The Toxicological Aspects of the Heat-Borne Toxicant 5-Hydroxymethylfurfural in Animals: A Review


 , ,
, ,  ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
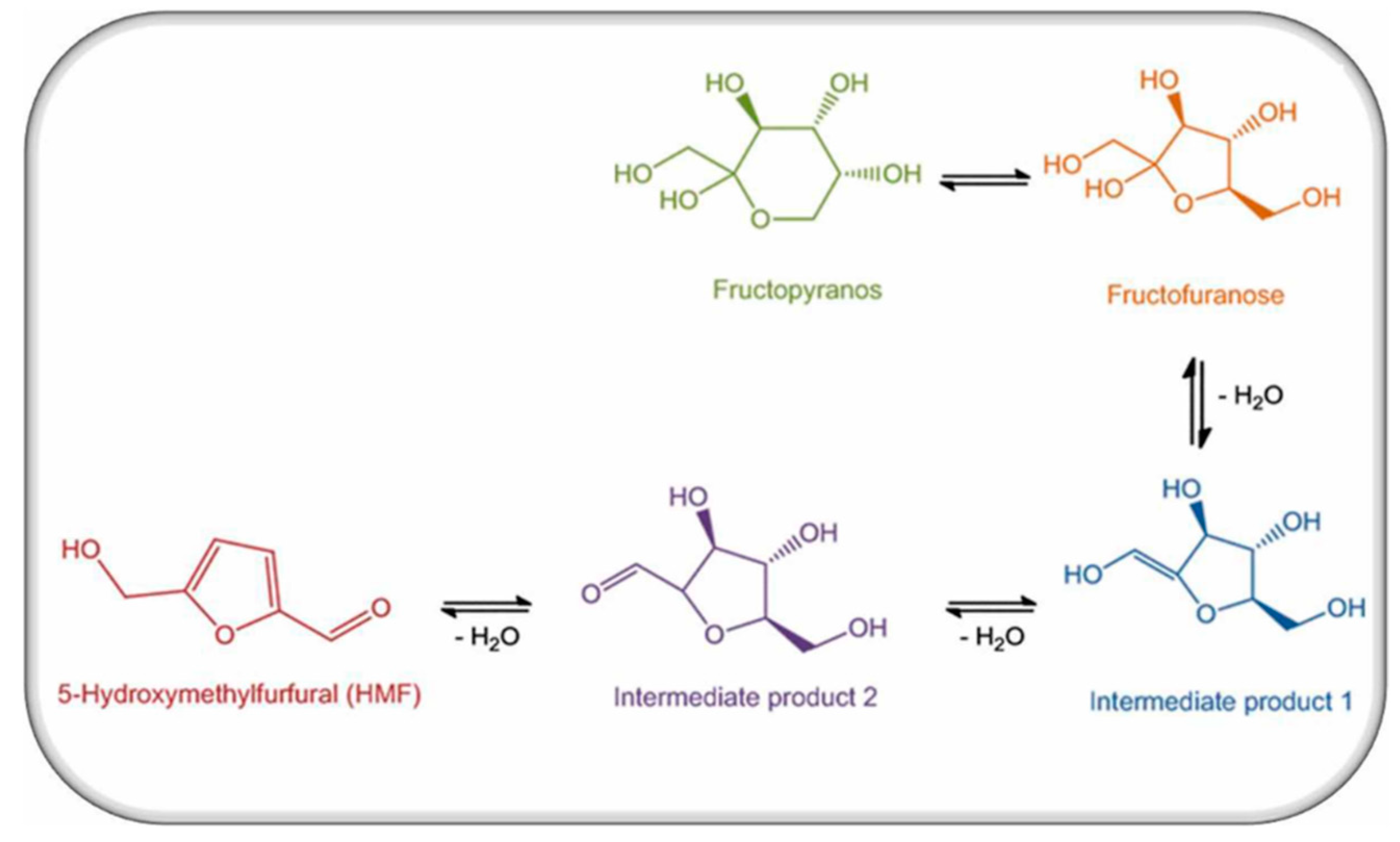
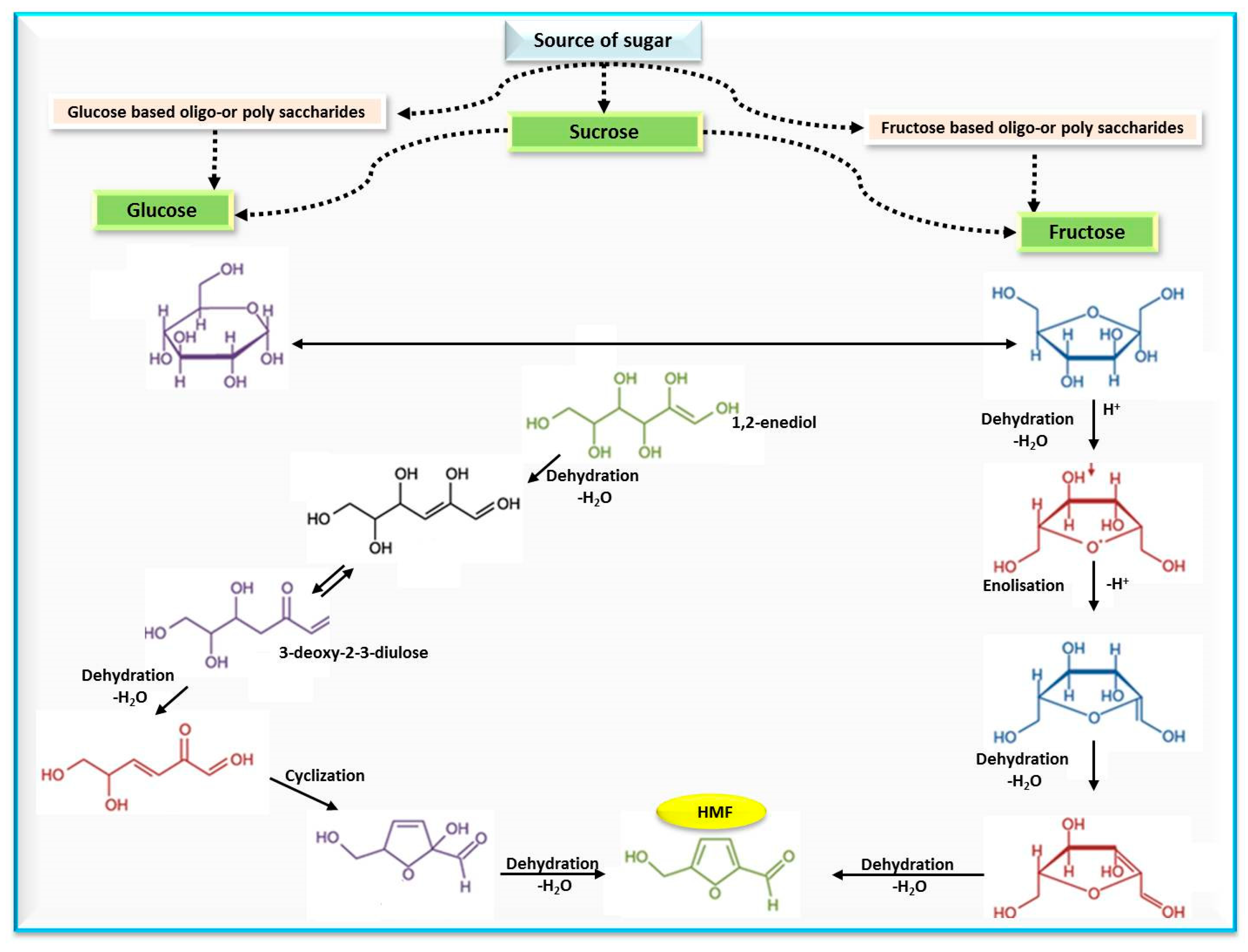
2. Definition of 5-Hydroxymethylfurfural (HMF)
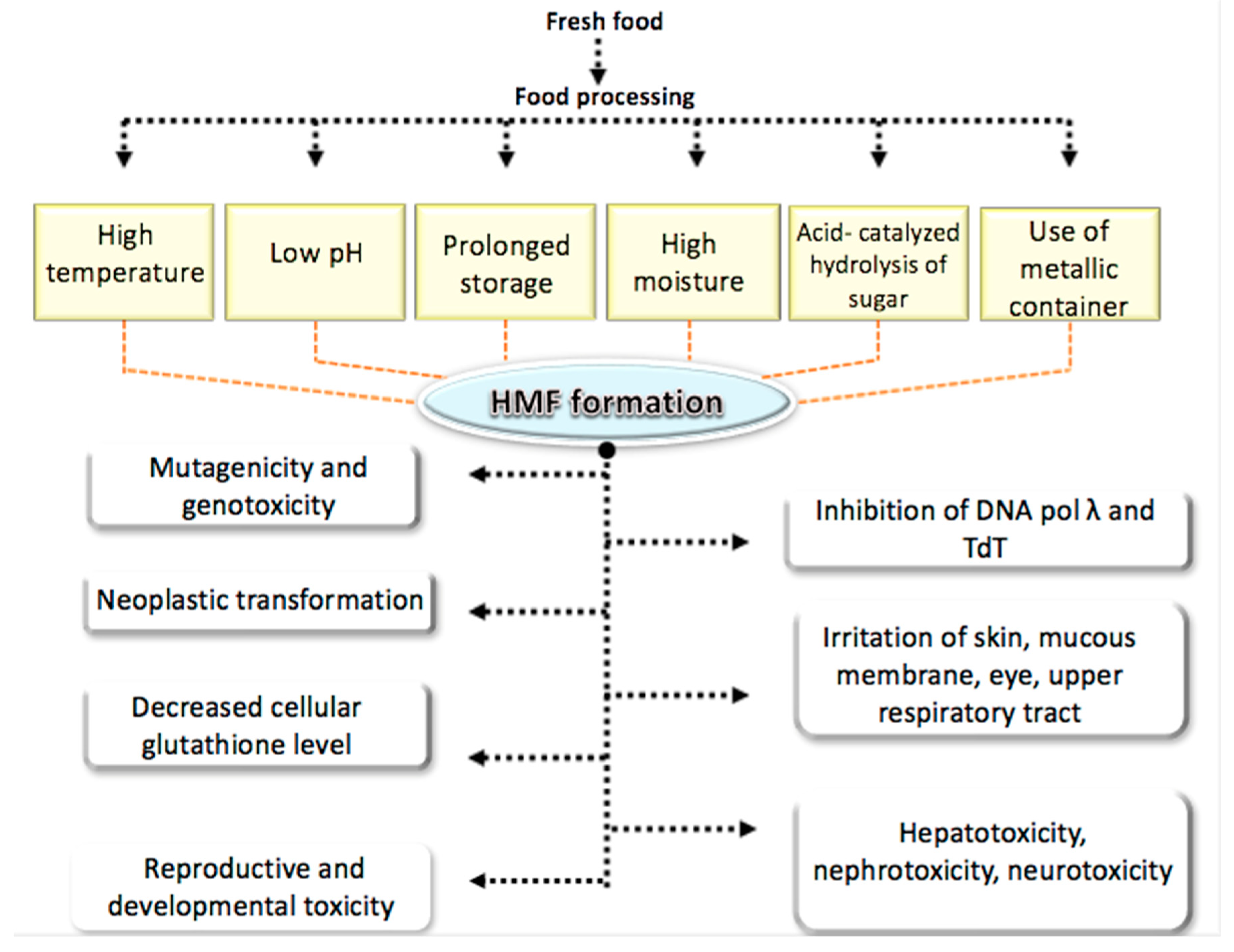
3. Occurrence and Dietary Exposure to HMF
4. Acceptable Daily Intake of HMF
5. Uses of HMF
6. Metabolism of HMF
7. Toxicity of HMF
7.1. Hepatotoxic Effects
7.2. Nephrotoxic Effect
7.3. Neurotoxic Effect
7.4. Reproductive and Developmental Toxicity
7.5. Genotoxic and Mutagenic Effect
7.6. Cytotoxicity Effect
7.7. Inhibition of Enzymes Effect
8. Prooxidant Effects of HMF
9. Carcinogenic Effects of HMF
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Van Boekel, M.; Fogliano, V.; Pellegrini, N.; Stanton, C.; Scholz, G.; Lalljie, S.; Somoza, V.; Knorr, D.; Jasti, P.R.; Eisenbrand, G. A review on the beneficial aspects of food processing. Mol. Nutr. Food Res. 2010, 54, 1215–1247. [Google Scholar] [CrossRef] [PubMed]
- Rada-Mendoza, M.; Sanz, M.; Olano, A.; Villamiel, M. Formation of hydroxymethylfurfural and furosine during the storage of jams and fruit-based infant foods. Food Chem. 2004, 85, 605–609. [Google Scholar] [CrossRef]
- Teixidó, E.; Santos, F.; Puignou, L.; Galceran, M. Analysis of 5-hydroxymethylfurfural in foods by gas chromatography–mass spectrometry. J. Chromatogr. A 2006, 1135, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Staszak-Kozinski, L.; Davidson, R. Up in smoke: I. Smoke-derived germination cues for postfire annual, Nicotiana attenuata torr. Ex. Watson. J. Chem. Ecol. 1994, 20, 2345–2371. [Google Scholar] [CrossRef]
- Chou, C.C.; Hee, S.S.Q. Bioassay-driven analysis of chewing tobacco extracts. Environ. Toxicol. Chem. Int. J. 1994, 13, 1177–1186. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Tannenbaum, S.R. Activation of the Maillard Reaction Product 5-(Hydroxymethyl)furfural to Strong Mutagens via Allylic Sulfonation and Chlorination. Chem. Res. Toxicol. 1994, 7, 313–318. [Google Scholar] [CrossRef]
- Bakhiya, N.; Monien, B.; Frank, H.; Seidel, A.; Glatt, H. Renal organic anion transporters OAT1 and OAT3 mediate the cellular accumulation of 5-sulfooxymethylfurfural, a reactive, nephrotoxic metabolite of the Maillard product 5-hydroxymethylfurfural. Biochem. Pharmacol. 2009, 78, 414–419. [Google Scholar] [CrossRef] [Green Version]
