
γ-Aminobutyric Acid Enhances Heat Tolerance Associated with the Change of Proteomic Profiling in Creeping Bentgrass



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
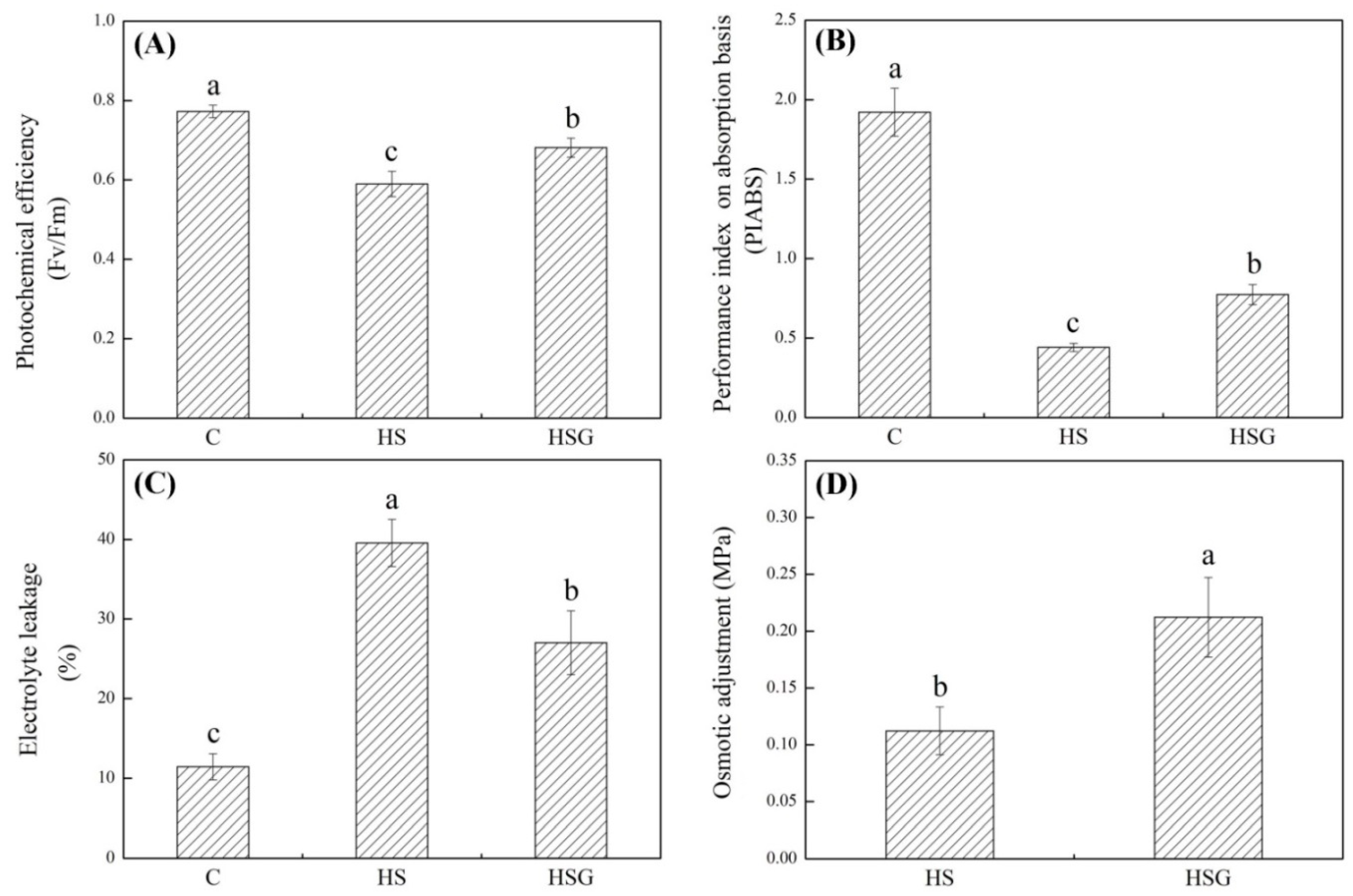
2.1. Physiological Changes Affected by Heat Stress and GABA Application
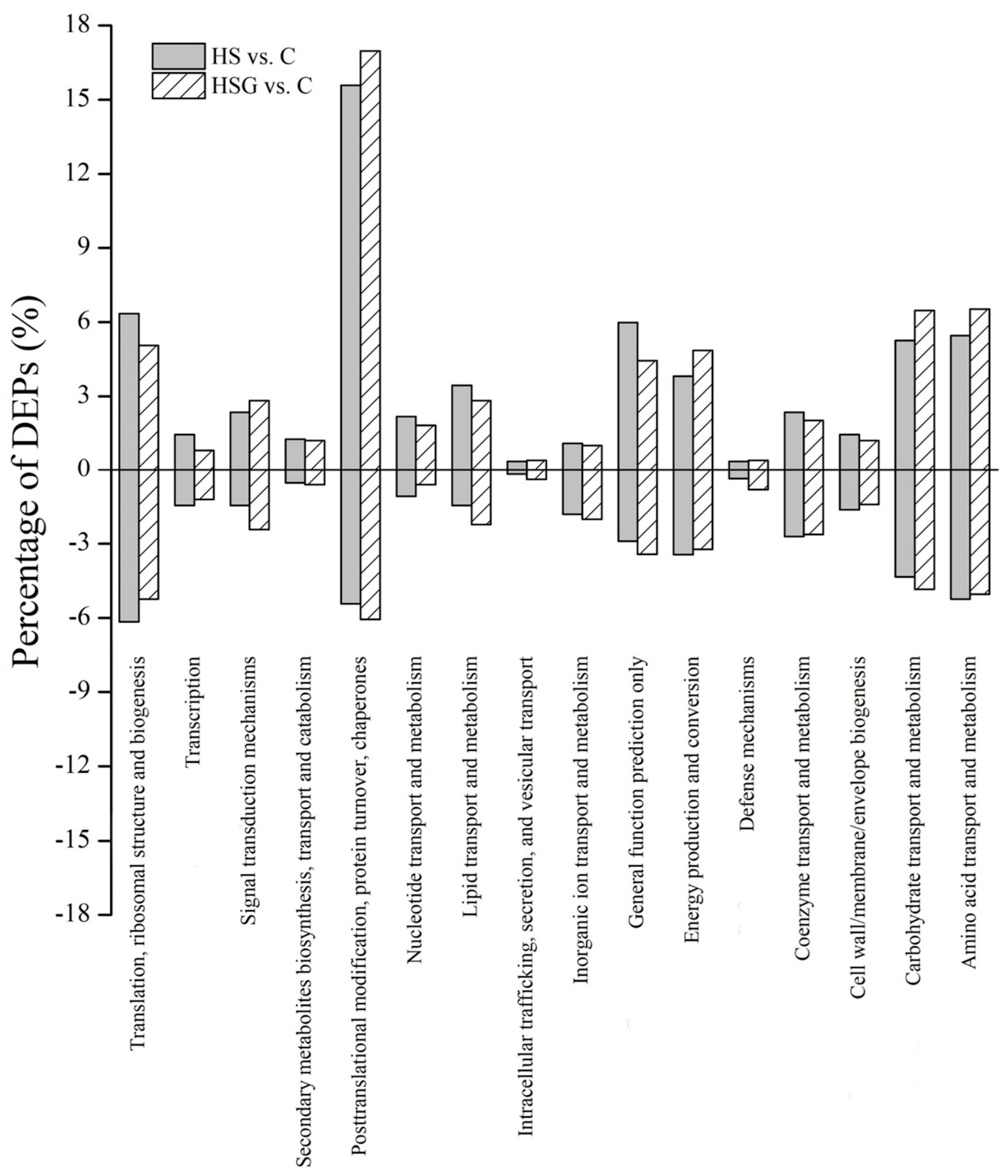
2.2. Changes of Proteins Profiling Affected by GABA and Heat Stress
2.3. The Interaction Network of DEPs
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Physiological Parameters
4.3. Reactive Oxygen Species and Antioxidant Enzyme Activities
4.4. Protein Extraction, iTRAQ Labeling, and Identification of Proteins
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sun, Q.; Miao, C.; Hanel, M.; Borthwick, A.G.; Duan, Q.; Ji, D.; Li, H. Global heat stress on health, wildfires, and agricultural crops under different levels of climate warming. Environ. Int. 2019, 128, 125–136. [Google Scholar] [CrossRef]
- Xu, Y.; Zhan, C.; Huang, B. Heat shock proteins in association with heat tolerance in grasses. Int. J. Proteom. 2011, 2011, 529648. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Ma, L.; Duan, W.; Wang, B.C.; Li, J.H.; Xu, H.G.; Yan, X.Q.; Yan, B.F.; Li, S.H.; Wang, L.J. Differential proteomic analysis of grapevine leaves by iTRAQ reveals responses to heat stress and subsequent recovery. BMC Plant Biol. 2014, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, Z.; Guan, X.; Hu, Z.; Lu, Y. Proteomic analysis of Potentilla fruticosa L. leaves by iTRAQ reveals responses to heat stress. PLoS ONE 2017, 12, e0182917. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, Y.; Fang, W.; Tao, J.; Yang, Z.; Yin, Y. iTRAQ-based proteomic and physiological analyses of mustard sprouts in response to heat stress. RSC Adv. 2020, 10, 6052–6062. [Google Scholar] [CrossRef]
- Planchet, E.; Limami, A.; D’Mello, J.P. Amino acid synthesis under abiotic stress. CAB Int. 2015, 262–276. [Google Scholar]
- Rodrigues-Corrêa, K.C.d.S.; Fett-Neto, A.G. Abiotic stresses and non-protein amino acids in plants. Crit. Rev. Plant Sci. 2019, 38, 411–430. [Google Scholar] [CrossRef]
- Ramos-Ruiz, R.; Martinez, F.; Knauf-Beiter, G. The effects of GABA in plants. Cogent Food Agric. 2019, 5, 1670553. [Google Scholar] [CrossRef]
