
Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production


Abstract
1. Introduction
2. Results and Discussion
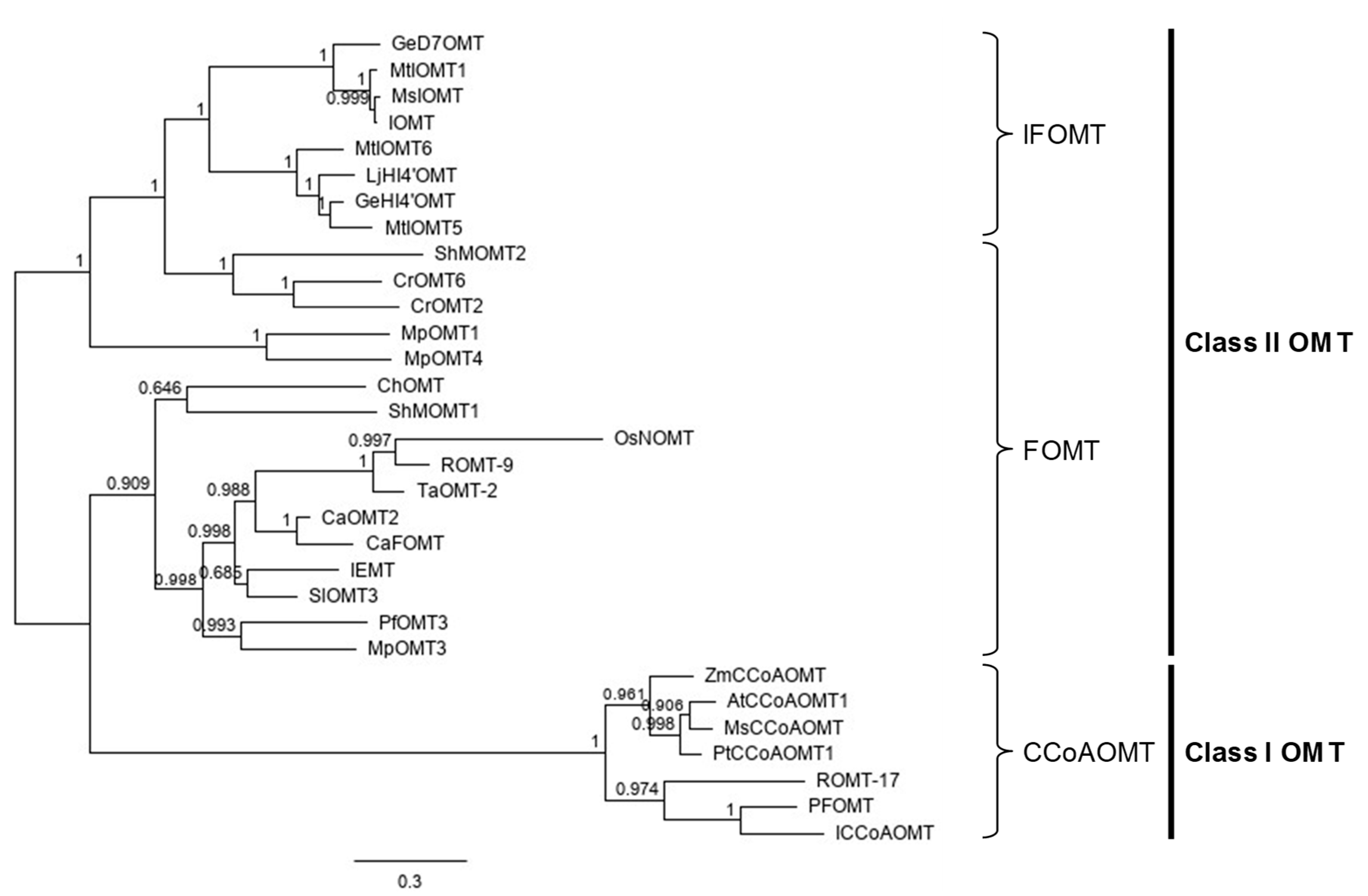
2.1. Cloning and Molecular Characterization of PfOMT3 from Perilla Leaves
2.2. PfOMT3 is a Flavonoid 7-O-Methyltransferase
2.3. Substrate Preference of PfOMT3
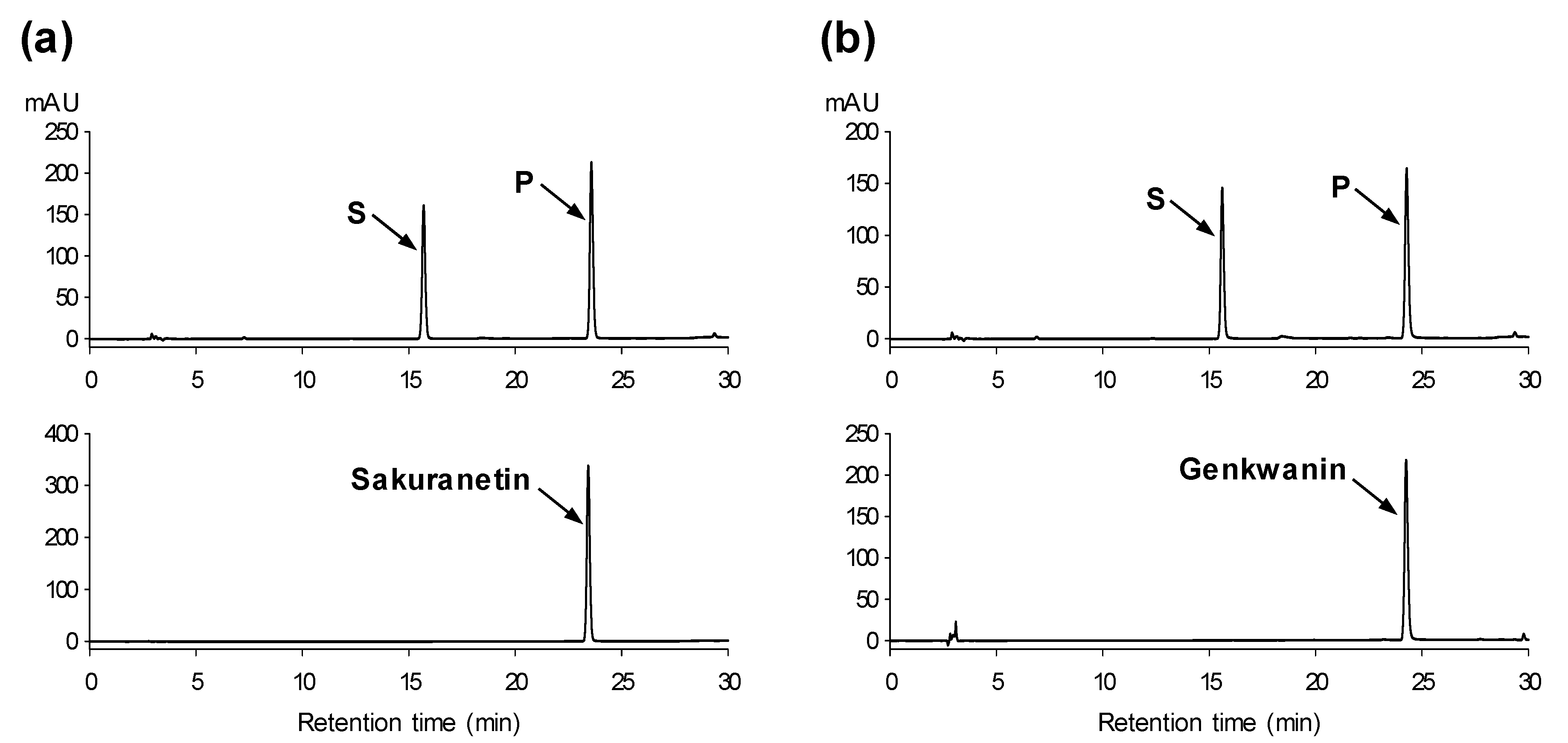
2.4. Application of PfOMT3 in the Biotechnological Production of 7-Methoxyflavonoids
3. Materials and Methods
3.1. Materials
3.2. Cloning and Phylogenetic Analysis of PfOMT3
3.3. Heterologous Expression and Purification of Recombinant PfOMT3
3.4. OMT Activity Assay and Kinetic Analysis
3.5. Biotransformation of Flavonoids using E. coli Bearing PfOMT3
3.6. Identification of Methylated Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 18–223. [Google Scholar] [CrossRef]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Kim, B.G.; Sung, S.H.; Chong, Y.; Lim, Y.; Ahn, J.H. Plant flavonoid O-methyltransferases: Substrate specificity and application. J. Plant Biol. 2010, 53, 321–329. [Google Scholar] [CrossRef]
- Ibrahim, R.; De Luka, V.; Khouri, H.; Latchinian, L.; Brisson, L.; Charest, P.M. Enzymology and compartmentation of polymethylated flavonol glucosides in Chrysosplenium americanum. Phytochemistry 1987, 26, 1237–1245. [Google Scholar] [CrossRef]
- Ibrahim, R.K.; Bruneau, A.; Bantignies, B. Plant O-methyltransferases: Molecular analysis, common signature and classification. Plant Mol. Biol. 1998, 36, 1–10. [Google Scholar] [CrossRef]
- Lam, K.C.; Ibrahim, R.K.; Behdad, B.; Dayanandan, S. Structure, function, and evolution of plant O-methyltransferases. Genome 2007, 50, 1001–1013. [Google Scholar] [CrossRef]
- Walle, T. Methylation of dietary flavones greatly improves their hepatic metabolic stability and intestinal absorption. Mol. Pharm. 2007, 4, 826–832. [Google Scholar] [CrossRef]
- Wen, X.; Walle, T. Methylated flavonoids have greatly improved intestinal absorption and metabolic stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef]