- Durling, L.J.; Busk, L.; Hellman, B.E. Evaluation of the DNA damaging effect of the heat-induced food toxicant 5-hydroxymethylfurfural (HMF) in various cell lines with different activities of sulfotransferases. Food Chem. Toxicol. 2009, 47, 880–884. [Google Scholar] [CrossRef]
- Svendsen, C.; Husøy, T.; Glatt, H.; Paulsen, J.E.; Alexander, J. 5-Hydroxymethylfurfural and 5-sulfooxymethylfurfural increase adenoma and flat ACF number in the intestine of Min/+ mice. Anticancer. Res. 2009, 29, 1921–1926. [Google Scholar]
- Høie, A.H.; Svendsen, C.; Brunborg, G.; Glatt, H.; Alexander, J.; Meinl, W.; Husøy, T. Genotoxicity of three food processing contaminants in transgenic mice expressing human sulfotransferases 1A1 and 1A2 as assessed by the in vivo alkaline single cell gel electrophoresis assay. Environ. Mol. Mutagen. 2015, 56, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Husøy, T.; Haugen, M.; Murkovic, M.; Jöbstl, D.; Stølen, L.; Bjellaas, T.; Rønningborg, C.; Glatt, H.; Alexander, J. Dietary exposure to 5-hydroxymethylfurfural from Norwegian food and correlations with urine metabolites of short-term exposure. Food Chem. Toxicol. 2008, 46, 3697–3702. [Google Scholar] [CrossRef] [PubMed]
- Svensson, K.; Abramsson, L.; Becker, W.; Glynn, A.; Hellenäs, K.-E.; Lind, Y.; Rosén, J. Dietary intake of acrylamide in Sweden. Food Chem. Toxicol. 2003, 41, 1581–1586. [Google Scholar] [CrossRef]
- Morehouse, K.M.; Nyman, P.J.; McNeal, T.P.; DiNovi, M.J.; Perfetti, G.A. Survey of furan in heat processed foods by headspace gas chromatography/mass spectrometry and estimated adult exposure. Food Addit. Contam. Part A 2008, 25, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Sachse, B.; Meinl, W.; Sommer, Y.; Glatt, H.; Seidel, A.; Monien, B.H. Bioactivation of food genotoxicants 5-hydroxymethylfurfural and furfuryl alcohol by sulfotransferases from human, mouse and rat: A comparative study. Arch. Toxicol. 2014, 90, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glatt, H.R. Health risks by 5-hydroxymethyl-furfural (HMF) and related compounds. Acrylamide other heal. Hazard. Compd. Heat-treated Foods 2006, 2, 328–353. [Google Scholar]
- Lee, Y.; Shlyankevich, M.; Jeong, H.; Douglas, J.; Surh, Y. Bioactivation of 5-Hydroxymethyl-2-Furaldehyde to an Electrophilic and Mutagenic Allylic Sulfuric Acid Ester. Biochem. Biophys. Res. Commun. 1995, 209, 996–1002. [Google Scholar] [CrossRef]
