The Genome Analysis of Methylobacterium populi YC-XJ1 with Diverse Xenobiotics Biodegrading Capacity and Degradation Characteristics of Related Hydrolase


Abstract
1. Introduction
2. Results
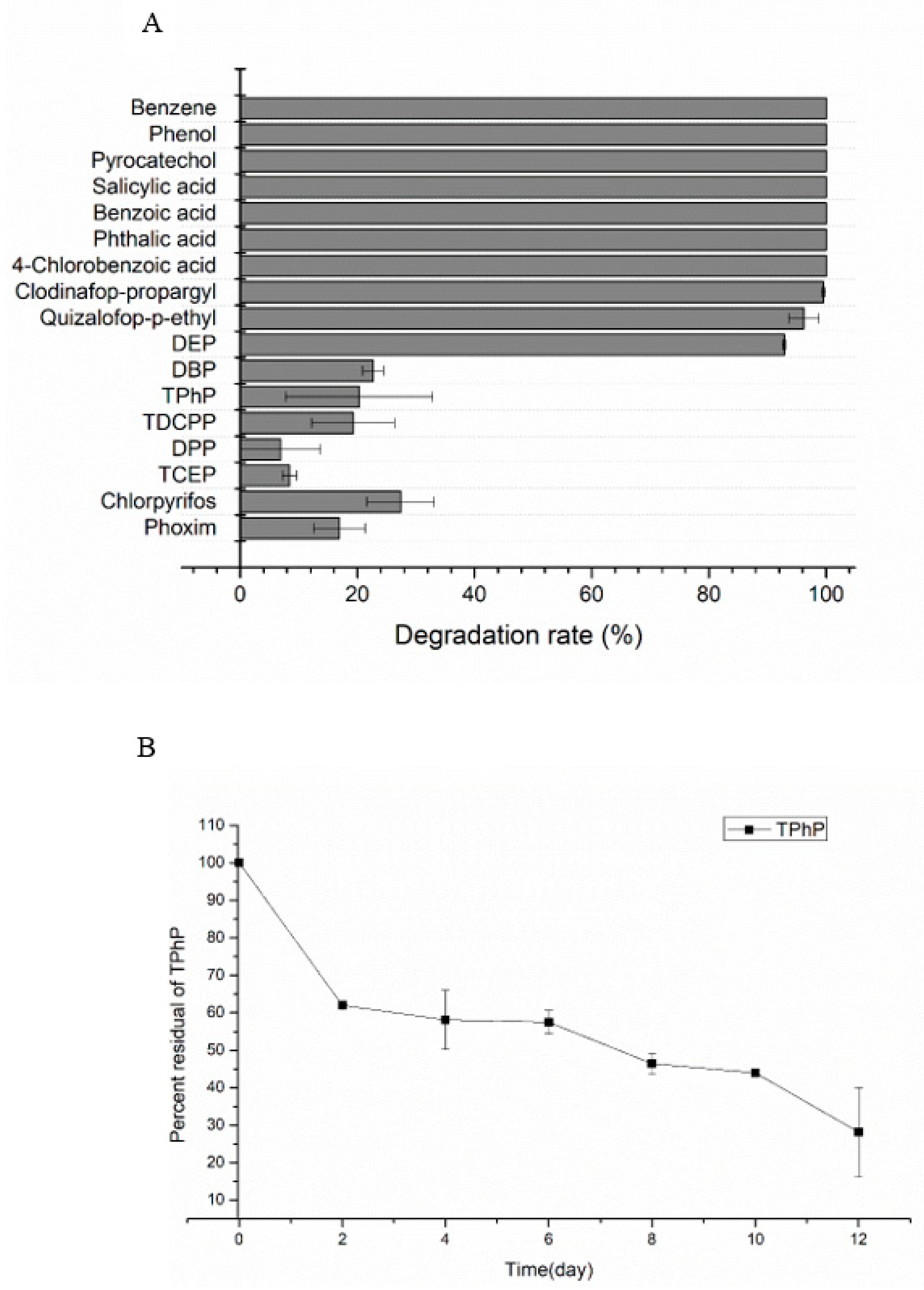
2.1. Substrates Utilization Test
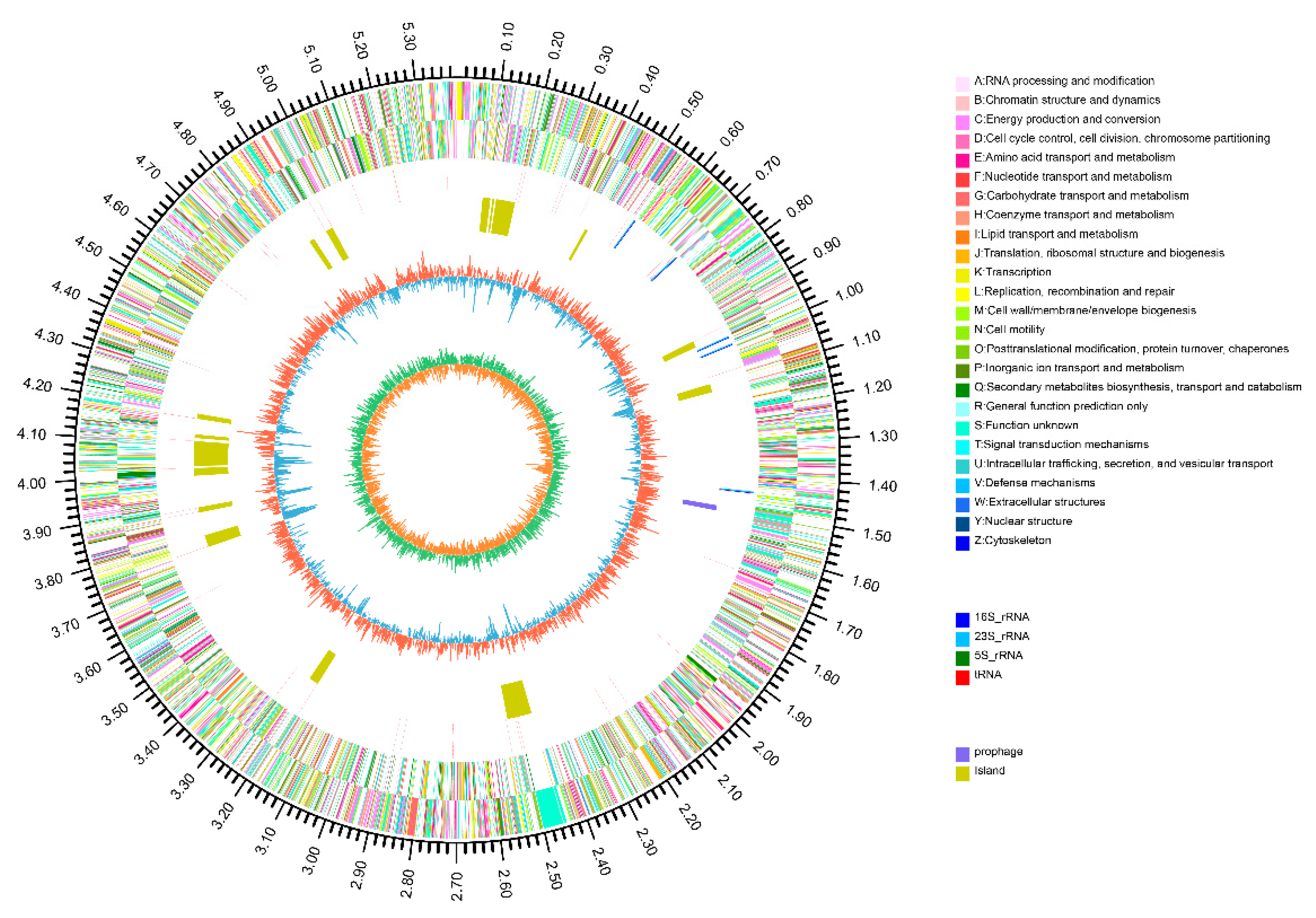
2.2. Genome Properties
2.3. ANI Analysis
2.4. Collinearity Analysis
2.5. Gene Function Analysis
2.5.1. COG Categories Reveal Metabolism-Related Functions
2.5.2. GO Terms Reveal Biological Relevance
2.5.3. CAZy Families Reveal Carbohydrate-Active Enzyme Genes
2.5.4. Analysis of Relative Metabolic Pathways
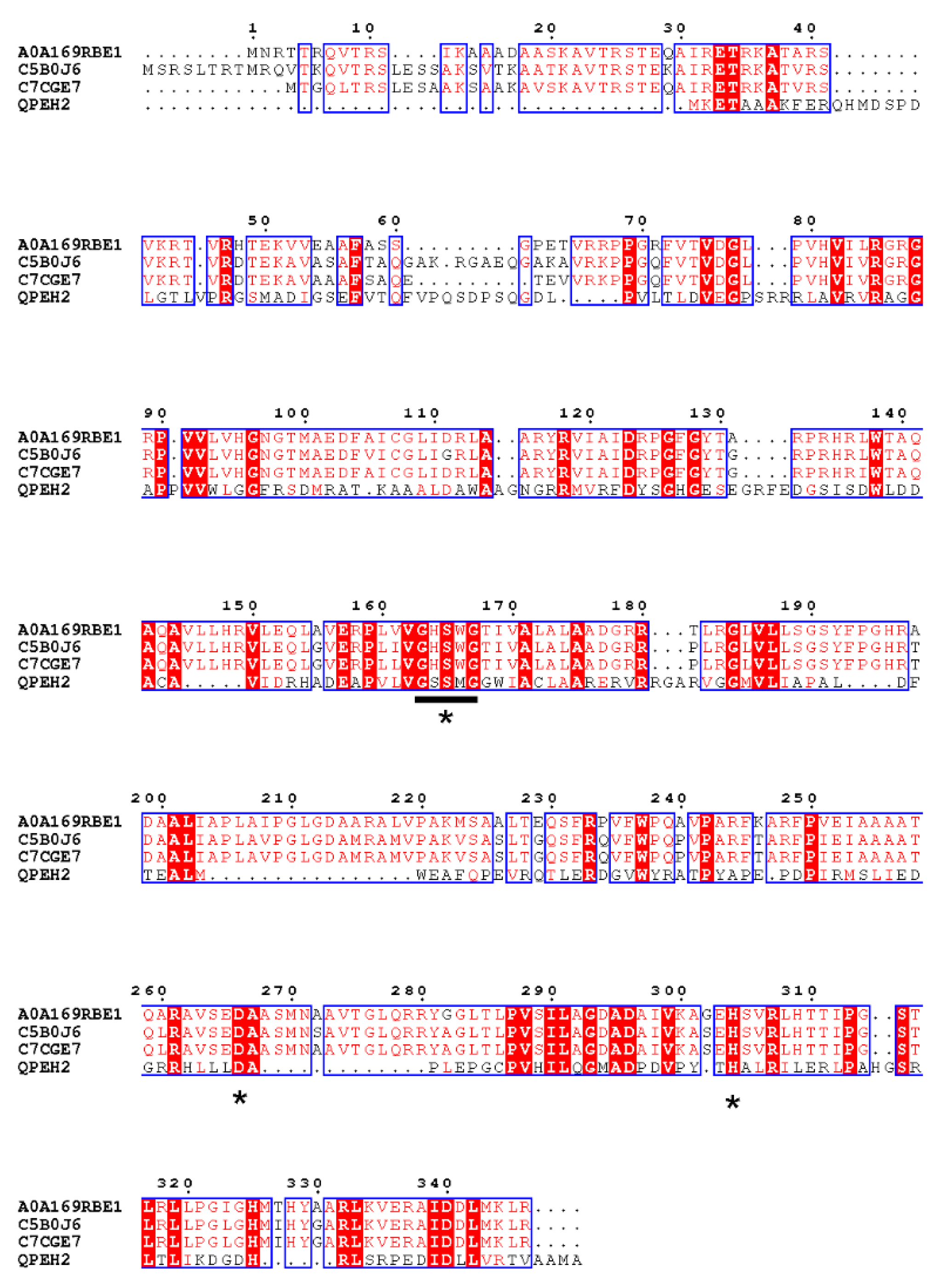
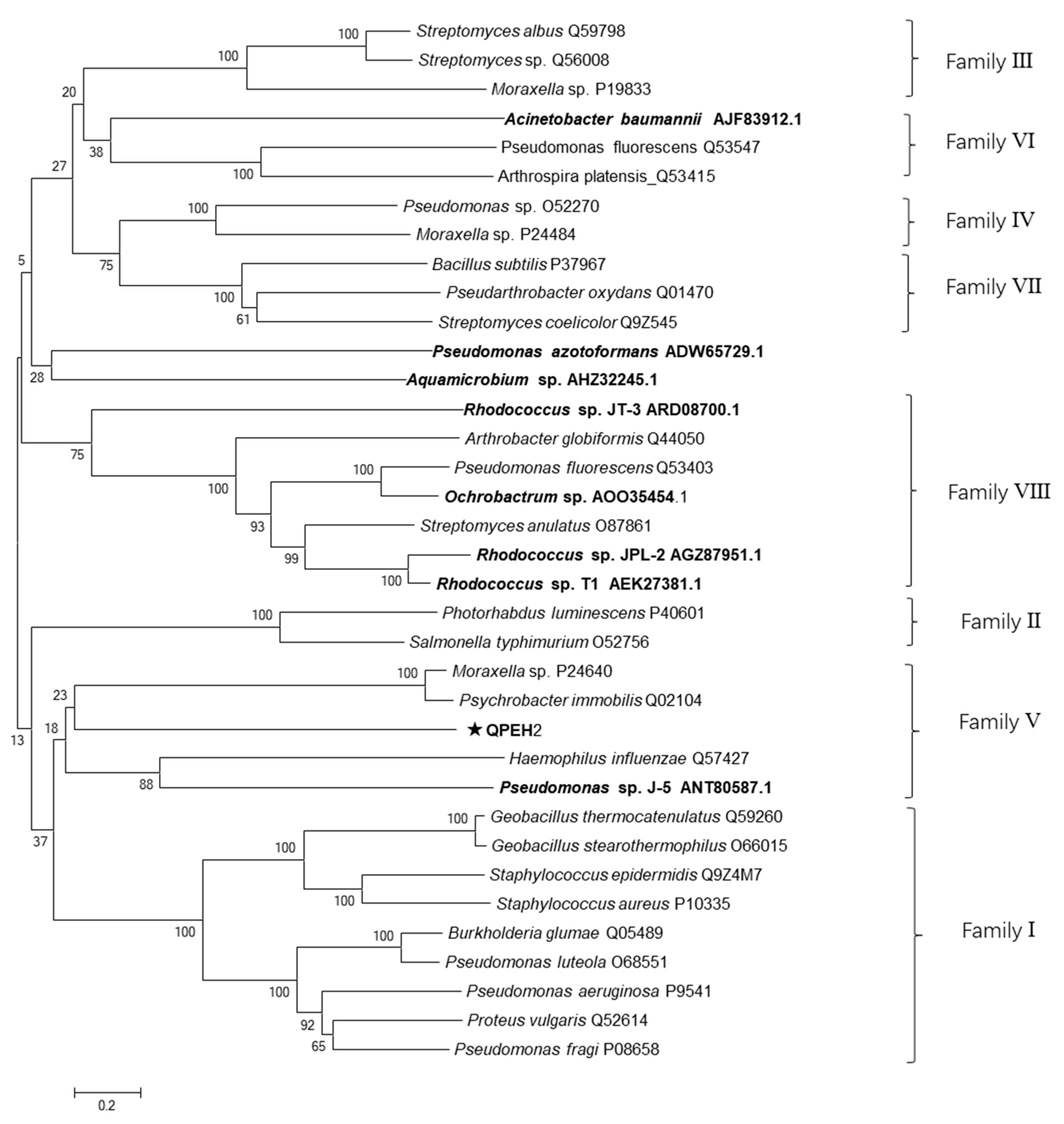
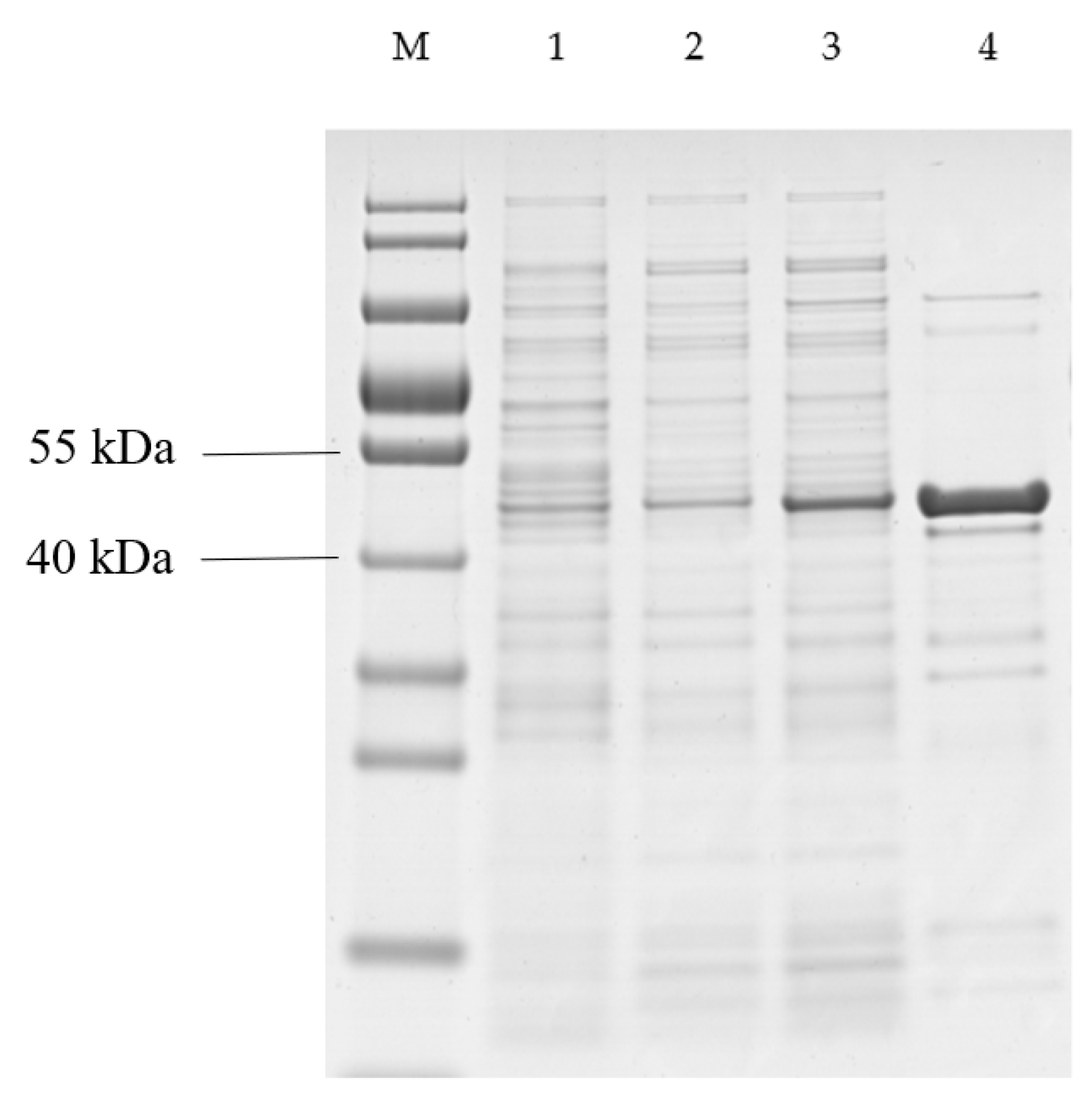
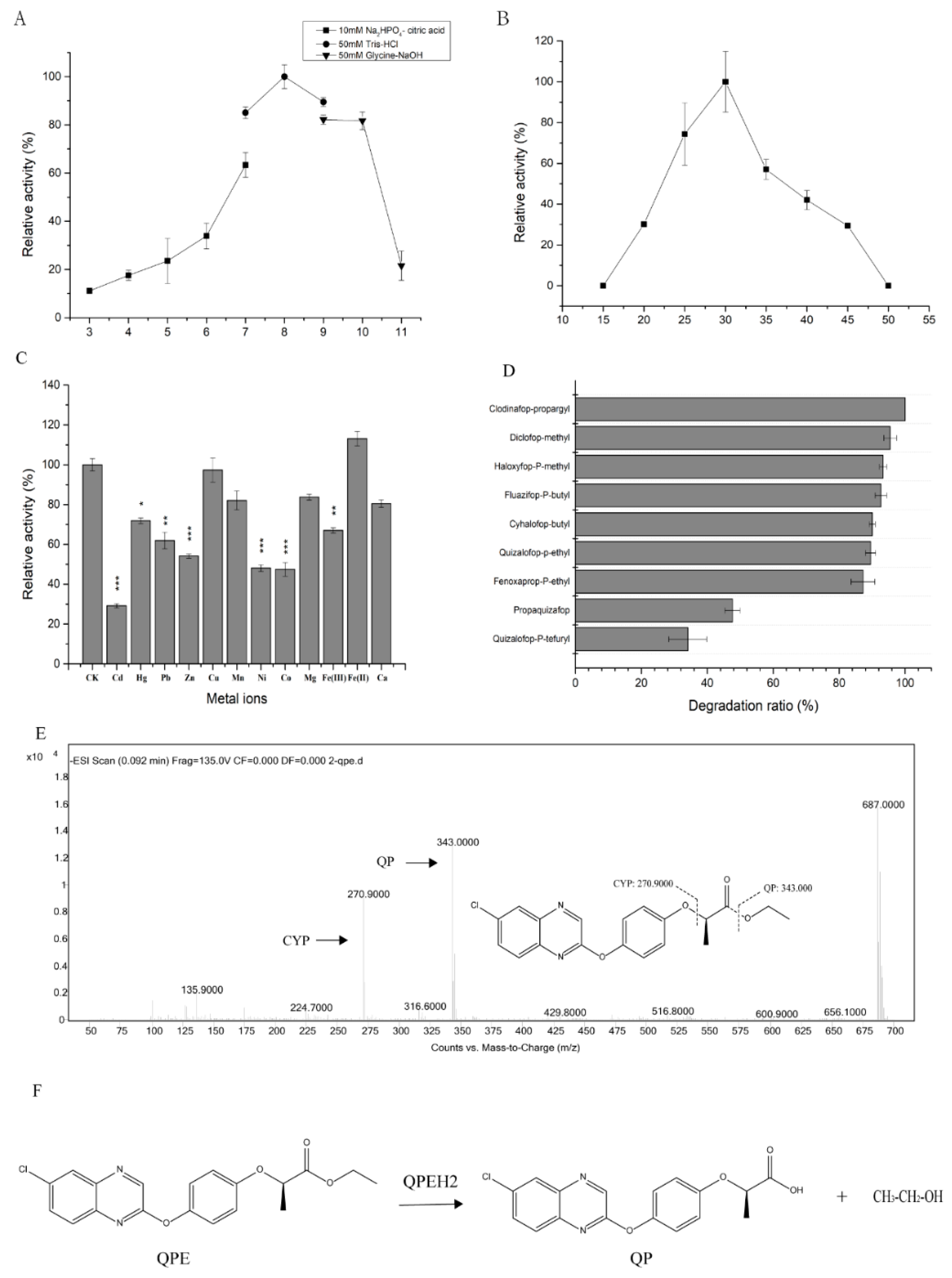
2.6. Characteristics of QPEH2 Hydrolase
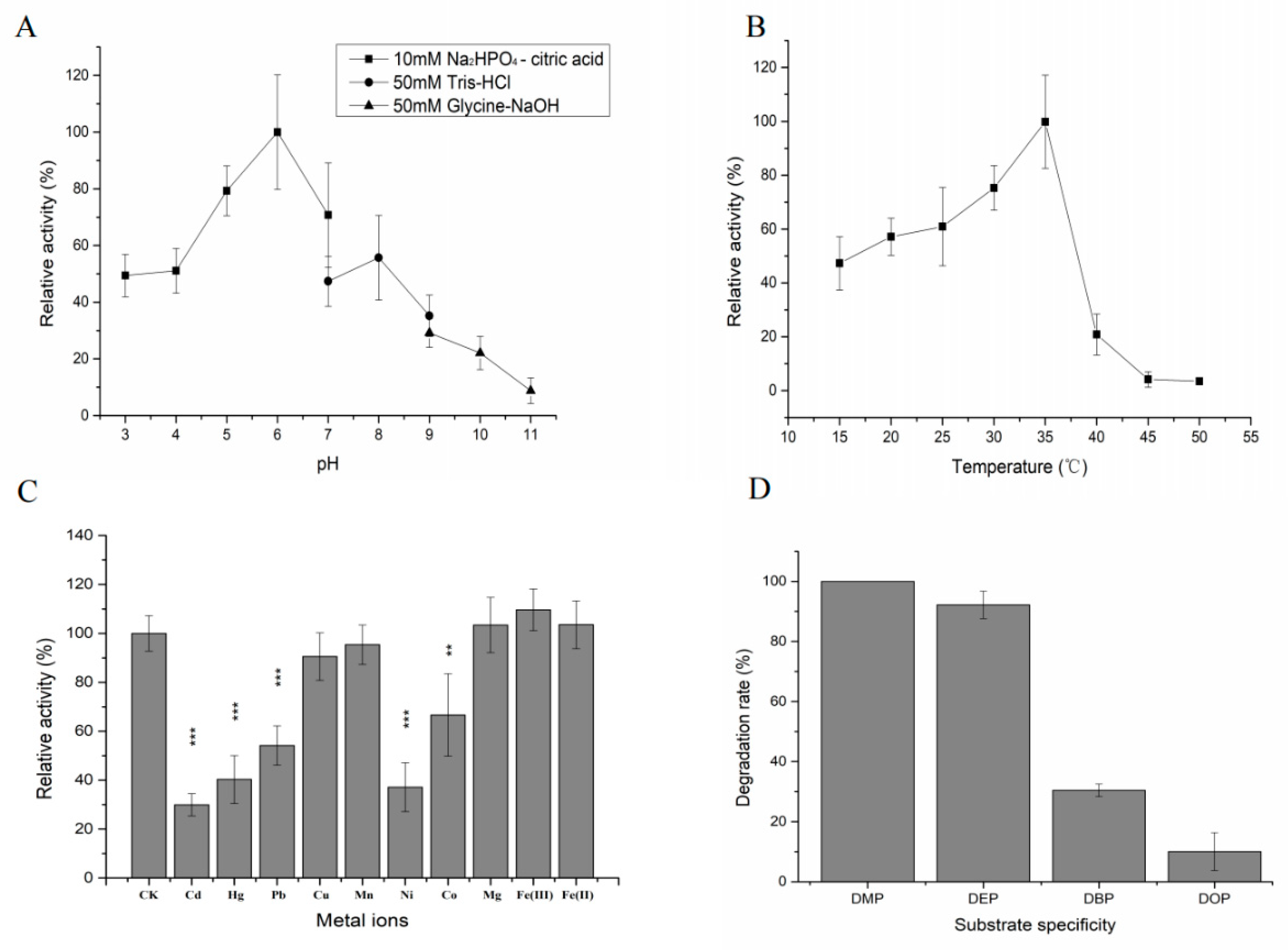
2.7. Enzymatic Characteristics of DEPH1
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Media
4.3. Analytical Methods
4.4. Substrates Utilization Tests
4.5. Genomic DNA Extractions, Library Construction and Sequencing
4.6. Gene Prediction and Annotation
4.7. Average Nucleotide Identity and Alignment Fraction
4.8. Collinearity Analysis
4.9. Sequence Analysis of qpeh2 and deph1
4.10. Cloning, Expression and Purification of the Recombinant QPEH2 and DEPH1
4.11. Biochemical Properties of the Purified Recombinant QPEH2 and DEPH1
4.12. Determination of Kinetic Constants
4.13. Accession Numbers
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mesnage, R.; Biserni, M.; Wozniak, E.; Xenakis, T.; Mein, C.A.; Antoniou, M.N. Comparison of transcriptome responses to glyphosate, isoxaflutole, quizalofop-p-ethyl and mesotrione in the HepaRG cell line. Toxicol. Rep. 2018, 5, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Elefsiniotis, I.S.; Liatsos, G.D.; Stamelakis, D.; Moulakakis, A. Case report: Mixed cholestatic/hepatocellular liver injury induced by the herbicide quizalofop-p-ethyl. Environ. Health Perspect. 2007, 115, 1479–1481. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.Z.; Qi, S.Z.; Cao, F.J.; Mu, X.Y.; Yang, Y.; Wang, C. Quizalofop-P-ethyl exposure increases estrogen axis activity in male and slightly decreases estrogen axis activity in female zebrafish (Danio rerio). Aquatic Toxicol. 