- Cao, H.; Jing, X.; Wu, D.; Shi, Y. Methylation of genistein and kaempferol improves their affinities for proteins. Int. J. Food Sci. Nutr. 2013, 64, 437–443. [Google Scholar] [CrossRef]
- Zhang, L.; Kong, Y.; Wu, D.; Zhang, H.; Wu, J.; Chen, J.; Ding, J.; Hu, L.; Jiang, H.; Shen, X. Three flavonoids targeting the β-hydroxyacyl-acyl carrier protein dehydratase from Helicobacter pylori: Crystal structure characterization with enzymatic inhibition assay. Protein Sci. 2008, 17, 1971–1978. [Google Scholar] [CrossRef]
- Kodama, O.; Miyakawa, J.; Akatsuka, T.; Kiyosawa, S. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 1992, 31, 3807–3809. [Google Scholar] [CrossRef]
- Park, H.L.; Yoo, Y.; Hahn, T.R.; Bhoo, S.H.; Lee, S.W.; Cho, M.H. Antimicrobial activity of UV-induced phenylamides from rice leaves. Molecules 2014, 19, 18139–18151. [Google Scholar] [CrossRef] [PubMed]
- Azerad, R. Chemical biotechnology: Better enzymes for green chemistry. Curr. Opin. Biotechnol. 2001, 12, 533–534. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalytic synthesis of intermediates for the synthesis of chiral drug substances. Curr. Opin. Biotechnol. 2001, 12, 587–604. [Google Scholar] [CrossRef]
- Fowler, Z.L.; Koffas, M.A.G. Biosynthesis and biotechnological production of flavanones: Current state and perspectives. Appl. Microbiol. Biotechnol. 2009, 83, 799–808. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, B.G.; Lee, Y.; Ryu, J.Y.; Lim, Y.; Hur, H.G.; Ahn, J.H. Regiospecific methylation of naringenin to ponciretin by soybean O-methyltransferase expressed in Escherichia coli. J. Biotechnol. 2005, 119, 155–162. [Google Scholar] [CrossRef]
- Kim, B.G.; Lee, Y.; Hur, H.G.; Lim, Y.; Ahn, J.H. Flavonoid 3′-O-methyltransferase from rice: cDNA cloning, characterization and functional expression. Phytochemistry 2006, 67, 387–394. [Google Scholar] [CrossRef]
- Cho, M.H.; Park, H.L.; Park, J.H.; Lee, S.W.; Bhoo, S.H.; Hahn, T.R. Characterization of a regiospecific flavonoid 3′/5′-O-methyltransferase from tomato and its application in flavonoid biotransformation. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 749–755. [Google Scholar] [CrossRef]