- Glatt, H.; Schneider, H.; Liu, Y. V79-hCYP2E1-hSULT1A1, a cell line for the sensitive detection of genotoxic effects induced by carbohydrate pyrolysis products and other food-borne chemicals. Mutat. Res. Toxicol. Environ. Mutagen. 2005, 580, 41–52. [Google Scholar] [CrossRef]
- Ames, J.M. The Maillard reaction. In Biochemistry of food proteins; Springer: Whiteknights, UK, 1992; pp. 99–153. [Google Scholar]
- Biluca, F.C.; Della Betta, F.; De Oliveira, G.P.; Pereira, L.M.; Gonzaga, L.V.; Costa, A.C.O.; Fett, R. 5-HMF and carbohydrates content in stingless bee honey by CE before and after thermal treatment. Food Chem. 2014, 159, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Nagy, S. Relative Reactivities of Sugars in the Formation of 5-Hydroxymethylfurfural in Sugar-Catalyst Model Systems. J. Food Process. Preserv. 1990, 14, 171–178. [Google Scholar] [CrossRef]
- Locas, C.P.; Yaylayan, V.A. Isotope Labeling Studies on the Formation of 5-(Hydroxymethyl)-2-furaldehyde (HMF) from Sucrose by Pyrolysis-GC/MS. J. Agric. Food Chem. 2008, 56, 6717–6723. [Google Scholar] [CrossRef] [PubMed]
- Akkan, A.A.; Özdemir, Y.; Ekiz, H.L. Derivative spectrophotometric determination of 5-(hydroxymethyl)-2-furaldehyde (HMF) and furfural in Locust bean extract. Food/Nahrung 2001, 45, 43–46. [Google Scholar] [CrossRef]
- Polovková, M.; Šimko, P. Determination and occurrence of 5-hydroxymethyl-2-furaldehyde in white and brown sugar by high performance liquid chromatography. Food Control. 2017, 78, 183–186. [Google Scholar] [CrossRef]
- Kowalski, S.; Łukasiewicz, M.; Duda-Chodak, A.; Zięć, G. 5-Hydroxymethyl-2-Furfural (HMF) – Heat-Induced Formation, Occurrence in Food and Biotransformation - a Review. Pol. J. Food Nutr. Sci. 2013, 63, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Mańkowska, D.; Majak, I.; Bartos, A.; Słowianek, M.; Łącka, A.; Leszczyńska, J. 5-hydroxymethylfurfural content in selected gluten- and gluten-free cereal food products. Biotechnol. Food Sci. 2017, 81, 11–21. [Google Scholar]
- Henares, J.; Ángel, R.; Garcia-Villanova, B.; Guerra-Hernandez, E.J. Occurrence of furosine and hydroxymethylfurfural as markers of thermal damage in dehydrated vegetables. Eur. Food Res. Technol. 2008, 228, 249–256. [Google Scholar] [CrossRef]