2017, 183, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Doganlar, Z.B. Quizalofop-p-ethyl-induced phytotoxicity and genotoxicity in Lemna minor and Lemna gibba. J. Environ. Sci. Health 2012, 47, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Bhaduri, D.; Pipariya, A.; Jain, N.K. Influence of imazethapyr and quizalofop-p-ethyl application on microbial biomass and enzymatic activity in peanut grown soil. Environ. Sci. Pollut. Res. Int. 2016, 23, 23758–23771. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.W.; Wen, Z.D. Phthalate esters in the environment: A critical review of their occurrence, biodegradation, and removal during wastewater treatment processes. Sci. Total Environ. 2016, 541, 986–1001. [Google Scholar] [CrossRef]
- Feng, L.; Liu, H.; Cheng, D.; Mao, X.; Wang, Y.; Wu, Z.; Wu, Q. Characterization and Genome Analysis of a Phthalate Esters-Degrading Strain Sphingobium yanoikuyae SHJ. BioMed Res. Int. 2018, 2018, 3917054. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, S.L.; Fan, C.Q. Atmospheric distribution of particulate- and gas-phase phthalic esters (PAEs) in a Metropolitan City, Nanjing, East China. Chemosphere 2008, 72, 1567–1572. [Google Scholar] [CrossRef]
- Teil, M.J.; Blanchard, M.; Chevreuil, M. Atmospheric fate of phthalate esters in an urban area (Paris-France). Sci. Total Environ. 2006, 354, 212–223. [Google Scholar] [CrossRef]
- Xie, Z.; Ebinghaus, R.; Temme, C.; Lohmann, R.; Caba, A.; Ruck, W. Occurrence and air-sea exchange of phthalates in the Arctic. Environ. Sci. Technol. 2007, 41, 4555–4560. [Google Scholar] [CrossRef]
- Pereira, C.; Mapuskar, K.; Rao, C.V. Chronic toxicity of diethyl phthalate in male Wistar rats--a dose-response study. Regul. Toxicol. Pharmacol. RTP 2006, 45, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Rao, C.V. Toxicity study of maternal transfer of polychlorinated biphenyls and diethyl phthalate to 21-day-old male and female weanling pups of Wistar rats. Ecotoxicol. Environ. Saf. 2007, 68, 118–125. [Google Scholar] [CrossRef]
