Stress-Induced Morphological, Cellular and Molecular Changes in the Brain—Lessons Learned from the Chronic Mild Stress Model of Depression


Abstract
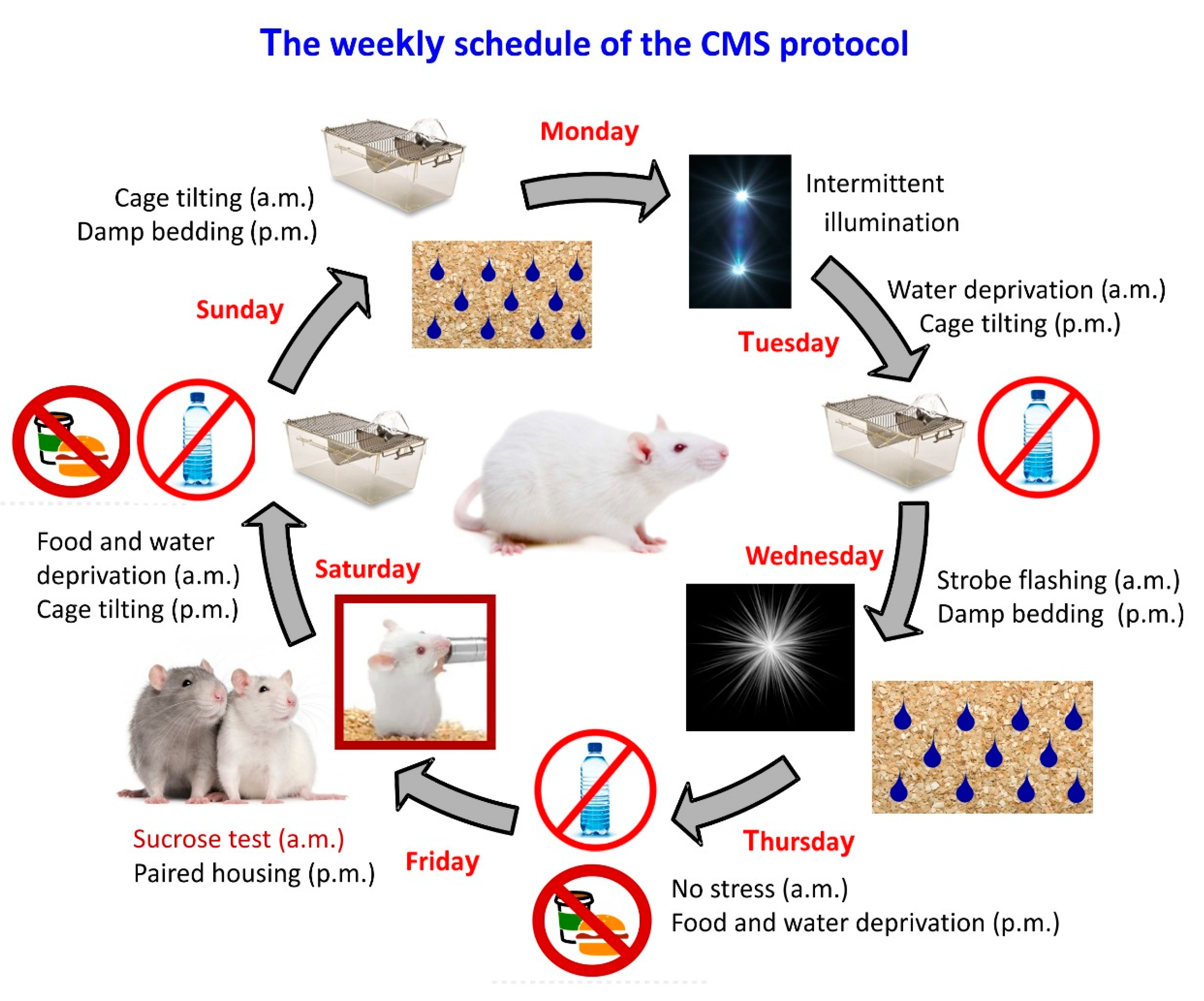
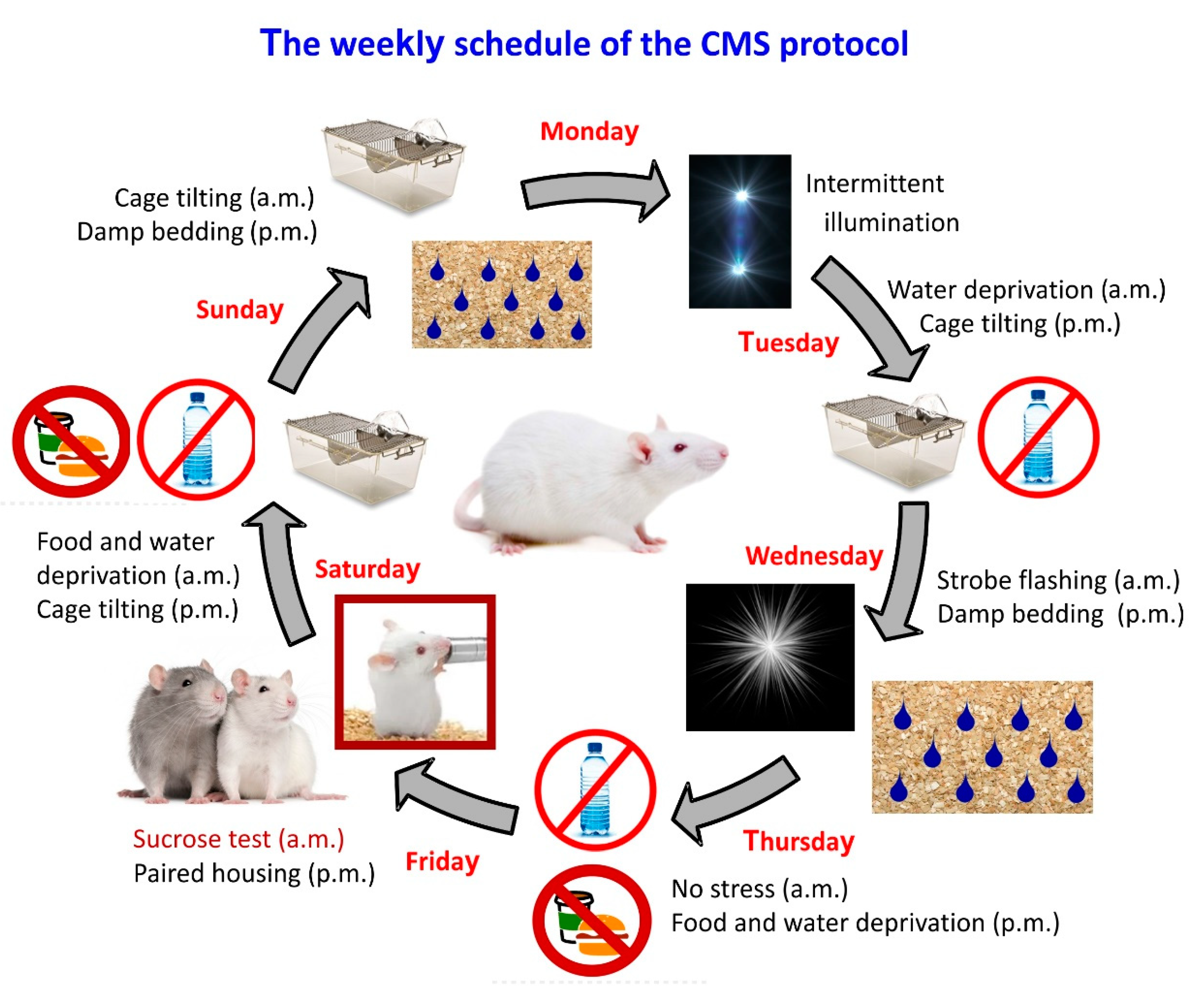
:1. Introduction
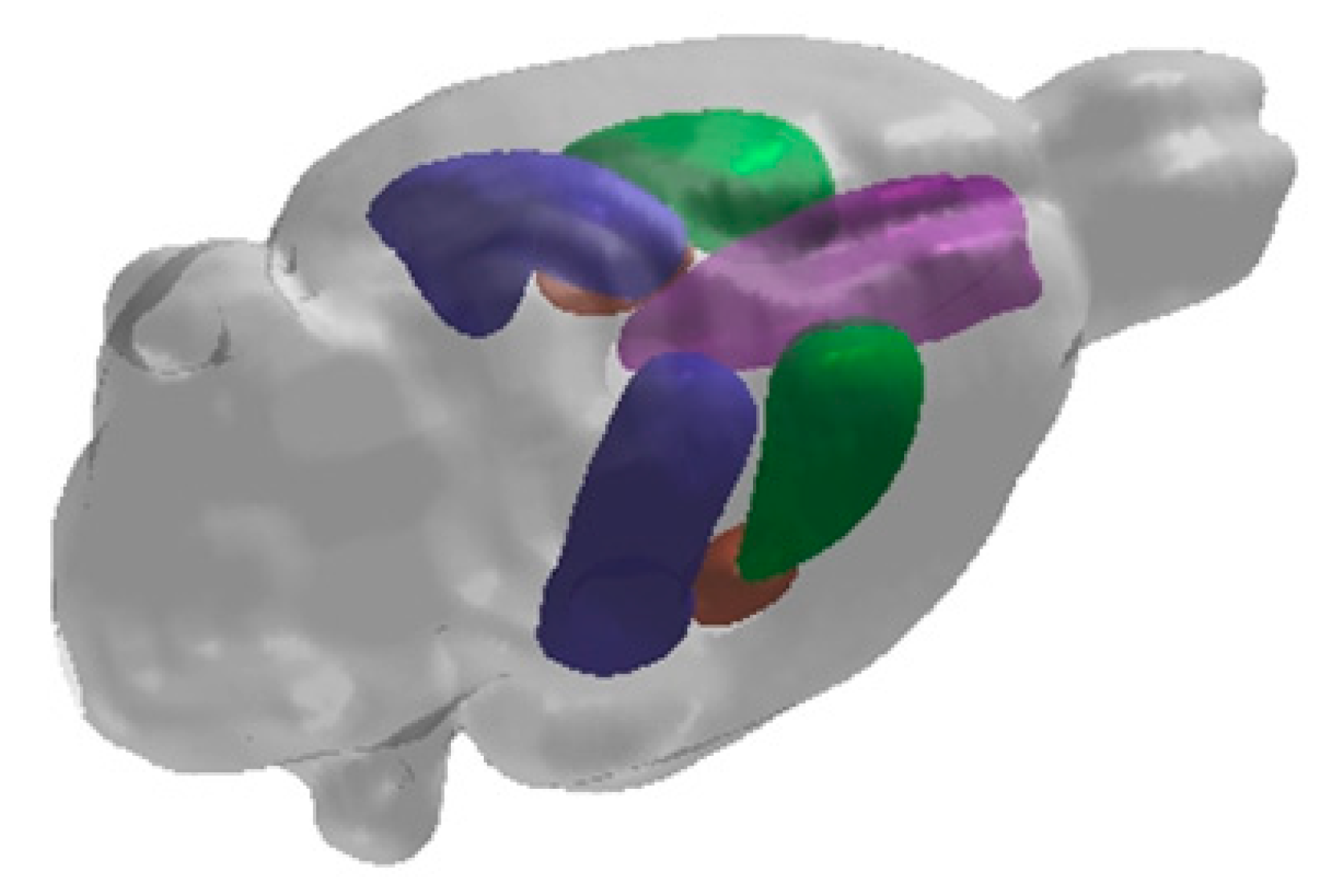
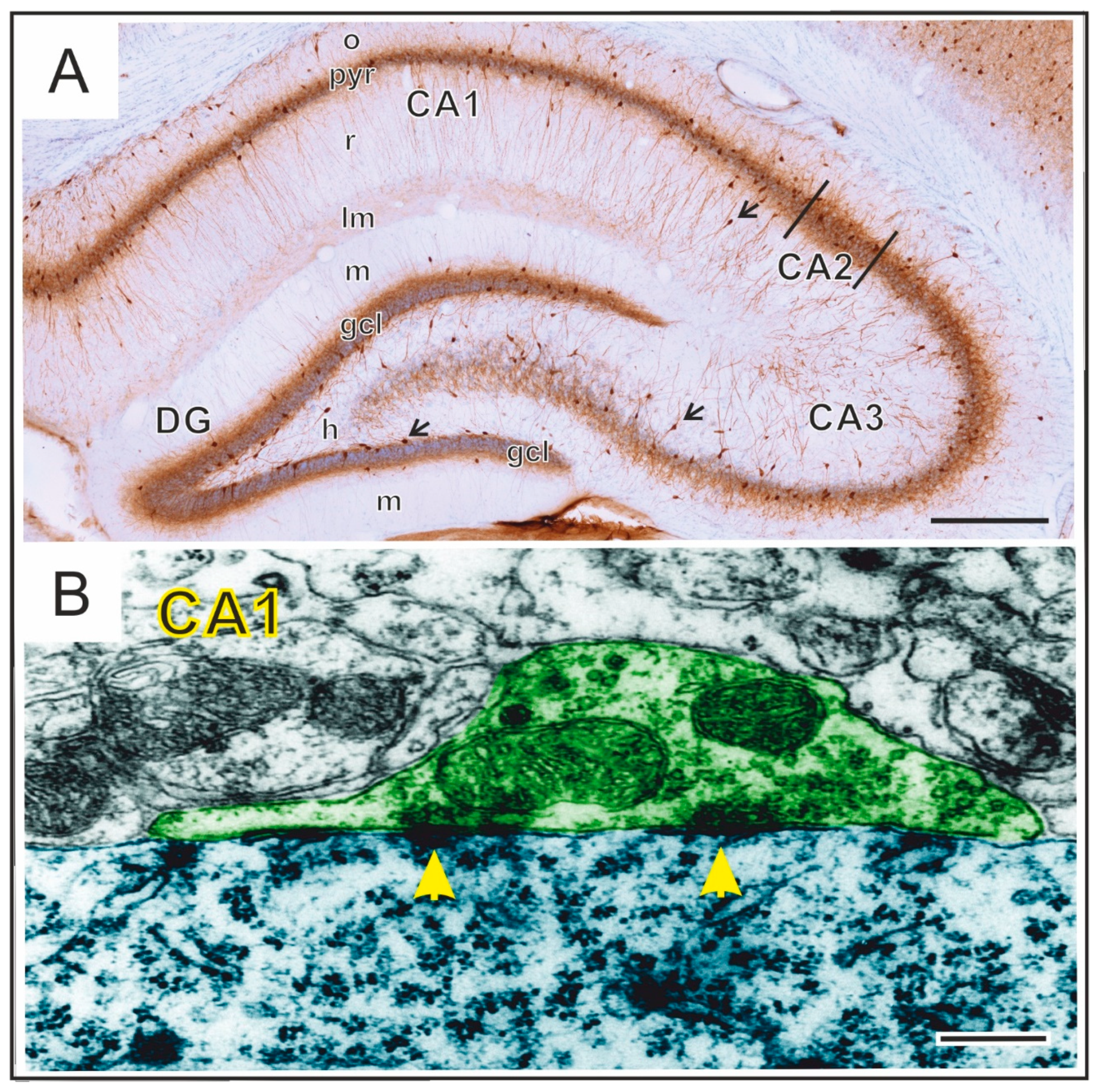
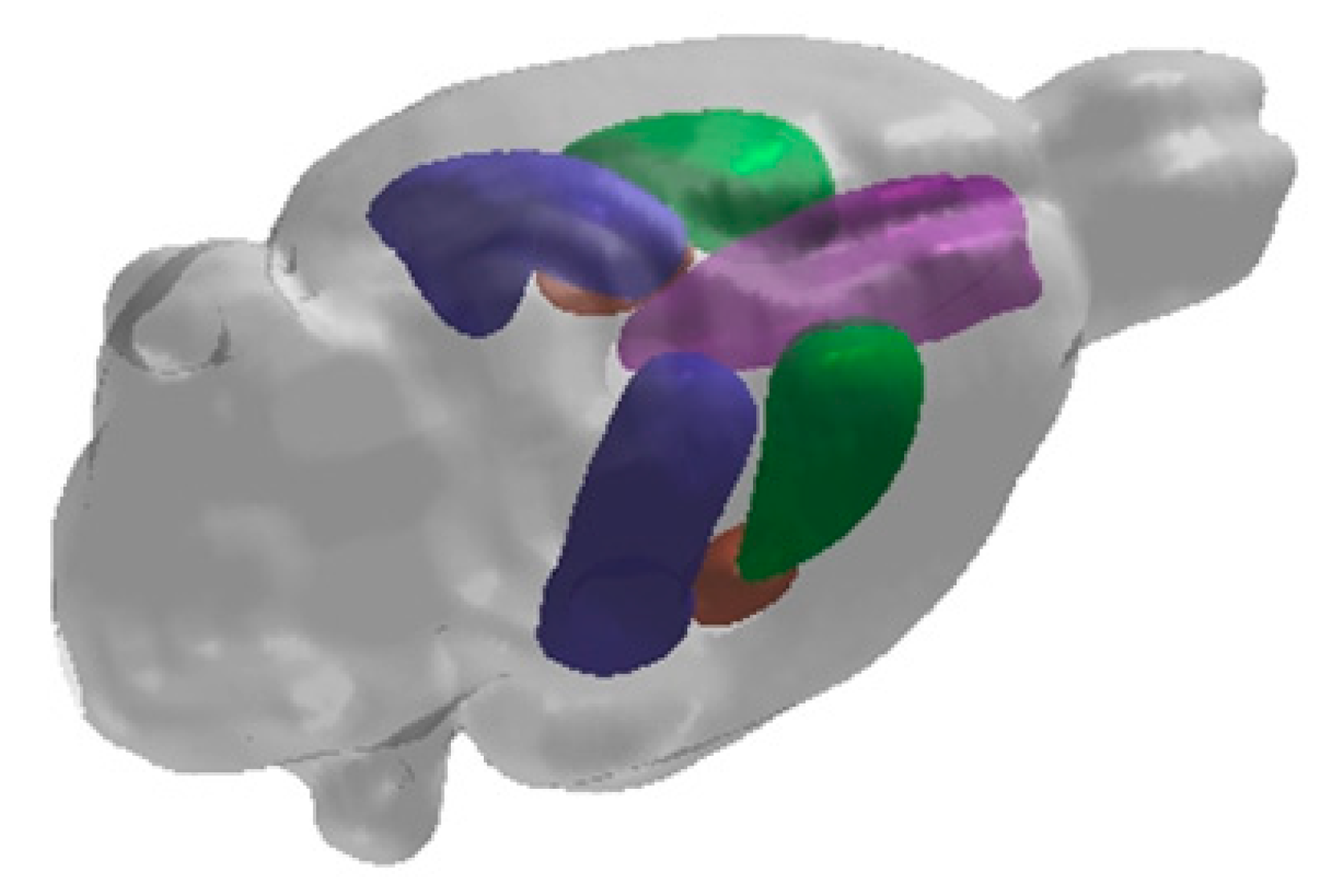
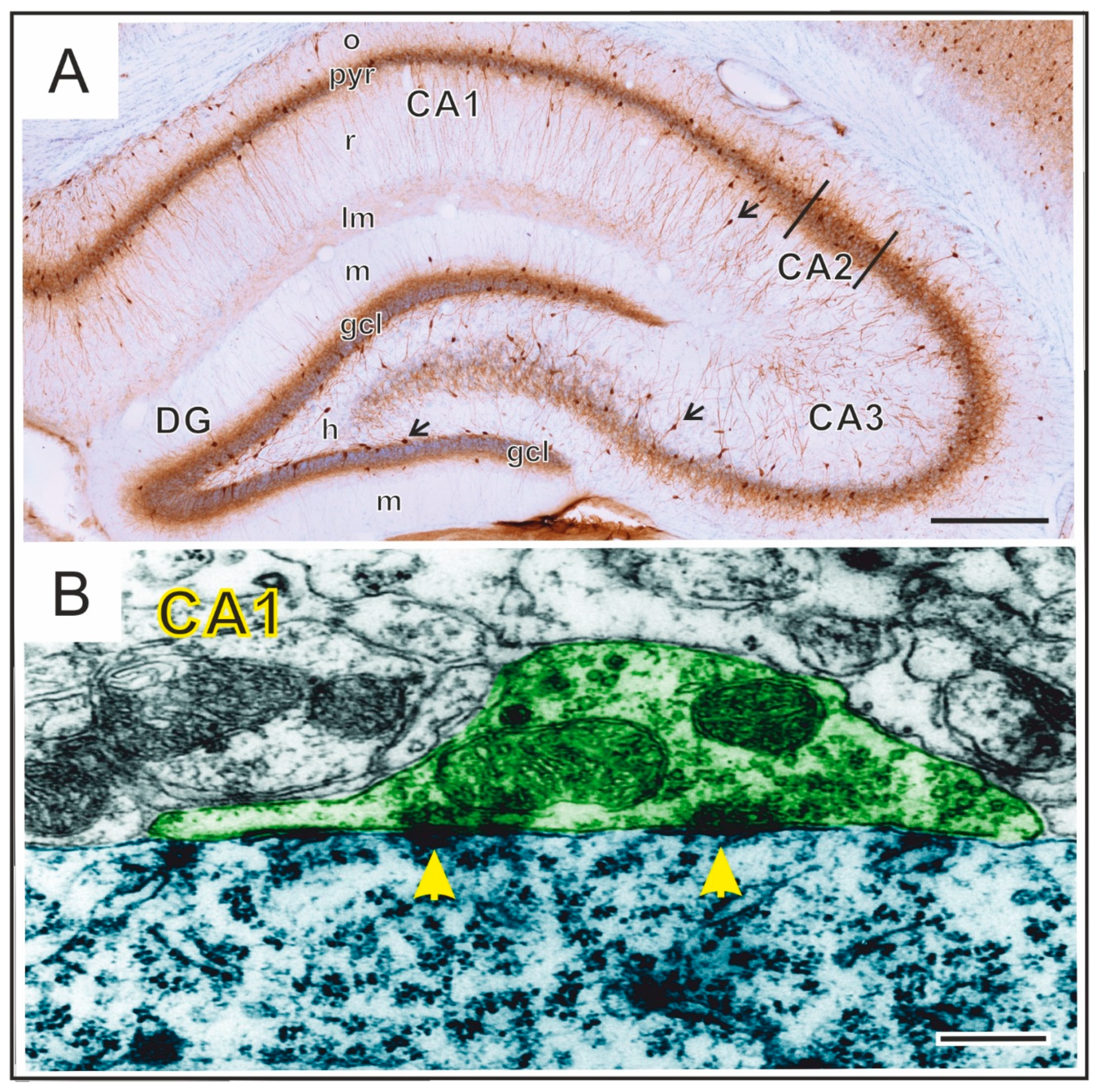
2. Hippocampal Formation
3. Prefrontal Cortex (PFC)
4. Other Neocortical Areas
5. Amygdala
6. The Dorsal Striatum
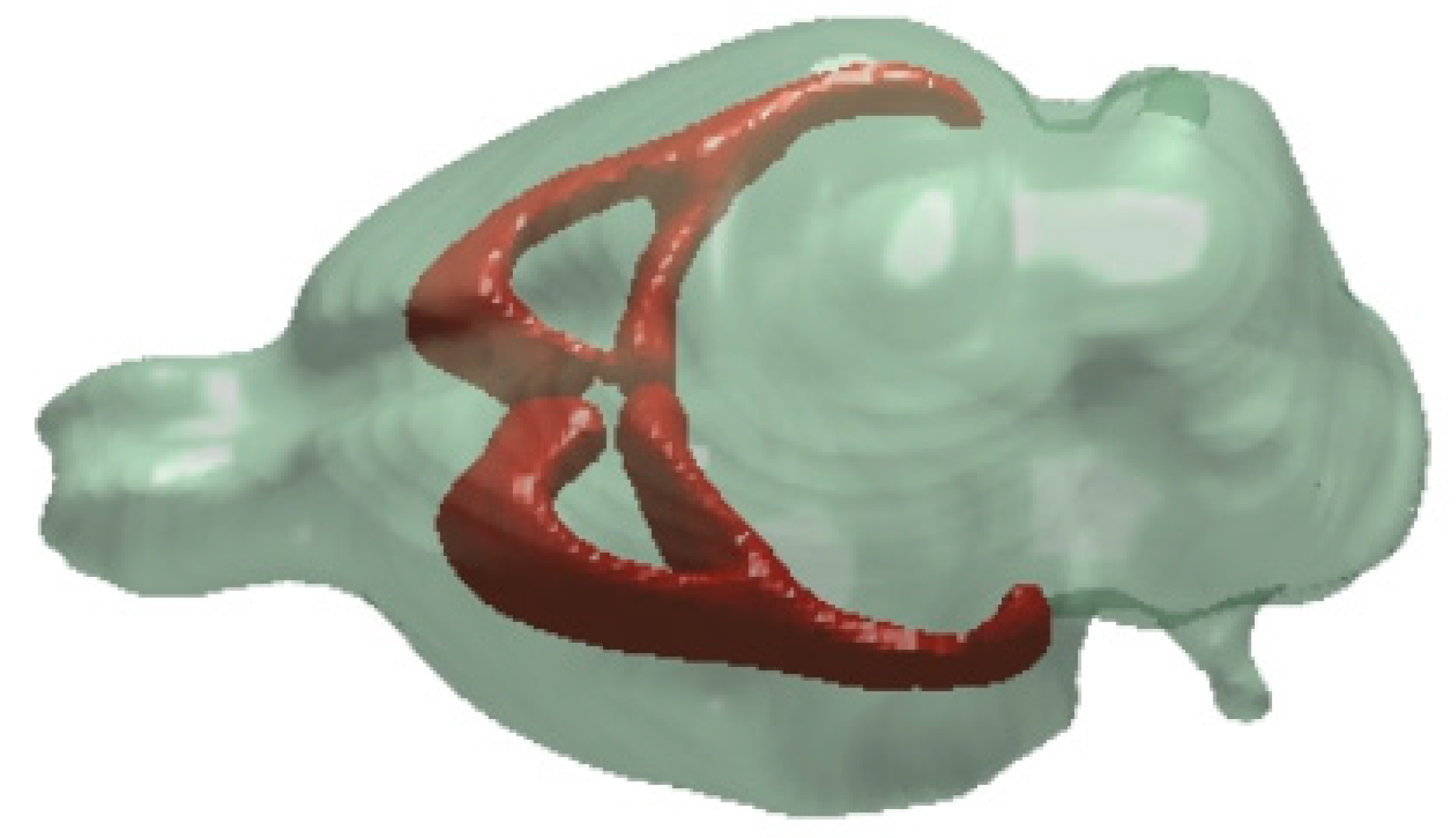
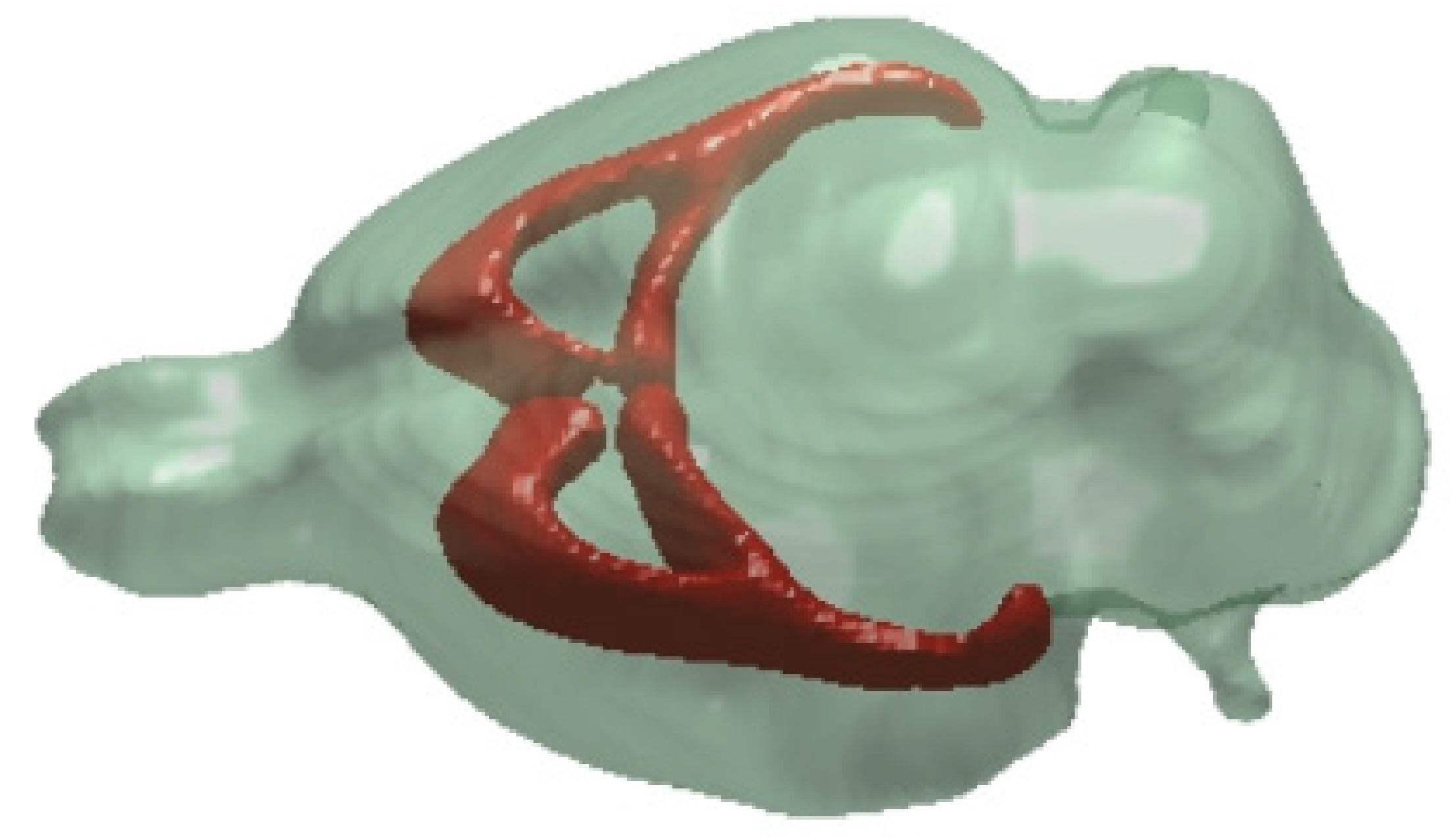
7. White Matter Regions
8. Other Brain Areas
9. MicroRNA Changes in the CMS Model
10. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Assembly. 65 Global Burden of Mental Disorders and the Need for a Comprehensive, Coordinated Response from Health and Social Sectors at the Country Level: Report by the Secretariat; World Health Organization: Geneva, Swizerland, 2012; p. 4. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; DSM-5; American Psychiatric Association: Arlington, VA, USA, 2013; Volume 991, ISBN 978-0-89042-555-8. [Google Scholar]
- First, M.B. DSM-5® Handbook of Differential Diagnosis; American Psychiatric Publishing: Wasington, DC, USA, 2013. [Google Scholar]
- Kempton, M.J. Structural Neuroimaging Studies in Major Depressive Disorder. Arch. Gen. Psychiatry 2011, 68, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Videbech, P.; Ravnkilde, B. Reviews and Overviews Hippocampal Volume and Depression: A Meta-Analysis of MRI Studies. Hippocampal Vol. Depress. A Meta Anal. MRI Stud. 2004, 161, 1957–1966. [Google Scholar]
- Koolschijn, P.C.M.P.; Van Haren, N.E.M.; Lensvelt-Mulders, G.J.L.M.; Hulshoff Pol, H.E.; Kahn, R.S. Brain volume abnormalities in major depressive disorder: A meta-analysis of magnetic resonance imaging studies. Hum. Brain Mapp. 2009, 30, 3719–3735. [Google Scholar] [CrossRef] [PubMed]
- Bromis, K.; Calem, M.; Reinders, A.A.T.S.; Williams, S.C.R.; Kempton, M.J. Meta-Analysis of 89 Structural MRI studies in posttraumatic stress disorder and comparison with major depressive disorder. Am. J. Psychiatry 2018, 175, 989–998. [Google Scholar] [CrossRef]
- Roddy, D.W.; Farrell, C.; Doolin, K.; Roman, E.; Tozzi, L.; Frodl, T.; O’Keane, V.; O’Hanlon, E. The Hippocampus in Depression: More Than the Sum of Its Parts? Advanced Hippocampal Substructure Segmentation in Depression. Biol. Psychiatry 2019, 85, 487–497. [Google Scholar] [CrossRef]
- Czéh, B.; Lucassen, P.J. What causes the hippocampal volume decrease in depression? Are neurogenesis, glial changes and apoptosis implicated? Eur. Arch. Psychiatry Clin. Neurosci. 2007, 257, 250–260. [Google Scholar] [CrossRef]