- Henares, J.; Ángel, R.; Delgado-Andrade, C.; Morales, F.J. Analysis of heat-damage indices in breakfast cereals: Influence of composition. J. Cereal Sci. 2006, 43, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Jiménez, A.; Guerra-Hernández, E.; García-Villanova, B. Browning Indicators in Bread. J. Agric. Food Chem. 2000, 48, 4176–4181. [Google Scholar] [CrossRef]
- Serpen, A.; Gökmen, V.; Mogol, B.A. Effects of different grain mixtures on Maillard reaction products and total antioxidant capacities of breads. J. Food Compos. Anal. 2012, 26, 160–168. [Google Scholar] [CrossRef]
- Mortas, M.; Gul, O.; Yazici, F.; Dervisoğlu, M. Effect of brewing process and sugar content on 5-hydroxymethylfurfural and related substances from Turkish coffee. Int. J. Food Prop. 2016, 20, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Arribas-Lorenzo, G.; Morales, F.J. Estimation of dietary intake of 5-hydroxymethylfurfural and related substances from coffee to Spanish population. Food Chem. Toxicol. 2010, 48, 644–649. [Google Scholar] [CrossRef]
- Shapla, U.M.; Solayman, M.; Alam, N.; Khalil, I.; Gan, S.H. 5-Hydroxymethylfurfural (HMF) levels in honey and other food products: Effects on bees and human health. Chem. Central J. 2018, 12, 35. [Google Scholar] [CrossRef]
- Erbersdobler, H.F.; Hupe, A. Determination of lysine damage and calculation of lysine bio-availability in several processed foods. Eur. J. Nutr. 1991, 30, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Albalá-Hurtado, S.; Veciana-Nogués, M.T.; Mariné-Font, A.; Vidal-Carou, M.C. Changes in Furfural Compounds during Storage of Infant Milks. J. Agric. Food Chem. 1998, 46, 2998–3003. [Google Scholar] [CrossRef]
- Cais-Sokolińska, D.; Pikul, J.; Danków, R. Measurement of colour parameters as index of the hidroxymethylfurfural content in the UHT sterilised milk during its storage. Electron. J. Pol. Agric. Univ. 2004, 7, 1–3. [Google Scholar]
- Alston, T.A.; Mela, L.; Bright, H.J. 3-Nitropropionate, the toxic substance of Indigofera, is a suicide inactivator of succinate dehydrogenase. Proc. Natl. Acad. Sci. 1977, 74, 3767–3771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, S.; Zhang, Q. Effects of Commercial-scale Pulsed Electric Field Processing on Flavor and Color of Tomato Juice. J. Food Sci. 2003, 68, 1600–1606. [Google Scholar] [CrossRef]