- Al-Quraan, N.; Locy, B.; Singh, N. Heat and cold stresses phenotypes of Arabidopsis thaliana calmodulin mutants: Regulation of gamma-aminobutyric acid shunt pathway under temperature stress. Int. J. Plant Biol. 2012, 3. [Google Scholar] [CrossRef]
- Priya, M.; Sharma, L.; Kaur, R.; H., B.; Nair, R.; Siddique, K.; Nayyar, H. GABA (γ-aminobutyric acid), as a thermo-protectant, to improve the reproductive function of heat-stressed mungbean plants. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Nayyar, H.; Kaur, R.; Kaur, S.; Singh, R. γ-Aminobutyric acid (GABA) imparts partial protection from heat stress injury to rice seedlings by improving leaf turgor and upregulating osmoprotectants and antioxidants. J. Plant Growth Regul. 2014, 33, 408–419. [Google Scholar] [CrossRef]
- Luo, L.; Li, Z.; Tang, M.Y.; Cheng, B.Z.; Zeng, W.H.; Peng, Y.; Nie, G.; Zhang, X.Q. Metabolic regulation of polyamines and γ-aminobutyric acid in relation to spermidine-induced heat tolerance in white clover. Plant Biol. 2020, 22, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Huang, B. Alteration of transcripts of stress-protective genes and transcriptional factors by γ-aminobutyric acid (GABA) associated with improved heat and drought tolerance in creeping bentgrass (Agrostis stolonifera). Int. J. Mol. Sci. 2018, 19, 1623. [Google Scholar] [CrossRef] [PubMed]
- Fry, J.; Huang, B. Applied Turfgrass Science and Physiology; John Wiley Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- McCarty, B. Creeping bentgrass management—Summer stresses, weeds and selected maladies. Hort. Technol. 2001, 11, 149. [Google Scholar] [CrossRef]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.-D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Faseela, P.; Sinisha, A.K.; Brestic, M.; Puthur, J. Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice. Photosynthetica 2019, 58, 108–115. [Google Scholar]
- Yamauchi, Y.; Kimura, Y.; Akimoto, S.; Marutani, Y.; Mizutani, M.; Sugimoto, Y. Plants switch photosystem at high temperature to protect photosystem II. Nat. Preced. 2011. [Google Scholar] [CrossRef]