- MacQueen, G.; Frodl, T. The hippocampus in major depression: Evidence for the convergence of the bench and bedside in psychiatric research. Mol. Psychiatry 2011, 16, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Malykhin, N.V.; Coupland, N.J. Hippocampal neuroplasticity in major depressive disorder. Neuroscience 2015, 309, 200–213. [Google Scholar] [CrossRef]
- Pittenger, C.; Duman, R.S. Stress, depression, and neuroplasticity: A convergence of mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef]
- Czéh, B.; Fuchs, E.; Wiborg, O.; Simon, M. Animal models of major depression and their clinical implications. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2016, 64, 293–310. [Google Scholar] [CrossRef]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology (Berlin) 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology (Berlin) 1997, 134, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiborg, O. Chronic mild stress for modeling anhedonia. Cell Tissue Res. 2013, 354, 155–169. [Google Scholar] [CrossRef]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci. Biobehav. Rev. 2019, 99, 101–116. [Google Scholar] [CrossRef]
- Vollmayr, B.; Gass, P. Learned helplessness: Unique features and translational value of a cognitive depression model. Cell Tissue Res. 2013, 354, 171–178. [Google Scholar] [CrossRef]
- Koo, J.W.; Chaudhury, D.; Han, M.H.; Nestler, E.J. Role of Mesolimbic Brain-Derived Neurotrophic Factor in Depression. Biol. Psychiatry 2019, 86, 738–748. [Google Scholar] [CrossRef]
- Bergström, A.; Jayatissa, M.N.; Mørk, A.; Wiborg, O. Stress sensitivity and resilience in the chronic mild stress rat model of depression; an in situ hybridization study. Brain Res. 2008, 1196, 41–52. [Google Scholar] [CrossRef]
- Martis, L.S.; Brision, C.; Holmes, M.C.; Wiborg, O. Resilient and depressive-like rats show distinct cognitive impairments in the touchscreen paired-associates learning (PAL) task. Neurobiol. Learn. Mem. 2018, 155, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Czéh, B.; Vardya, I.; Varga, Z.; Febbraro, F.; Csabai, D.; Martis, L.S.; Højgaard, K.; Henningsen, K.; Bouzinova, E.V.; Miseta, A.; et al. Long-term stress disrupts the structural and functional integrity of GABAergic neuronal networks in the medial prefrontal cortex of rats. Front. Cell. Neurosci. 2018, 12, 148. [Google Scholar] [CrossRef]
