Dehydroascorbic Acid Affects the Stability of Catechins by Forming Conjunctions


Abstract
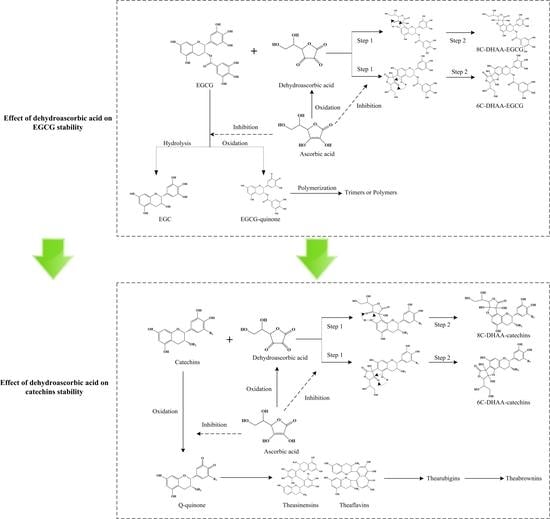
1. Introduction
2. Results
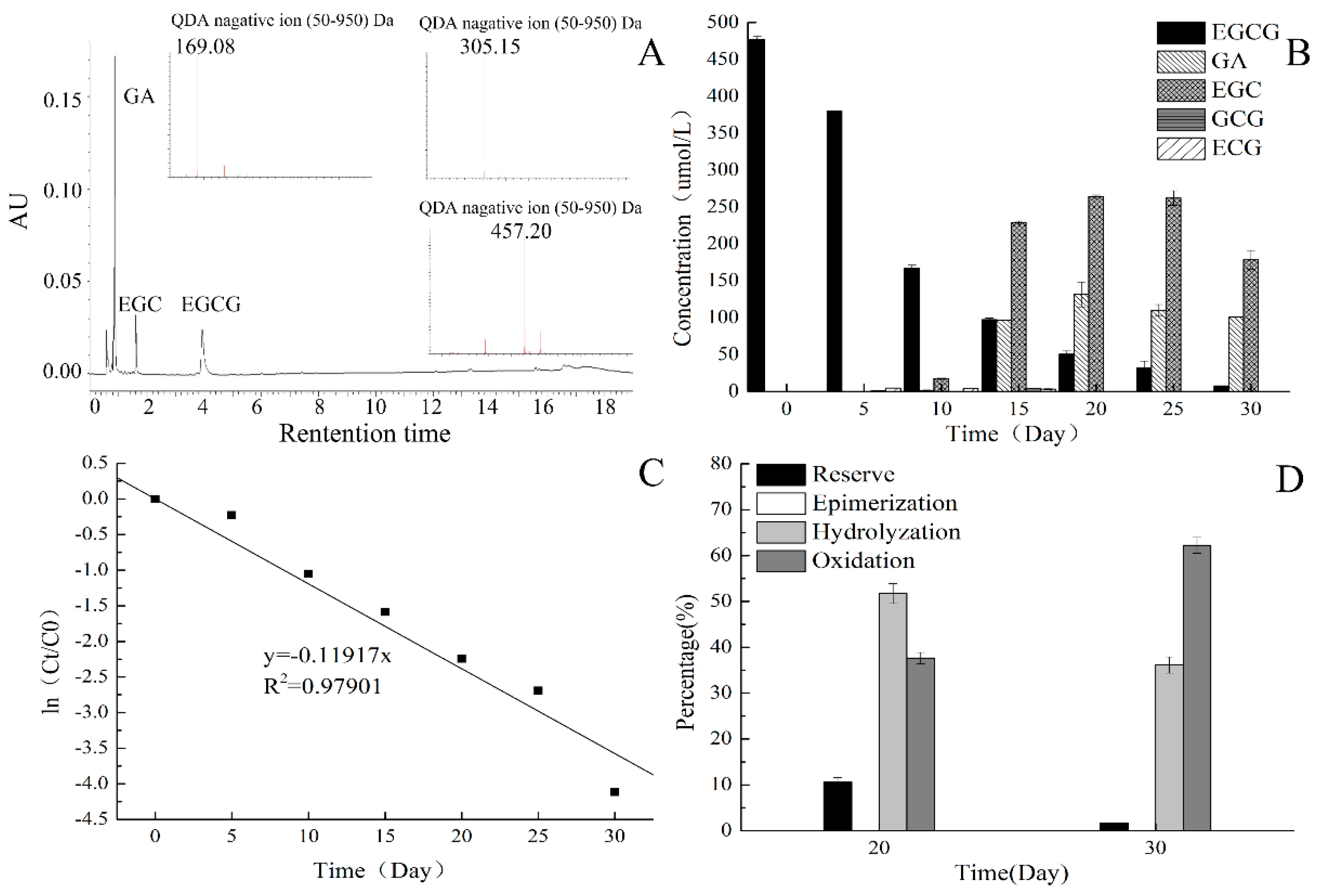
2.1. EGCG Stability at 25 °C
2.2. Effect of Different Concentration of DHAA on EGCG Stability
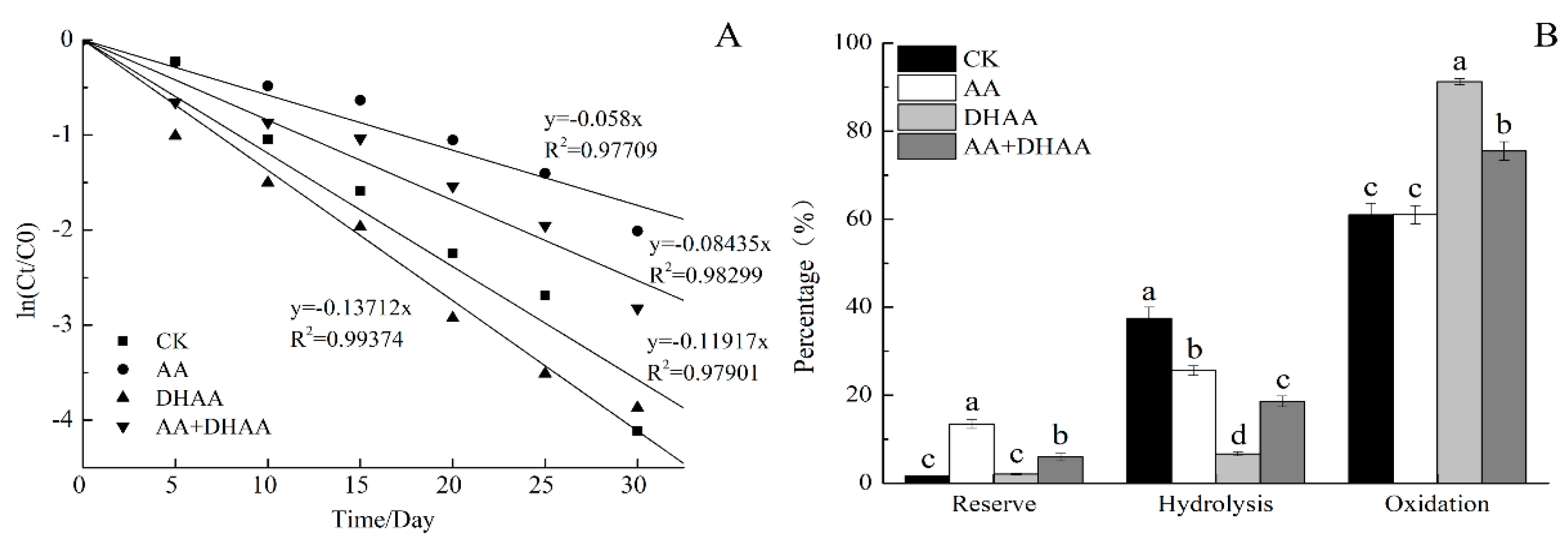
2.3. Effect of AA and DHAA on the EGCG Stability
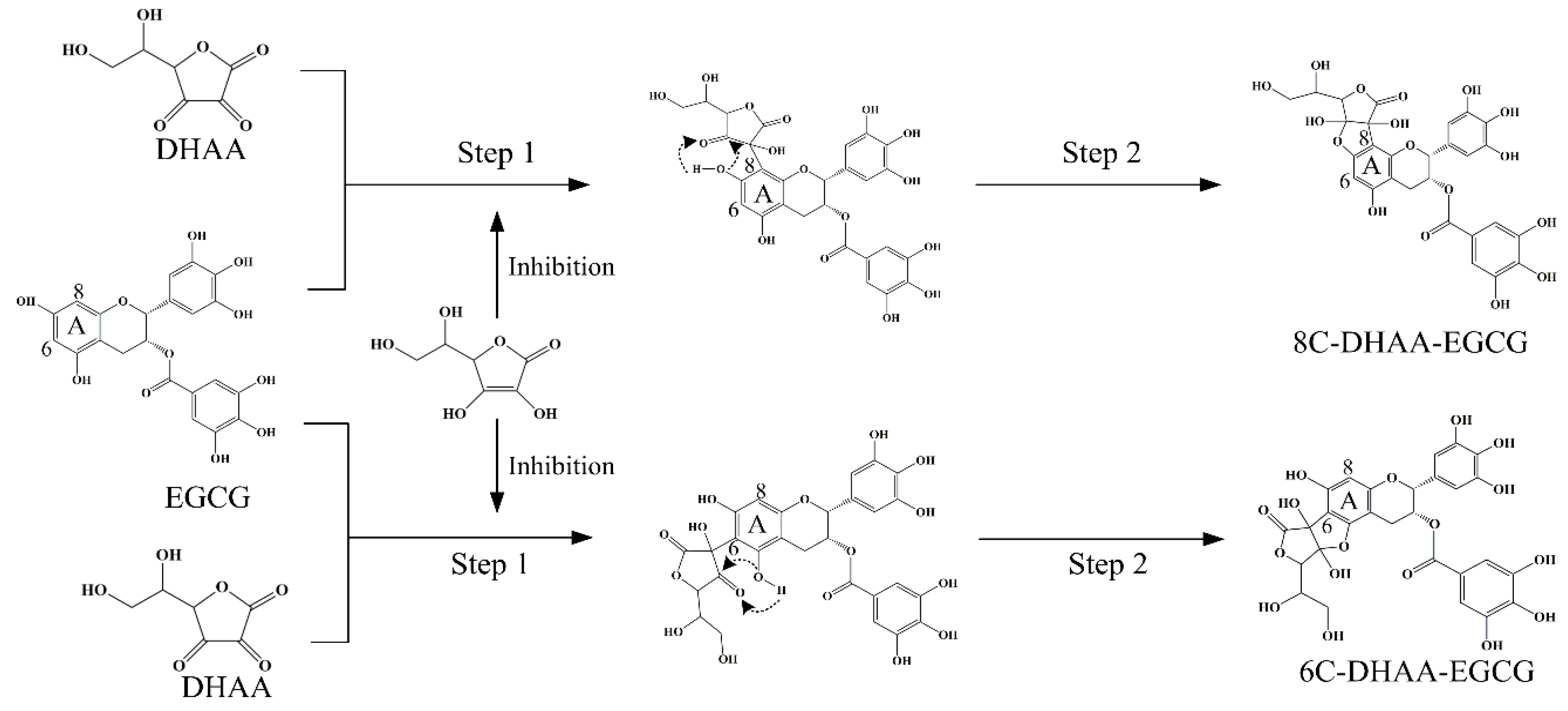
2.4. Formation of Ascorbyl Adducts of Four Catechins
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Treatment and Sample Preparation
4.2.1. Effects of Dehydroascorbic Acid Concentration on the EGCG Stability
4.2.2. Effect of Ascorbic Acid and Dehydroascorbic Acid on the EGCG Stability
4.2.3. Formation of Ascorbyl Adducts of Four Catechins
4.3. Detection Method
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Robichaud, J.L.; Noble, A.C. Astringency and bitterness of selected phenolics in wine. J. Sci. Food Agric. 1990, 53, 343–353. [Google Scholar] [CrossRef]
- Narukawa, M.; Kimata, H.; Noga, C.; Watanabe, T. Taste characterisation of green tea catechins. Int. J. Food Sci. Technol. 2010, 45, 1579–1585. [Google Scholar] [CrossRef]
- Abd-Elsalam, H.A.H.; Al-Ghobashy, M.A.; Zaazaa, H.E.; Ibrahim, M.A. Stability of catechins in green tea nutraceutical products: Application of solid phase extraction–thin layer chromatography densitometry. Food Chem. 2014, 156, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, S.A.; Balentine, D.A.; Frei, B. Antioxidants in tea. Crit. Rev. Food Sci. Nutr. 1998, 37, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Nagle, D.G.; Ferreira, D.; Zhou, Y.D. Epigallocatechin-3-gallate (EGCG): Chemical and biomedical perspectives. Phytochemistry 2006, 67, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, E.; Suzuki, H.; Menegazzi, M. Antiinflammatory Action of EGCG, the Main Component of Green Tea, through STAT-1 Inhibition. Ann. N. Y. Acad. Sci. 2002, 973, 435–437. [Google Scholar] [CrossRef]
- Nakamura, Y.; Isemura, M.; Monira, P.; Goto, S.; Ohishi, T. Anti-inflammatory Action of Green Tea. Anti-Infect. Agents Med. Chem. 2016, 15, 74–90. [Google Scholar]
- Venables, M.C.; Hulston, C.J.; Cox, H.R.; Jeukendrup, A.E. Green tea extract ingestion, fat oxidation, and glucose tolerance in healthy humans. Am. J. Clin. Nutr. 2008, 87, 778–784. [Google Scholar] [CrossRef]
- Fraga, C.G.; Motchnik, P.A.; Shigenaga, M.K.; Helbock, H.J.; Jacob, R.A.; Ames, B.N. Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc. Natl. Acad. Sci. USA 1991, 88, 11003–11006. [Google Scholar] [CrossRef]
- Fraga, D.B.; Olescowicz, G.; Moretti, M.; Sitenes, A.; Tavares, M.K.; Azevedo, D.; Colla, A.R.S.; Rodrigues, A.L.S. Anxiolytic effects of ascorbic acid and ketamine in mice. J. Psychiatr. Res. 2018, 100, 16–23. [Google Scholar] [CrossRef]
- Moretti, M.; Fraga, D.B.; Rodrigues, A.L.S. Ascorbic Acid to Manage Psychiatric Disorders. CNS Drugs 2017, 31, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Gillis, K.; Stevens, K.K.; Bell, E.; Patel, R.K.; Jardine, A.G.; Morris, S.T.W.; Schneider, M.P.; Delles, C.; Mark, P.B.; Jardine, A.G.; et