- Murkovic, M.; Pichler, N. Analysis of 5-hydroxymethylfurfual in coffee, dried fruits and urine. Mol. Nutr. Food Res. 2006, 50, 842–846. [Google Scholar] [CrossRef]
- Fallico, B.; Arena, E.; Zappalà, M. Roasting of hazelnuts. Role of oil in colour development and hydroxymethylfurfural formation. Food Chem. 2003, 81, 569–573. [Google Scholar] [CrossRef]
- Monakhova, Y.B.; Lachenmeier, D.W. The Margin of Exposure of 5-Hydroxymethylfurfural (HMF) in Alcoholic Beverages. Environ. Heal. Toxicol. 2012, 27, e2012016. [Google Scholar] [CrossRef]
- Appel, K.E.; Gürtler, R.; Berg, K.; Heinemeyer, G.; Lampen, A.; Appel, K.E. Toxicology and risk assessment of 5-Hydroxymethylfurfural in food. Mol. Nutr. Food Res. 2011, 55, 667–678. [Google Scholar] [CrossRef]
- Henares, J.; Ángel, R.; De La Cueva, S.P. Assessment of hydroxymethylfurfural intake in the Spanish diet. Food Addit. Contam. Part A 2008, 25, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Janzowski, C.; Glaab, V.; Samimi, E.; Schlatter, J.; Eisenbrand, G. 5-Hydroxymethylfurfural: Assessment of mutagenicity, DNA-damaging potential and reactivity towards cellular glutathione. Food Chem. Toxicol. 2000, 38, 801–809. [Google Scholar] [CrossRef]
- Ulbricht, R.J.; Northup, S.J.; Thomas, J.A. A Review of 5-Hydroxymethylfurfural (HMF) in Parenteral Solutions. Toxicol. Sci. 1984, 4, 843–853. [Google Scholar] [CrossRef]
- Zaitzev, A.N.; Simonian, T.A.; Pozdniakov, A.L. Hygienic standards for hydroxymethylfurfural in food products. Vopr Pitan 1975, 1, 52–55. [Google Scholar]
- De La Cueva, S.P.; Álvarez, J.; Vegvari, A.; Montilla-Gómez, J.; Cruz-López, O.; Delgado-Andrade, C.; Henares, J. Ángel R. Relationship between HMF intake and SMF formation in vivo: An animal and human study. Mol. Nutr. Food Res. 2016, 61, 1600773. [Google Scholar] [CrossRef]
- Dauberte, B.; Estienne, J.; Guerere, M. Hydroxymethylfurfural formation in fruit juice drinks and torrefacto coffees. Ann. des Falsif. l’Expertise Chim. Toxicol. 1990, 83, 231–253. [Google Scholar]
- Fallico, B.; Zappala, M.; Arena, E.; Verzera, A. Effects of conditioning on HMF content in unifloral honeys. Food Chem. 2004, 85, 305–313. [Google Scholar] [CrossRef]
- Garcia-Villanova, B.; Guerra-Hernandez, E.J.; Martinez-Gomez, E.; Montilla, J. Liquid chromatography for the determination of 5-(hydroxymethyl)-2-furaldehyde in breakfast cereals. J. Agric. Food Chem. 1993, 41, 1254–1255. [Google Scholar] [CrossRef]