- Jespersen, D.; Zhang, J.; Huang, B. Chlorophyll loss associated with heat-induced senescence in bentgrass. Plant Sci. 2016, 249, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brestic, M.; Zivcak, M.; Kunderlikova, K.; Allakhverdiev, S.I. High temperature specifically affects the photoprotective responses of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 2016, 130, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Echevarría-Zomeño, S.; Fernández-Calvino, L.; Castro-Sanz, A.B.; López, J.A.; Vázquez, J.; Castellano, M.M. Dissecting the proteome dynamics of the early heat stress response leading to plant survival or death in Arabidopsis. Plant Cell Environ. 2016, 39, 1264–1278. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, X.; Huang, B. Protein changes in response to heat stress in acclimated and nonacclimated creeping bentgrass. J. Am. Soc. Hortic. Sci. 2004, 130, 521–526. [Google Scholar] [CrossRef]
- Bäurle, I. Plant heat adaptation: Priming in response to heat stress. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Jespersen, D.; Huang, B. Proteins associated with heat-induced leaf senescence in creeping bentgrass as affected by foliar application of nitrogen, cytokinins, and an ethylene inhibitor. Proteomics 2014, 15, 798–812. [Google Scholar] [CrossRef]
- Thomas, V. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar]
- Li, Z.; Cheng, B.; Zeng, W.; Liu, Z.; Peng, Y. The transcriptional and post-transcriptional regulation in perennial creeping bentgrass in response to γ-aminobutyric acid (GABA) and heat stress. Environ. Exp. Bot. 2019, 162, 515–524. [Google Scholar] [CrossRef]
- Savchenko, T.; Walley, J.W.; Chehab, E.W.; Xiao, Y.; Kaspi, R.; Pye, M.F.; Mohamed, M.E.; Lazarus, C.M.; Bostock, R.M.; Dehesh, K. Arachidonic acid: An evolutionarily conserved signaling molecule modulates plant stress signaling networks. Plant Cell 2010, 22, 3193–3205. [Google Scholar] [CrossRef]
- Garcia-Pineda, E.; Lozoya-Gloria, E. Induced gene expression of 1-aminocyclopropane-1-carboxylic acid (ACC oxidase) in pepper (Capsicum annuum L.) by arachidonic acid. Plant Sci. 1999, 145, 11–21. [Google Scholar] [CrossRef]