al. Ascorbic acid lowers central blood pressure and asymmetric dimethylarginine in chronic kidney disease. Clin. Kidney J. 2018, 11, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Lu, J.I.; Min, C.; Yan-Qiu, C.; Cheng-Yu, H.; Jian-Qin, S. Effect of tea polyphenols and vitamin C intervention on patients with hyperuricemia and dyslipidemia. Acta Nutr. Sin. 2010, 32, 575–578. [Google Scholar]
- Bo, X.; Hui-Ge, Q.U.; Qing-Rong, H.; Chuan-Chuan, Y.U.; Bo, Z. Study on the Inhibition of Mixture of Tea-Polyphenol and Vitamin C on Helicobacter Pylori. J. Tea Sci. 2006, 26, 295–298. [Google Scholar]
- Bendich, A.; Machlin, L.J.; Scandurra, O.; Burton, G.W.; Wayner, D.D.W. The antioxidant role of vitamin C. Adv. Free Radic. Biol. Med. 1986, 2, 419–444. [Google Scholar] [CrossRef]
- Deutsch, J.C. Spontaneous Hydrolysis and Dehydration of Dehydroascorbic Acid in Aqueous Solution. Anal. Biochem. 1998, 260, 223–229. [Google Scholar] [CrossRef]
- Hung, W.L.; Wang, S.; Sang, S.; Xiaochun, W.; Yu, W.; Chi-Tang, H. Quantification of ascorbyl adducts of epigallocatechin gallate and gallocatechin gallate in bottled tea beverages. Food Chem. 2018, 261, 246–252. [Google Scholar] [CrossRef]
- Su, Y.L.; Leung, L.K.; Huang, Y.; Chen, Z.Y. Stability of tea theaflavins and catechins. Food Chem. 2003, 83, 189–195. [Google Scholar]
- Zhu, Y.; Zhao, Y.; Wang, P.; Ahmedna, M.; Ho, C.T.; Sang, S. Tea Flavanols Block Advanced Glycation of Lens Crystallins Induced by Dehydroascorbic Acid. Chem. Res. Toxicol. 2015, 28, 135–143. [Google Scholar] [CrossRef]