- Jayatissa, M.N.; Bisgaard, C.; Tingström, A.; Papp, M.; Wiborg, O. Hippocampal cytogenesis correlates to escitalopram-mediated recovery in a chronic mild stress rat model of depression. Neuropsychopharmacology 2006, 31, 2395–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belleau, E.L.; Treadway, M.T.; Pizzagalli, D.A. The Impact of Stress and Major Depressive Disorder on Hippocampal and Medial Prefrontal Cortex Morphology. Biol. Psychiatry 2019, 85, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Kroenke, C.D.; Wiborg, O.; Chuhutin, A.; Nyengaard, J.R.; Hansen, B.; Jespersen, S.N. Differential microstructural alterations in rat cerebral cortex in a model of chronic mild stress depression. PLoS ONE 2018, 13, e0192329. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Hansen, B.; Danladi, J.; Chuhutin, A.; Wiborg, O.; Nyengaard, J.R.; Jespersen, S.N. Neurite atrophy in dorsal hippocampus of rat indicates incomplete recovery of chronic mild stress induced depression. NMR Biomed. 2019, 32, e4057. [Google Scholar] [CrossRef]
- Khan, A.R.; Hansen, B.; Wiborg, O.; Kroenke, C.D.; Jespersen, S.N. Diffusion MRI and MR spectroscopy reveal microstructural and metabolic brain alterations in chronic mild stress exposed rats: A CMS recovery study. Neuroimage 2018, 167, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.P.; Ostrander, M.M.; Mueller, N.K.; Figueiredo, H. Limbic system mechanisms of stress regulation: Hypothalamo-pituitary- adrenocortical axis. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2005, 29, 1201–1213. [Google Scholar] [CrossRef]
- McEwen, B.S.; De Kloet, E.R.; Rostene, W. Adrenal steroid receptors and actions in the nervous system. Physiol. Rev. 1986, 66, 1121–1188. [Google Scholar] [CrossRef]
- McEwen, B.S. Stress and hippocampal plasticity. Annu. Rev. Neurosci. 1999, 22, 105–122. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Akil, H. Revisiting the stress concept: Implications for affective disorders. J. Neurosci. 2020, 40, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, H.; Kozlovsky, N.; Matar, M.A.; Zohar, J.; Kaplan, Z. Distinctive hippocampal and amygdalar cytoarchitectural changes underlie specific patterns of behavioral disruption following stress exposure in an animal model of PTSD. Eur. Neuropsychopharmacol. 2014, 24, 1925–1944. [Google Scholar] [CrossRef] [PubMed]
- Lucassen, P.J.; Pruessner, J.; Sousa, N.; Almeida, O.F.X.; Van Dam, A.M.; Rajkowska, G.; Swaab, D.F.; Czéh, B. Neuropathology of stress. Acta Neuropathol. 2014, 127, 109–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucassen, P.J.; Vollmann-Honsdorf, G.K.; Gleisberg, M.; Czéh, B.; De Kloet, E.R.; Fuchs, E. Chronic psychosocial stress differentially affects apoptosis in hippocampal subregions and cortex of the adult tree shrew. Eur. J. Neurosci. 2001, 14, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Lucassen, P.J.; Heine, V.; Muller, M.; van der Beek, E.; Wiegant, V.; De Kloet, R.E.; Joels, M.; Fuchs, E.; Swaab, D.; Czeh, B. Stress, Depression and Hippocampal Apoptosis. CNS Neurol. Disord. Drug Targets 2006, 5, 531–546. [Google Scholar] [CrossRef]