- Araceli, A.C.; Elda, C.M.; Edmundo, L.G.; Ernesto, G.P. Capsidiol production in pepper fruits (Capsicum annuum L.) induced by arachidonic acid is dependent of an oxidative burst. Physiol. Mol. Plant Pathol. 2007, 70, 76. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Vijayakumari, K.; Jos, T.P. γ-Aminobutyric acid (GABA) priming enhances the osmotic stress tolerance in Piper nigrum Linn. plants subjected to PEG-induced stress. Plant Growth Regul. 2016, 1, 57–67. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, W.; Meng, Y.; Xie, T.; Li, L.; Li, J.; Wei, S. γ-Aminobutyric acid imparts partial protection from salt stress injury to maize seedlings by improving photosynthesis and upregulating osmoprotectants and antioxidants. Sci. Rep. 2017, 7, 43609. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.S.; Li, Z.; Chen, Y.; Chen, M.; Li, L.C.; Ma, Y.Z. Heat shock protein 90 in plants: Molecular mechanisms and roles in stress responses. Int. J. Mol. Sci. 2012, 13, 15706–15723. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Huang, B. Differential protein expression for geothermal Agrostis scabra and turf-type Agrostis stolonifera differing in heat tolerance. Environ. Exp. Bot. 2008, 64, 58–64. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, X.; Goatley, M.; Ervin, E. Heat shock proteins in relation to heat stress tolerance of creeping bentgrass at different N levels. PLoS ONE 2014, 9, e102914. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Z.; Li, Z.; Peng, Y.; He, L. Regulation of heat shock factor pathways by γ-aminobutyric acid (GABA) associated with thermotolerance of creeping bentgrass. Int. J. Mol. Ences. 2019, 20, 4713. [Google Scholar] [CrossRef]
- Carnal, N.W.; Black, C.C. Pyrophosphate-dependent 6-phosphofructokinase, a new glycolytic enzyme in pineapple leaves. Biochem. Biophys. Res. Commun. 1979, 86, 20–26. [Google Scholar] [CrossRef]
- Pego, V.J.; Smeekens, S.C.M. Plant fructokinases: A sweet family get-together. Trends Plant Sci. 2001, 5, 531–536. [Google Scholar] [CrossRef]
- Donggiun, K. Biochemical properties and physiological functions of plant β-dfructofuranosidase. J. Life Sci. 2017, 27, 849–856. [Google Scholar]
- Li, S.; Li, T.; Kim, W.D.; Kitaoka, M.; Yoshida, S.; Nakajima, M.; Kobayashi, H. Characterization of raffinose synthase from rice (Oryza sativa L. var. Nipponbare). Biotechnol. Lett. 2007, 29, 635–640. [Google Scholar] [CrossRef]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological functions of asparagine synthetase in plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- O’Neil, J.D.; Bugno, M.; Stanley, M.S.; Barham-Morris, J.B.; Hooley, P. Cloning of a novel gene encoding a C2H2 zinc finger protein that alleviates sensitivity to abiotic stresses in Aspergillus nidulans. Mycol. Res. 2002, 106, 491–498. [Google Scholar] [CrossRef]