- West, M.E.; Mauer, L.J. Color and Chemical Stability of a Variety of Anthocyanins and Ascorbic Acid in Solution and Powder Forms. J. Agric. Food Chem. 2013, 61, 4169–4179. [Google Scholar] [CrossRef]
- Liu, Z.H.; Cao, Y.H.; Xu, Y.; Li, C.W.; Si, W.Y. Assessment of the Content and Stability of Tea Polyphenols, Free Amino Acids and Vitamin C in Four Kinds of Tea Beverages. Sic. Technol. Food Ind. 2019, 40, 38–44. [Google Scholar]
- Sharma, A.; Zhou, W. A stability study of green tea catechins during the biscuit making process. Food Chem. 2011, 126, 568–573. [Google Scholar] [CrossRef]
- Deutsch, J.C. Ascorbic Acid Oxidtion by Hydrogen Peroxide. Anal. Biochem. 1998, 255, 1–7. [Google Scholar]
- Yong-Quan, X.U.; Gen-Sheng, C.; Xiao-Yu, Z.; Fang, W.; Yu-Zhou, Y.; Hai-Bo, Y. Effect of Ca2+ on the Chemical Components and Sensory Quality of Extracted Green Tea Infusion. J. Tea Sci. 2011, 31, 230–236. [Google Scholar]
- Friedman, M.; Levin, C.E.; Lee, S.-U.; Kozukue, N. Stability of Green Tea Catechins in Commercial Tea Leaves during Storage for 6 Months. J. Food Sci. 2010, 74, H47–H51. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, W.; Jiang, X. Reaction Kinetics of Degradation and Epimerization of Epigallocatechin Gallate (EGCG) in Aqueous System over a Wide Temperature Range. J. Agric. Food Chem. 2008, 56, 2694–2701. [Google Scholar] [CrossRef]
- Mochizuki, M.; Yamazaki, S.I.; Kano, K.; Ikeda, T. Kinetic analysis and mechanistic aspects of autoxidation of catechins. Biochim. Biophys. Acta 2002, 1569, 35–44. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Scan Mode | Precursor Ion | Fragment Ion 1 | Fragment Ion 2 |