- Khan, A.R.; Chuhutin, A.; Wiborg, O.; Kroenke, C.D.; Nyengaard, J.R.; Hansen, B.; Jespersen, S.N. Biophysical modeling of high field diffusion MRI demonstrates micro-structural aberration in chronic mild stress rat brain. Neuroimage 2016, 142, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Chuhutin, A.; Wiborg, O.; Kroenke, C.D.; Nyengaard, J.R.; Hansen, B.; Jespersen, S.N. Summary of high field diffusion MRI and microscopy data demonstrate microstructural aberration in chronic mild stress rat brain. Data Br. 2016, 8, 934–937. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Jespersen, S.N.; Wiborg, O.; Kroenke, C.; Hansen, B. Microstructural and metabolic recovery of anhedonic rat brains: An in vivo diffusion MRI and 1H-MRS approach. Data 2018, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Sousa, N.; Lukoyanov, N.V.; Madeira, M.D.; Almeida, O.F.X.; Paula-Barbosa, M.M. Reorganization of the morphology of hippocampal neurites and synapses after stress-induced damage correlates with behavioral improvement. Neuroscience 2000, 97, 253–266. [Google Scholar] [CrossRef]
- Morais, M.; Santos, P.A.R.; Mateus-Pinheiro, A.; Patrício, P.; Pinto, L.; Sousa, N.; Pedroso, P.; Almeida, S.; Filipe, A.; Bessa, J.M. The effects of chronic stress on hippocampal adult neurogenesis and dendritic plasticity are reversed by selective MAO-A inhibition. J. Psychopharmacol. 2014, 28, 1178–1183. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; An, S.C.; Xu, C.; Ma, X.M. Role of proBDNF and BDNF in dendritic spine plasticity and depressive-like behaviors induced by an animal model of depression. Brain Res. 2017, 1663, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, V.; Costa, J.C.; Morgado, P.; Mota, C.; Miranda, A.; Bravo, F.V.; Oliveira, T.G.; Cerqueira, J.J.; Sousa, N. Differential impact of chronic stress along the hippocampal dorsal-ventral axis. Brain Struct. Funct. 2015, 220, 1205–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, P.C.; Tan, Z.N.; Jia, Z.Y.; Wang, B.; Grady, J.J.; Ma, X.M. Treadmill exercise reverses depression model-induced alteration of dendritic spines in the brain areas of mood circuit. Front. Behav. Neurosci. 