- Shimizu, T.; Lin, F.; Hasegawa, M.; Okada, K.; Nojiri, H.; Yamane, H. Purification and identification of naringenin 7-O-methyltransferase, a key enzyme in biosynthesis of flavonoid phytoalexin sakuranetin in rice. J. Biol. Chem. 2012, 287, 19315–19325. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Park, H.L.; Lee, S.W.; Bhoo, S.H.; Cho, M. Biotechnological production of dimethoxyflavonoids using a fusion flavonoid O-methyltransferase possessing both 3′- and 7-O-methyltransferase activities. J. Nat. Prod. 2017, 80, 1467–1474. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.K.; Yoo, J.H.; Yu, C.Y.; Chung, I.M. GC-MS analysis of volatile compounds of Perilla frutescens Britton var. Japonica accessions: Morphological and seasonal variability. Asian Pac. J. Trop. Med. 2017, 10, 643–651. [Google Scholar]
- Lee, Y.H.; Kim, B.; Kim, S.; Kim, M.S.; Kim, H.; Hwang, S.R.; Kim, K.; Lee, J.H. Characterization of metabolite profiles from the leaves of green perilla (Perilla frutescens) by ultra high performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight mass spectrometry and screening for their antioxidant properties. J. Food Drug Anal. 2017, 25, 776–788. [Google Scholar]
- Yu, H.; Qui, J.F.; Ma, L.J.; Hu, Y.J.; Li, P.; Wan, J.B. Phytochemical and phytopharmacological review of Perilla frutescens L. (Labiatae), a traditional edible-medicinal herb in China. Food Chem. Toxicol. 2017, 108, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Ohk, H.C.; Chae, Y.A. Analysis of volatile oil components and identification of chemotypes in Jaso (Perilla frutescens) collected in Korea. Korean J. Med. Crop Sci. 2004, 12, 97–101. [Google Scholar]
- Fukushima, A.; Nakamura, M.; Suzuki, H.; Saito, K.; Yamazaki, M. High-throughput sequencing and de novo assembly of red and green forms of the Perilla frutescens var. crispa transcriptome. PLoS ONE 2015, 10, e0129154. [Google Scholar] [CrossRef]
- Joshi, C.; Chiang, V.L. Conserved sequence motifs in plant S-adenosyl-L-methionine-dependent methyltransferases. Plant Mol. Biol. 1998, 37, 663–674. [Google Scholar] [CrossRef]