- Ghorpade, N.; Mehta, V.; Khare, M.; Sinkar, P.; Krishnan, S.; Rao, C.V. Toxicity study of diethyl phthalate on freshwater fish Cirrhina mrigala. Ecotoxicol. Environ. Saf. 2002, 53, 255–258. [Google Scholar] [CrossRef]
- Sonde, V.; D’Souza, A.; Tarapore, R.; Pereira, L.; Khare, M.P.; Sinkar, P.; Krishnan, S.; Rao, C.V. Simultaneous administration of diethylphthalate and ethyl alcohol and its toxicity in male Sprague-Dawley rats. Toxicology 2000, 147, 23–31. [Google Scholar] [CrossRef]
- Xu, H.; Shao, X.; Zhang, Z.; Zou, Y.; Chen, Y.; Han, S.; Wang, S.; Wu, X.; Yang, L.; Chen, Z. Effects of di-n-butyl phthalate and diethyl phthalate on acetylcholinesterase activity and neurotoxicity related gene expression in embryonic zebrafish. Bull. Environ. Contam. Toxicol. 2013, 91, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Marklund, A.; Andersson, B.; Haglund, P. Screening of organophosphorus compounds and their distribution in various indoor environments. Chemosphere 2003, 53, 1137–1146. [Google Scholar] [CrossRef]
- Rodriguez, I.; Calvo, F.; Quintana, J.B.; Rubi, E.; Rodil, R.; Cela, R. Suitability of solid-phase microextraction for the determination of organophosphate flame retardants and plasticizers in water samples. J. Chromatogr. A 2006, 1108, 158–165. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, J.; Gan, Z.; Yan, Y.; Ding, S.; Su, S.; Bao, X. Grain size distribution and exposure evaluation of organophosphorus and brominated flame retardants in indoor and outdoor dust and PM10 from Chengdu, China. J. Hazard. Mater. 2019, 365, 280–288. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, W.; Kannan, K.; Moon, H.B. Occurrence and exposure assessment of organophosphate flame retardants (OPFRs) through the consumption of drinking water in Korea. Water Res. 2016, 103, 182–188. [Google Scholar] [CrossRef]
- Wolschke, H.; Suhring, R.; Xie, Z.; Ebinghaus, R. Organophosphorus flame retardants and plasticizers in the aquatic environment: A case study of the Elbe River, Germany. Environ. Pollut. (Barking, Essex 1987) 2015, 206, 488–493. [Google Scholar] [CrossRef]