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Jayatissa, M.N.; Henningsen, K.; West, M.J.; Wiborg, O. Decreased cell proliferation in the dentate gyrus does not associate with development of anhedonic-like symptoms in rats. Brain Res. 2009, 1290, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Kim, S.J.; Kim, S.W.; Choi, S.H.; Shin, Y.C.; Park, S.H.; Moon, B.H.; Cho, E.; Lee, M.S.; Choi, S.H.; et al. Chronic mild stress decreases survival, but not proliferation, of new-born cells in adult rat hippocampus. Exp. Mol. Med. 2006, 38, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mineur, Y.S.; Belzung, C.; Crusio, W.E. Functional implications of decreases in neurogenesis following chronic mild stress in mice. Neuroscience 2007, 150, 251–259. [Google Scholar] [CrossRef]
- Zhou, Q.G.; Hu, Y.; Hua, Y.; Hu, M.; Luo, C.X.; Han, X.; Zhu, X.J.; Wang, B.; Xu, J.S.; Zhu, D.Y. Neuronal nitric oxide synthase contributes to chronic stress-induced depression by suppressing hippocampal neurogenesis. J. Neurochem. 2007, 103, 1843–1854. [Google Scholar] [CrossRef]
- Goshen, I.; Kreisel, T.; Ben-Menachem-Zidon, O.; Licht, T.; Weidenfeld, J.; Ben-Hur, T.; Yirmiya, R. Brain interleukin-1 mediates chronic stress-induced depression in mice via adrenocortical activation and hippocampal neurogenesis suppression. Mol. Psychiatry 2008, 13, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Toth, E.; Gersner, R.; Wilf-Yarkoni, A.; Raizel, H.; Dar, D.E.; Richter-Levin, G.; Levit, O.; Zangen, A. Age-dependent effects of chronic stress on brain plasticity and depressive behavior. J. Neurochem. 2008, 107, 522–532. [Google Scholar] [CrossRef]
- Tanti, A.; Rainer, Q.; Minier, F.; Surget, A.; Belzung, C. Differential environmental regulation of neurogenesis along the septo-temporal axis of the hippocampus. Neuropharmacology 2012, 63, 374–384. [Google Scholar] [CrossRef]
- Tanti, A.; Westphal, W.P.; Girault, V.; Brizard, B.; Devers, S.; Leguisquet, A.M.; Surget, A.; Belzung, C. Region-dependent and stage-specific effects of stress, environmental enrichment, and antidepressant treatment on hippocampal neurogenesis. Hippocampus 2013, 23, 797–811. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, J.S.; Céspedes, I.C.; Abrão, R.O.; dos Santos, T.B.; Diniz, L.; Britto, L.R.G.; Spadari-Bratfisch, R.C.; Ortolani, D.; Melo-Thomas, L.; da Silva, R.C.B.; et al. Chronic unpredictable mild stress alters an anxiety-related defensive response, Fos immunoreactivity and hippocampal adult neurogenesis. Behav. Brain Res. 2013, 250, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Hattiangady, B.; Shuai, B.; Shetty, A.K. Mood and memory deficits in a model of gulf war illness are linked with reduced neurogenesis, partial neuron loss, and mild inflammation in the hippocampus. Neuropsychopharmacology 2013, 38, 2348–2362. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.D.; Correia, J.S.; Patrício, P.; Mateus-Pinheiro, A.; Machado-Santos, A.R.; Loureiro-Campos, E.; Morais, M.; Bessa, J.M.; Sousa, N.; Pinto, L. Adult hippocampal neuroplasticity triggers susceptibility to recurrent depression. Transl. Psychiatry 2017, 7, e1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culig, L.; Surget, A.; Bourdey, M.; Khemissi, W.; Le Guisquet, A.M.; Vogel, E.; Sahay, A.; Hen, R.; Belzung, C. Increasing adult hippocampal neurogenesis in mice after exposure to unpredictable chronic mild stress may counteract some of the effects of stress. Neuropharmacology 2017, 126, 179–189. [Google Scholar] [CrossRef]
- Tunc-Ozcan, E.; Peng, C.Y.; Zhu, Y.; Dunlop, S.R.; Contractor, A.; Kessler, J.A. Activating newborn neurons suppresses depression and anxiety-like behaviors. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Jayatissa, M.N.; Bisgaard, C.F.; West, M.J.; Wiborg, O. The number of granule cells in rat hippocampus is reduced after chronic mild stress and re-established after chronic escitalopram treatment. Neuropharmacology 2008, 54, 530–541. [Google Scholar] [CrossRef]
- Jayatissa, M.N.; Henningsen, K.; Nikolajsen, G.; West, M.J.; Wiborg, O. A reduced number of hippocampal granule cells does not associate with an anhedonia-like phenotype in a rat chronic mild stress model of depression. Stress 2010, 13, 95–105. [Google Scholar] [CrossRef]
- Yang, C.; Wang, G.; Wang, H.; Liu, Z.; Wang, X. Cytoskeletal alterations in rat hippocampus following chronic unpredictable mild stress and re-exposure to acute and chronic unpredictable mild stress. Behav. Brain Res. 2009, 205, 518–524. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Oitzl, M.S.; Champagne, D.L.; van der Veen, R.; de Kloet, E.R. Brain development under stress: Hypotheses of glucocorticoid actions revisited. Neurosci. Biobehav. Rev. 2010, 34, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. The neuroendocrinology of stress and aging: The glucocorticoid cascade hypothesis. Endocr. Rev. 1986, 7, 284–301. [Google Scholar] [CrossRef] [PubMed]
- Czeh, B.; Simon, M.; Van Der Hart, M.G.C.; Schmelting, B.; Hesselink, M.B.; Fuchs, E. Chronic stress decreases the number of parvalbumin-immunoreactive interneurons in the hippocampus: Prevention by treatment with a substance P receptor (NK1) antagonist. Neuropsychopharmacology 2005, 30, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Su, L.; Xu, Y.Q.; Zhang, H.; Wang, L.W. Behavioral and [F-18] fluorodeoxyglucose micro positron emission tomography imaging study in a rat chronic mild stress model of depression. Neuroscience 2010, 169, 171–181. [Google Scholar] [CrossRef]