|---|---|---|---|---|
| AA | Negative ion | 175.01 | 114.97 | / |
| DHAA | Negative ion | 173.09 | 110.98 | / |
| EGCG | Negative ion | 457.13 | 305.01 | 169.07 |
| ECG | Negative ion | 305.15 | / | / |
| EC | Negative ion | 289.10 | / | / |
| EGC | Negative ion | 441.18 | / | / |
| DHAA–EGCG | Negative ion | 631.22 | 495.20 | 343.14 |
| DHAA–ECG | Negative ion | 615.21 | 479.10 | 327.12 |
| DHAA–EC | Negative ion | 463.20 | 327.12 | 191.04 |
| DHAA–EGC | Negative ion | 479.07 | 451.29 | 343.14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Wang, W.; Zhang, J.; Wang, W.; Ni, D.; Jiang, H. Dehydroascorbic Acid Affects the Stability of Catechins by Forming Conjunctions. Molecules 2020, 25, 4076. https://doi.org/10.3390/molecules25184076
Chen L, Wang W, Zhang J, Wang W, Ni D, Jiang H. Dehydroascorbic Acid Affects the Stability of Catechins by Forming Conjunctions. Molecules. 2020; 25(18):4076. https://doi.org/10.3390/molecules25184076
Chicago/Turabian StyleChen, Lin, Wei Wang, Jianyong Zhang, Weiwei Wang, Dejiang Ni, and Heyuan Jiang. 2020. "Dehydroascorbic Acid Affects the Stability of Catechins by Forming Conjunctions" Molecules 25, no. 18: 4076. https://doi.org/10.3390/molecules25184076
APA StyleChen, L., Wang, W., Zhang, J., Wang, W., Ni, D., & Jiang, H. (2020). Dehydroascorbic Acid Affects the Stability of Catechins by Forming Conjunctions. Molecules, 25(18), 4076. https://doi.org/10.3390/molecules25184076