- Isales, G.M.; Hipszer, R.A.; Raftery, T.D.; Chen, A.; Stapleton, H.M.; Volz, D.C. Triphenyl phosphate-induced developmental toxicity in zebrafish: Potential role of the retinoic acid receptor. Aquat. Toxicol. 2015, 161, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Jin, Y.; Wu, Y.; Liu, L.; Fu, Z. Exposure of male mice to two kinds of organophosphate flame retardants (OPFRs) induced oxidative stress and endocrine disruption. Environ. Toxicol. Pharmacol. 2015, 40, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques-classification based on site of application: Principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 2016, 32, 180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, M.; Li, J.; Wang, G.; Liu, Y. Purification and properties of a novel quizalofop-p-ethyl-hydrolyzing esterase involved in quizalofop-p-ethyl degradation by Pseudomonas sp. J-2. Microb. Cell Factories 2017, 16, 80. [Google Scholar] [CrossRef]
- Wang, C.; Qiu, J.; Yang, Y.; Zheng, J.; He, J.; Li, S. Identification and characterization of a novel carboxylesterase (FpbH) that hydrolyzes aryloxyphenoxypropionate herbicides. Biotechnol. Lett. 2017, 39, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Jiang, S.; Shi, K.; Wang, F.; Li, S.; Zhou, J.; Huang, F.; Wang, Y.; Zheng, Y.; Hou, Y.; et al. Biodegradation of fenoxaprop-P-ethyl (FE) by Acinetobacter sp. strain DL-2 and cloning of FE hydrolase gene afeH. Bioresour. Technol. 2015, 186, 114–121. [Google Scholar] [CrossRef]
- Hou, Y.; Tao, J.; Shen, W.; Liu, J.; Li, J.; Li, Y.; Cao, H.; Cui, Z. Isolation of the fenoxaprop-ethyl (FE)-degrading bacterium Rhodococcus sp. T1, and cloning of FE hydrolase gene feh. FEMS Microbiol. Lett. 2011, 323, 196–203. [Google Scholar] [CrossRef]
- Wu, J.; Liao, X.; Yu, F.; Wei, Z.; Yang, L. Cloning of a dibutyl phthalate hydrolase gene from Acinetobacter sp. strain M673 and functional analysis of its expression product in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 2483–2491. [Google Scholar] [CrossRef]
- Fang, H.H.; Liang, D.; Zhang, T. Aerobic degradation of diethyl phthalate by Sphingomonas sp. Bioresour. Technol. 2007, 98, 717–720. [Google Scholar] [CrossRef]