- Verma, S.; Chinnusamy, V.; Asif, M.; Bansal, K.C. Synergistic effect of C2H2 type Zinc-finger protein with LEA promoter to enhance abiotic stress tolerance in Brassica juncea. Plant OMICS 2013, 6, 208–214. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 357–359. [Google Scholar]
- Blum, A. Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Li, Z.; Zhang, Y.; Xu, Y.; Zhang, X.; Peng, Y.; Ma, X.; Huang, L.; Yan, Y. Physiological and iTRAQ-based proteomic analyses reveal the function of spermidine on improving drought tolerance in white clover. J. Proteome Res. 2016, 15, 1563–1579. [Google Scholar] [CrossRef]
- Philip, J.; David, B.; Hsin-Yu, C.; Matthew, F.; Weizhong, L.; Craig, M.A.; Hamish, M.W.; John, M.; Alex, M.; Gift, N. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar]
- Wei, H.D.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar]
- Andrea, F.; Damian, S.; Sune, F.; Michael, K.; Milan, S.; Alexander, R.; Jianyi, L.; Pablo, M.; Peer, B.; Christian, V.M. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zeng, W.; Cheng, B.; Huang, T.; Peng, Y.; Zhang, X. γ-Aminobutyric Acid Enhances Heat Tolerance Associated with the Change of Proteomic Profiling in Creeping Bentgrass. Molecules 2020, 25, 4270. https://doi.org/10.3390/molecules25184270
Li Z, Zeng W, Cheng B, Huang T, Peng Y, Zhang X. γ-Aminobutyric Acid Enhances Heat Tolerance Associated with the Change of Proteomic Profiling in Creeping Bentgrass. Molecules. 2020; 25(18):4270. https://doi.org/10.3390/molecules25184270
Chicago/Turabian StyleLi, Zhou, Weihang Zeng, Bizhen Cheng, Ting Huang, Yan Peng, and Xinquan Zhang. 2020. "γ-Aminobutyric Acid Enhances Heat Tolerance Associated with the Change of Proteomic Profiling in Creeping Bentgrass" Molecules 25, no. 18: 4270. https://doi.org/10.3390/molecules25184270
APA StyleLi, Z., Zeng, W., Cheng, B., Huang, T., Peng, Y., & Zhang, X. (2020). γ-Aminobutyric Acid Enhances Heat Tolerance Associated with the Change of Proteomic Profiling in Creeping Bentgrass. Molecules, 25(18), 4270. https://doi.org/10.3390/molecules25184270