- Zubieta, C.; He, X.Z.; Dixon, R.A.; Noel, J.P. Structures of two natural product methyltransferases reveal the basis for substrate specificity in plant O-methyltransferases. Nat. Struct. Biol. 2001, 3, 271–279. [Google Scholar] [CrossRef]
- Wang, J.; Dudareva, N.; Bhakta, S.; Raguso, R.A.; Pichersky, E. Floral scent production in Clarkia breweri (Onagraceae). II. Localization and developmental modulation of the enzyme S-adenosyl-L-methionine:(iso)eugenol O-methyltransferase and phenylpropanoid emission. Plant Physiol. 1997, 114, 213–221. [Google Scholar] [CrossRef]
- Gang, D.R.; Lavid, N.; Zubieta, C.; Chen, F.; Beuerle, T.; Lewinsohn, E.; Noel, J.P.; Pichersky, E. Characterization of phenylpropene O-methyltransferases from sweet basil: Facile change of substrate specificity and convergent evolution within a plant O-methyltransferase family. Plant Cell 2002, 14, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Berim, A.; Hyatt, D.C.; Gang, D.R. A set of regioselective O-methyltransferases gives rise to the complex pattern of methoxylated flavones in sweet basil. Plant Physiol. 2012, 160, 1052–1069. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, R.X.; Gao, S.; Cheng, A.X. The functional characterization of a site-specific apigenin 4′-O-methyltransferase synthesized by the liverwort species Plagiochasma appendiculatum. Molecules 2017, 22, 769. [Google Scholar]
- Liu, Q.; Liu, L.; Zhou, J.; Shin, H.D.; Chen, R.R.; Madzak, C.; Li, J.; Du, G.; Chen, J. Biosynthesis of homoeriodictyol from eriodictyol by flavone 3’-O-methyltransferase from recombinant Yarrowia lioplytica: Heterologous expression, biochemical characterization, and optimal transformation. J. Biotechnol. 2013, 167, 472–478. [Google Scholar] [CrossRef]
- Koirala, N.; Pandey, R.P.; Parajuli, P.; Jung, H.J.; Sohng, J.K. Methylation and subsequent glycosylation of 7,8-dihydroxyflavone. J. Biotechnol. 2014, 184, 128–137. [Google Scholar] [CrossRef]