- Jin, L.; Sun, X.; Zhang, X.; Guo, Y.; Shi, H. Co-metabolic biodegradation of DBP by Paenibacillus sp. S-3 and H-2. Curr. Microbiol. 2014, 68, 708–716. [Google Scholar] [CrossRef]
- Wu, X.L.; Wang, Y.Y.; Liang, R.X.; Dai, Q.Y.; Chao, W.L. Degradation of di-n-butyl phthalate by newly isolated Ochrobactrum sp. Bull. Environ. Contam. Toxicol. 2010, 85, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Yin, H.; Peng, H.; Lu, G.; Dang, Z. Bioremediation of triphenyl phosphate by Brevibacillus brevis: Degradation characteristics and role of cytochrome P450 monooxygenase. Sci. Total. Environ. 2018, 627, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Satake, I.; Konuma, I.; Kawashima, K.; Kawasaki, M.; Mori, S.; Morino, J.; Mori, J.; Xu, H.; Abe, K.; et al. Isolation and identification of persistent chlorinated organophosphorus flame retardant-degrading bacteria. Appl. Environ. Microbiol. 2010, 76, 5292–5296. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, Y.; Nakamura, S.; Nishio, T.; Fukunaga, I. Isolation of aryl-phosphate ester-degrading bacterium from leachate of a sea-based waste disposal site. J. Biosci. Bioeng. 2004, 98, 464–469. [Google Scholar] [CrossRef]
- Wang, J.; Khokhar, I.; Ren, C.; Li, X.; Wang, J.; Fan, S.; Jia, Y.; Yan, Y. Characterization and 16S metagenomic analysis of organophosphorus flame retardants degrading consortia. J. Hazard. Mater. 2019, 380, 120881. [Google Scholar] [CrossRef]
- Chaudhary, N.; Gonzalez, E.; Chang, S.H.; Geng, F.; Rafii, S.; Altorki, N.K.; McGraw, T.E. Adenovirus Protein E4-ORF1 Activation of PI3 Kinase Reveals Differential Regulation of Downstream Effector Pathways in Adipocytes. Cell Rep. 2016, 17, 3305–3318. [Google Scholar] [CrossRef]
- Roth, C.; Weizenmann, N.; Bexten, N.; Saenger, W.; Zimmermann, W. Amylose recognition and ring-size determination of amylomaltase. Sci Adv. 2017, 3, e1601386. [Google Scholar] [CrossRef]
- Watanabe, M.; Fukada, H.; Inoue, H.; Ishikawa, K. Crystal structure of an acetylesterase from Talaromyces cellulolyticus and the importance of a disulfide bond near the active site. FEBS Lett. 2015, 589, 1200–1206. [Google Scholar] [CrossRef]
