d | 0.205 ± 0.009 d | 0.694 ± 0.009 c |
| Nutrient | Mean ± SD | Minimum | Maximum |
|---|---|---|---|
| Lipid | 8.13 ± 3.13 | 2.27 | 13.03 |
| Protein | 18.75 ± 2.78 | 15.58 | 26.45 |
| Carbohydrate | 51.94 ± 6.54 | 38.71 | 64.84 |
| Fiber (ND) | 28.06 ± 3.48 | 21.72 | 36.41 |
| Ca | 0.445 ± 0.246 | 0.11 | 0.906 |
| Fe | 0.014 ± 0.002 | 0.009 | 0.018 |
| Mg | 0.439 ± 0.113 | 0.157 | 0.568 |
| Zn | 0.004 ± 0.0007 | 0.002 | 0.005 |
| Mn | 0.007 ± 0.002 | 0.002 | 0.01 |
| Parameter | Estimate | Sum of Squares | F Ratio | p > F |
|---|---|---|---|---|
| Carbohydrate | 5.4 × 10−4 ± 4.3 × 10−5 | 1121.48 | 158.76 | <0.0001 |
| Fiber | −7.4 × 10−4 ± 8.1 × 10−5 | 584.35 | 82.72 | <0.0001 |
| Parameter | Estimate | Sum of Squares | F Ratio | p > F |
|---|---|---|---|---|
| Carbohydrate | 1.2 × 10−4 ± 1.0 × 10−5 | 27.22 | 120.17 | <0.0001 |
| Fiber | −1.6 × 10−4 ± 1.5 × 10−5 | 24.88 | 109.83 | <0.0001 |
| Mg | 0.017 ± 0.003 | 6.14 | 27.1 | <0.0001 |
| Mn | −0.667 ± 0.14 | 4.14 | 22.71 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Ramos, J.A.; Rojas, M.G.; Kelstrup, H.C.; Emery, V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects 2020, 11, 827. https://doi.org/10.3390/insects11120827
Morales-Ramos JA, Rojas MG, Kelstrup HC, Emery V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects. 2020; 11(12):827. https://doi.org/10.3390/insects11120827
Chicago/Turabian StyleMorales-Ramos, Juan A., M. Guadalupe Rojas, Hans C. Kelstrup, and Virginia Emery. 2020. "Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake" Insects 11, no. 12: 827. https://doi.org/10.3390/insects11120827
APA StyleMorales-Ramos, J. A., Rojas, M. G., Kelstrup, H. C., & Emery, V. (2020). Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects, 11(12), 827. https://doi.org/10.3390/insects11120827