- Resmini, P.; Pagani, M.A.; De Noni, I.; Pellegrino, L. Formation of 2-acetyl-3-d-glucopy-ranosilfuran (glucosylisomatol) from nonenzymatic browning in pasta drying. Ital. J. Food Sci. 1993, 5, 341–353. [Google Scholar]
- Fernández-Artigas, P.; Guerra-Hernandez, E.J.; García-Villanova, B. Browning indicators in model systems and baby cereals. J. Agric. Food Chem. 1999, 47, 2872–2878. [Google Scholar] [CrossRef]
- Ramirez-Jiménez, A.; Guerra-Hernandez, E.J.; Garcia-Villanova, B. Evolution of non-enzymatic browning during storage of infant rice cereal. Food Chem. 2003, 83, 219–225. [Google Scholar] [CrossRef]
- Pérez-Burillo, S.; Henares, J.; Ángel, R.; Pastoriza, S. Effect of home cooking on the antioxidant capacity of vegetables: Relationship with Maillard reaction indicators. Food Res. Int. 2018, 121, 514–523. [Google Scholar] [CrossRef]
- Godfrey, L.-J.C.V.B. DISTRIBUTION AND METABOLISM OF (5-HYDROXYMETHYL)FURFURAL IN MALE F344 RATS AND B6C3F1 MICE AFTER ORAL ADMINISTRATION. J. Toxicol. Environ. Heal. Part A 1999, 57, 199–210. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Seiquer, I.; Navarro, M.P.; Morales, F.J. Estimation of hydroxymethylfurfural availability in breakfast cereals. Studies in Caco-2 cells. Food Chem. Toxicol. 2008, 46, 1600–1607. [Google Scholar] [CrossRef]
- Germond, J.; Philippossian, G.; Richli, U.; Bracco, I.; Arnaud, M.J. Rapid and complete urinary elimination of [ 14 C]?5?hydroxymethyl?2?furaldehyde administered orally or intravenously to rats. J. Toxicol. Environ. Heal. Part A 1987, 22, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Monien, B.H.; Frank, H.; Seidel, A.; Glatt, H. Conversion of the Common Food Constituent 5-Hydroxymethylfurfural into a Mutagenic and Carcinogenic Sulfuric Acid Ester in the Mouse in Vivo. Chem. Res. Toxicol. 2009, 22, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Bauer-Marinovic, M.; Taugner, F.; Florian, S.; Glatt, H. Toxicity studies with 5-hydroxymethylfurfural and its metabolite 5-sulphooxymethylfurfural in wild-type mice and transgenic mice expressing human sulphotransferases 1A1 and 1A2. Arch. Toxicol. 2012, 86, 701–711. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program. NTP toxicology and carcinogenesis studies of 5-(Hydroxymethyl)-2-furfural (CAS No. 67-47-0) in F344/N rats and B6C3F1 mice (gavage studies). Natl. Toxicol. Program Tech. Rep. Ser. 2010, 554, 1–180. [Google Scholar]