- Nakamura, T.; Fahmi, M. Genome-Wide Analysis of Whole Human Glycoside Hydrolases by Data-Driven Analysis in Silico. Int. J. Mol. Sci. 2019, 20, 6290. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Dourado, M.N.; Camargo Neves, A.A.; Santos, D.S.; Araujo, W.L. Biotechnological and agronomic potential of endophytic pink-pigmented methylotrophic Methylobacterium spp. BioMed. Res. Int. 2015, 2015, 909016. [Google Scholar] [CrossRef] [PubMed]
- McDonald, I.R.; Warner, K.L.; McAnulla, C.; Woodall, C.A.; Oremland, R.S.; Murrell, J.C. A review of bacterial methyl halide degradation: Biochemistry, genetics and molecular ecology. Environ. Microbiol. 2002, 4, 193–203. [Google Scholar] [CrossRef]
- Ventorino, V.; Sannino, F.; Piccolo, A.; Cafaro, V.; Carotenuto, R.; Pepe, O. Methylobacterium populi VP2: Plant growth-promoting bacterium isolated from a highly polluted environment for polycyclic aromatic hydrocarbon (PAH) biodegradation. Sci. World J. 2014, 2014, 931793. [Google Scholar] [CrossRef] [PubMed]
- Eevers, N.; Van Hamme, J.D.; Bottos, E.M.; Weyens, N.; Vangronsveld, J. Draft Genome Sequence of Methylobacterium radiotolerans, a DDE-Degrading and Plant Growth-Promoting Strain Isolated from Cucurbita pepo. Genome Announc. 2015, 3, e00488-15. [Google Scholar] [CrossRef] [PubMed]
- Fournier, D.; Trott, S.; Hawari, J.; Spain, J. Metabolism of the aliphatic nitramine 4-nitro-2,4-diazabutanal by Methylobacterium sp. strain JS178. Appl. Environ. Microbiol. 2005, 71, 4199–4202. [Google Scholar] [CrossRef]
- Srivastva, N.; Vishwakarma, P.; Bhardwaj, Y.; Singh, A.; Manjunath, K.; Dubey, S.K. Kinetic and molecular analyses reveal isoprene degradation potential of Methylobacterium sp. Bioresour. Technol. 2017, 242, 87–91. [Google Scholar] [CrossRef]
- Zhao, B.; Hua, X.; Wang, F.; Dong, W.; Li, Z.; Yang, Y.; Cui, Z.; Wang, M. Biodegradation of propyzamide by Comamonas testosteroni W1 and cloning of the propyzamide hydrolase gene camH. Bioresour. Technol. 2015, 179, 144–149. [Google Scholar] [CrossRef]