- Koirala, N.; Pandey, R.P.; Thuan, N.H.; Ghimire, G.P.; Jung, H.J.; Oh, T.J.; Sohng, J.K. Metabolic engineering of Escherichia coli for the production of isoflavonoid-4′-O-methoxides and their biological activities. Biotechnol. Appl. Biochem. 2019, 66, 484–493. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.G.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Relative Activity (%) a |
|---|---|
| Flavanone | |
| Naringenin | 97.79 |
| Eriodictyol | 53.13 |
| Taxifolin | ND b |
| Flavone | |
| Apigenin | 95.17 |
| Luteolin | 46.80 |
| Chrysin | 100.00 |
| 4′,7-Dihydroxyflavone | 62.76 |
| Flavonol | |
| 3,6-Dihydroxyflavone | ND b |
| Kaempferol | 40.34 |
| Quercetin | ND b |
| Substrate | KM (μM) | Vmax (nmol min−1 mg−1) | kcat (min−1) | kcat/KM (M−1 s−1) |
|---|---|---|---|---|
| Naringenin | 13.40 ± 1.29 | 51.65 ± 1.21 | 2.116 | 2.632 × 103 |
| Eriodictyol | 13.08 ± 1.42 | 7.84 ± 0.513 | 0.321 | 4.094 × 102 |
| Chrysin | 1.31 ± 0.25 | 2.05 ± 0.22 | 0.084 | 1.067 × 103 |
| Apigenin | 3.72 ± 0.05 | 13.32 ± 1.12 | 0.546 | 2.442 × 103 |
| Luteolin | 34.21 ± 4.65 | 4.18 ± 0.16 | 0.171 | 8.335 × 101 |
| Kaempferol | 17.88 ± 0.49 | 5.81 ± 0.26 | 0.238 | 2.219 × 102 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.L.; Lee, J.C.; Lee, K.; Lee, J.M.; Nam, H.J.; Bhoo, S.H.; Lee, T.H.; Lee, S.-W.; Cho, M.-H. Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production. Molecules 2020, 25, 4455. https://doi.org/10.3390/molecules25194455
Park HL, Lee JC, Lee K, Lee JM, Nam HJ, Bhoo SH, Lee TH, Lee S-W, Cho M-H. Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production. Molecules. 2020; 25(19):4455. https://doi.org/10.3390/molecules25194455
Chicago/Turabian StylePark, Hye Lin, Jae Chul Lee, Kyungha Lee, Jeong Min Lee, Hyo Jeong Nam, Seong Hee Bhoo, Tae Hoon Lee, Sang-Won Lee, and Man-Ho Cho. 2020. "Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production" Molecules 25, no. 19: 4455. https://doi.org/10.3390/molecules25194455
APA StylePark, H. L., Lee, J. C., Lee, K., Lee, J. M., Nam, H. J., Bhoo, S. H., Lee, T. H., Lee, S.-W., & Cho, M.-H. (2020). Biochemical Characterization of a Flavonoid O-methyltransferase from Perilla Leaves and Its Application in 7-Methoxyflavonoid Production. Molecules, 25(19), 4455. https://doi.org/10.3390/molecules25194455