- Zhao, L.; Chen, J.; Su, J.; Li, L.; Hu, S.-Q.; Li, B.; Zhang, X.; Xu, Z.; Chen, T. In VitroAntioxidant and Antiproliferative Activities of 5-Hydroxymethylfurfural. J. Agric. Food Chem. 2013, 61, 10604–10611. [Google Scholar] [CrossRef]
- Gu, H.; Jiang, Y.-B.; Jiang, H.-Y.; Xu, N.-Q.; Yu, J.-T.; Ding, X.; Zhao, F.-M.; Zhan, Z.; Wang, M.-Y. Effect of 5-hydroxymethyl furfural on BCL-2 and NF-kappaB gene expression of apoptotic rat hippocampal neurons injured by H2O2. Zhong yao cai = Zhongyaocai = J. Chin. Med. Mater. 2011, 34, 1753–1756. [Google Scholar]
- Elmaogullari, S.; Kadan, E.; Anadol, E.; Gokceoglu, A.; Cetinkaya, S.; Yarim, G.F.; Ucakturk, S.A.; Aycan, Z. Effects of 5-Hydroxymethylfurfural on Pubertal Development of Female Wistar Rats. J. Clin. Res. Pediatr. Endocrinol. 2020, 12, 79–85. [Google Scholar] [CrossRef]
- Severin, I.; Dumont, C.; Jondeau-Cabaton, A.; Graillot, V.; Chagnon, M.-C. Genotoxic activities of the food contaminant 5-hydroxymethylfurfural using different in vitro bioassays. Toxicol. Lett. 2010, 192, 189–194. [Google Scholar] [CrossRef]
- Nishi, Y.; Miyakawa, Y.; Kato, K. Chromosome aberrations induced by pyrolysates of carbohydrates in Chinese hamster V79 cells. Mutat. Res. Lett. 1989, 227, 117–123. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Liem, A.; Miller, J.A.; Tannenbaum, S.R. 5-Sulfooxymethylfurfural as a possible ultimate mutagenic and carcinogenic metabolite of the Maillard reaction product, 5-hydroxymethylfurfural. Carcinog. 1994, 15, 2375–2377. [Google Scholar] [CrossRef] [PubMed]
- Stadler, R.H.; Lineback, D.R. Process-Induced Food Toxicants: Occurrence, Formation, Mitigation, and Health Risks; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; ISBN 0470430095. [Google Scholar]
- Hubscher, U.; Maga, G.; Spadari, S. Eukaryotic DNA Polymerases. Annu. Rev. Biochem. 2002, 71, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Diaz, M.; Bebenek, K. Multiple functions of DNA polymerases. Crit. Rev. Plant Sci. 2007, 26, 105–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushina, Y.; Yagita, E.; Kuramochi, K.; Kuriyama, I.; Shimazaki, N.; Koiwai, O.; Uchiyama, Y.; Yomezawa, Y.; Sugawara, F.; Kobayashi, S.; et al. 5-(Hydroxymethyl)-2-furfural: A selective inhibitor of DNA polymerase λ and terminal deoxynucleotidyltransferase. Arch. Biochem. Biophys. 