- Liu, H.M.; Lou, X.; Ge, Z.J. Isolation of an aryloxyphenoxy propanoate (AOPP) herbicide-degrading strain Rhodococcus ruber JPL-2 and the cloning of a novel carboxylesterase gene (feh). Braz. J. Microbiol. 2015, 46, 425–432. [Google Scholar] [CrossRef]
- Nie, Z.J.; Hang, B.J.; Cai, S.; Xie, X.T.; He, J.; Li, S.P. Degradation of cyhalofop-butyl (CyB) by Pseudomonas azotoformans strain QDZ-1 and cloning of a novel gene encoding CyB-hydrolyzing esterase. J. Agric. Food Chem. 2011, 59, 6040–6046. [Google Scholar] [CrossRef]
- Huang, L.; Meng, D.; Tian, Q.; Yang, S.; Deng, H.; Guan, Z.; Cai, Y.; Liao, X. Characterization of a novel carboxylesterase from Bacillus velezensis SYBC H47 and its application in degradation of phthalate esters. J. Biosci. Bioeng. 2020, 129, 588–594. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Yang, Y.; Yue, D.; Xiao, L.; Yang, L. Biodegradation of an endocrine-disrupting chemical di-n-butyl phthalate by newly isolated Camelimonas sp. and enzymatic properties of its hydrolase. Biodegradation 2015, 26, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Zheng, S.W.; Shen, Y.J.; Zheng, G.D.; Liu, H.T.; Wu, Z.Y. Complete genome sequence provides insights into the biodrying-related microbial function of Bacillus thermoamylovorans isolated from sewage sludge biodrying material. Bioresour. Technol. 2018, 260, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57 Pt 1, 81–91. [Google Scholar] [CrossRef]
- Varghese, N.J.; Mukherjee, S.; Ivanova, N.; Konstantinidis, K.T.; Mavrommatis, K.; Kyrpides, N.C.; Pati, A. Microbial species delineation using whole genome sequences. Nucleic Acids Res. 2015, 43, 6761–6771. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Meyers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, J.; Ren, C.; Nahurira, R.; Khokhar, I.; Wang, J.; Fan, S.; Yan, Y. Identification and characterization of a meta-cleavage product hydrolase involved in biphenyl degradation from Arthrobacter sp. YC-RL1. Appl. Microbiol. Biotechnol. 2019, 103, 6825–6836. [Google Scholar] [CrossRef]
- Ren, L.; Jia, Y.; Zhang, R.; Lin, Z.; Zhen, Z.; Hu, H.; Yan, Y. Insight into Metabolic Versatility of an Aromatic Compounds-Degrading Arthrobacter sp. YC-RL1. Front. Microbiol. 2018, 9, 2438. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway ID | Description | Gene No. |
|---|---|---|
| ko00982 | Drug metabolism—cytochrome P450 | 13 |
| ko00980 | Metabolism of xenobiotics by cytochrome P450 | 12 |
| ko00791 | Atrazine degradation | 10 |
| ko00362 | Benzoate degradation | 8 |
| ko00930 | Caprolactam degradation | 8 |
| ko00627 | Aminobenzoate degradation | 8 |
| ko00625 | Chloroalkane and chloroalkene degradation | 7 |
| ko00643 | Styrene degradation | 7 |
| ko00983 | Drug metabolism—other enzymes | 7 |
| ko00361 | Chlorocyclohexane and chlorobenzene degradation | 4 |
| ko00633 | Nitrotoluene degradation | 3 |
| ko00621 | Dioxin degradation | 2 |
| ko00626 | Naphthalene degradation | 2 |
| ko00622 | Xylene degradation | 2 |
| ko00364 | Fluorobenzoate degradation | 1 |
| ko00624 | Polycyclic aromatic hydrocarbon degradation | 1 |
| ko00623 | Toluene degradation | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, J.; Jia, Y.; Reheman, A.; Yan, Y. The Genome Analysis of Methylobacterium populi YC-XJ1 with Diverse Xenobiotics Biodegrading Capacity and Degradation Characteristics of Related Hydrolase. Int. J. Mol. Sci. 2020, 21, 4436. https://doi.org/10.3390/ijms21124436
Li X, Wang J, Jia Y, Reheman A, Yan Y. The Genome Analysis of Methylobacterium populi YC-XJ1 with Diverse Xenobiotics Biodegrading Capacity and Degradation Characteristics of Related Hydrolase. International Journal of Molecular Sciences. 2020; 21(12):4436. https://doi.org/10.3390/ijms21124436
Chicago/Turabian StyleLi, Xianjun, Junhuan Wang, Yang Jia, Aikebaier Reheman, and Yanchun Yan. 2020. "The Genome Analysis of Methylobacterium populi YC-XJ1 with Diverse Xenobiotics Biodegrading Capacity and Degradation Characteristics of Related Hydrolase" International Journal of Molecular Sciences 21, no. 12: 4436. https://doi.org/10.3390/ijms21124436
APA StyleLi, X., Wang, J., Jia, Y., Reheman, A., & Yan, Y. (2020). The Genome Analysis of Methylobacterium populi YC-XJ1 with Diverse Xenobiotics Biodegrading Capacity and Degradation Characteristics of Related Hydrolase. International Journal of Molecular Sciences, 21(12), 4436. https://doi.org/10.3390/ijms21124436