2006, 446, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, H.; Amanuma, T.; Takashima, Y.; Yoshiji, H.; Nakae, D.; Tsutsumi, M.; Tsujiuchi, T.; Denda, A.; Konishi, Y. Possible enhancing effect of the immunosuppressive agent, 6-mercaptopurine(6-MP) on focal lesion development in cirrhotic liver induced by carbon tetrachloride but not furfural in F344 rats. Carcinog. 1992, 13, 1365–1369. [Google Scholar] [CrossRef]
- Shimizu, A.; Kanisawa, M. Experimental studies on hepatic cirrhosis and hepatocarcinogenesis. I. Production of hepatic cirrhosis by furfural administration. Acta Pathol. Jpn. 1986, 36, 1027–1038. [Google Scholar]
- Zhang, X.-M.; Chan, C.C.; Stamp, D.; Minkin, S.; C.Archer, M.; Bruce, W. Initiation and promotion of colonic aberrant crypt foci in rats by 5-hydroxymethy1-2-furaldehyde in thermolyzed sucrose. Carcinog. 1993, 14, 773–775. [Google Scholar] [CrossRef]
- Florian, S.; Bauer-Marinovic, M.; Taugner, F.; Dobbernack, G.; Monien, B.H.; Meinl, W.; Glatt, H. Study of 5-hydroxymethylfurfural and its metabolite 5-sulfooxymethylfurfural on induction of colonic aberrant crypt foci in wild-type mice and transgenic mice expressing human sulfotransferases 1A1 and 1A2. Mol. Nutr. Food Res. 2012, 56, 593–600. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farag, M.R.; Alagawany, M.; Bin-Jumah, M.; Othman, S.I.; Khafaga, A.F.; Shaheen, H.M.; Samak, D.; Shehata, A.M.; Allam, A.A.; Abd El-Hack, M.E. The Toxicological Aspects of the Heat-Borne Toxicant 5-Hydroxymethylfurfural in Animals: A Review. Molecules 2020, 25, 1941. https://doi.org/10.3390/molecules25081941
Farag MR, Alagawany M, Bin-Jumah M, Othman SI, Khafaga AF, Shaheen HM, Samak D, Shehata AM, Allam AA, Abd El-Hack ME. The Toxicological Aspects of the Heat-Borne Toxicant 5-Hydroxymethylfurfural in Animals: A Review. Molecules. 2020; 25(8):1941. https://doi.org/10.3390/molecules25081941
Chicago/Turabian StyleFarag, Mayada R., Mahmoud Alagawany, May Bin-Jumah, Sarah I. Othman, Asmaa F. Khafaga, Hazem M. Shaheen, Dalia Samak, Abdelrazeq M. Shehata, Ahmed A. Allam, and Mohamed E. Abd El-Hack. 2020. "The Toxicological Aspects of the Heat-Borne Toxicant 5-Hydroxymethylfurfural in Animals: A Review" Molecules 25, no. 8: 1941. https://doi.org/10.3390/molecules25081941