- Gilabert-Juan, J.; Bueno-Fernandez, C.; Castillo-Gomez, E.; Nacher, J. Reduced interneuronal dendritic arborization in CA1 but not in CA3 region of mice subjected to chronic mild stress. Brain Behav. 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Banasr, M.; Lepack, A.; Fee, C.; Duric, V.; Maldonado-Aviles, J.; DiLeone, R.; Sibille, E.; Duman, R.S.; Sanacora, G. Characterization of GABAergic Marker Expression in the Chronic Unpredictable Stress Model of Depression. Chron. Stress 2017, 1, 247054701772045. [Google Scholar] [CrossRef] [Green Version]
- Czéh, B.; Varga, Z.K.K.; Henningsen, K.; Kovács, G.L.; Miseta, A.; Wiborg, O. Chronic stress reduces the number of GABAergic interneurons in the adult rat hippocampus, dorsal-ventral and region-specific differences. Hippocampus 2015, 25, 393–405. [Google Scholar] [CrossRef]
- Czéh, B.; Ábrahám, H.; Tahtakran, S.; Houser, C.R.; Seress, L. Number and regional distribution of GAD65 mRNA-expressing interneurons in the rat hippocampal formation. Acta Biol. Hung. 2013, 64, 395–413. [Google Scholar] [CrossRef]
- Roux, L.; Buzsáki, G. Tasks for inhibitory interneurons in intact brain circuits. Neuropharmacology 2015, 88, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Pelkey, K.A.; Chittajallu, R.; Craig, M.T.; Tricoire, L.; Wester, J.C.; McBain, C.J. Hippocampal gabaergic inhibitory interneurons. Physiol. Rev. 2017, 97, 1619–1747. [Google Scholar] [CrossRef]
- Ren, Z.; Pribiag, H.; Jefferson, S.J.; Shorey, M.; Fuchs, T.; Stellwagen, D.; Luscher, B. Bidirectional Homeostatic Regulation of a Depression-Related Brain State by Gamma-Aminobutyric Acidergic Deficits and Ketamine Treatment. Biol. Psychiatry 2016, 80, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Sanacora, G.; Krystal, J.H. Altered Connectivity in Depression: GABA and Glutamate Neurotransmitter Deficits and Reversal by Novel Treatments. Neuron 2019, 102, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Fogaça, M.V.; Duman, R.S. Cortical GABAergic dysfunction in stress and depression: New insights for therapeutic interventions. Front. Cell. Neurosci. 2019, 13, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Zhang, M.; Czéh, B.; Flügge, G.; Zhang, W. Stress impairs GABAergic network function in the hippocampus by activating nongenomic glucocorticoid receptors and affecting the integrity of the parvalbumin-expressing neuronal network. Neuropsychopharmacology 2010, 35, 1693–1707. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, A.C.; Paladini, M.S.; Colombo, M.; Gruca, P.; Lason-Tyburkiewicz, M.; Tota-Glowczyk, K.; Papp, M.; Riva, M.A.; Molteni, R. Chronic Stress Exposure Reduces Parvalbumin Expression in the Rat Hippocampus through an Imbalance of Redox Mechanisms: Restorative Effect of the Antipsychotic Lurasidone. Int. J. Neuropsychopharmacol. 2018, 21, 883–893. [Google Scholar] [CrossRef]
- Csabai, D.; Seress, L.; Varga, Z.; Ábrahám, H.; Miseta, A.; Wiborg, O.; Czéh, B. Electron Microscopic Analysis of Hippocampal Axo-Somatic Synapses in a Chronic Stress Model for Depression. Hippocampus 2017, 27, 17–27. [Google Scholar] [CrossRef]
- Magarinos, A.M.; Verdugo, J.M.G.; McEwen, B.S. Chronic stress alters synaptic terminal structure in hippocampus. Proc. Natl. Acad. Sci. USA 1997, 94, 14002–14008. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.G.; Davies, H.A.; Sandi, C.; Kraev, I.V.; Rogachevsky, V.V.; Peddie, C.J.; Rodriguez, J.J.; Cordero, M.I.; Donohue, H.S.; Gabbott, P.L.A.; et al. Stress suppresses and learning induces plasticity in CA3 of rat hippocampus: A three-dimensional ultrastructural study of thorny excrescences and their postsynaptic densities. Neuroscience 2005, 131, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Donohue, H.S.; Gabbott, P.L.A.; Davies, H.A.; Rodríguez, J.J.; Cordero, M.I.; Sandi, C.; Medvedev, N.I.; Popov, V.I.; Colyer, F.M.; Peddie, C.J.; et al. Chronic restraint stress induces changes in synapse morphology in stratum lacunosum-moleculare CA1 rat hippocampus: A stereological and three-dimensional ultrastructural study. Neuroscience 2006, 140, 597–606. [Google Scholar] [CrossRef]
- Hajszan, T.; Dow, A.; Warner-Schmidt, J.L.; Szigeti-Buck, K.; Sallam, N.L.; Parducz, A.; Leranth, C.; Duman, R.S. Remodeling of Hippocampal Spine Synapses in the Rat Learned Helplessness Model of Depression. Biol. Psychiatry 2009, 65, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-L.; Yuan, Y.-G.; Xu, H.; Wu, D.; Gong, W.-G.; Geng, L.-Y.; Wu, F.-F.; Tang, H.; Xu, L.; Zhang, Z.-J. Changed Synaptic Plasticity in Neural Circuits of Depressive-Like and Escitalopram-Treated Rats. Int. J. Neuropsychopharmacol. 2015, 18, pyv046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hei, M.; Chen, P.; Wang, S.; Li, X.; Xu, M.; Zhu, X.; Wang, Y.; Duan, J.; Huang, Y.; Zhao, S. Effects of chronic mild stress induced depression on synaptic plasticity in mouse hippocampus. Behav. Brain Res. 2019, 365, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.M.; Nieto-Gonzalez, J.L.; Vardya, I.; Henningsen, K.; Jayatissa, M.N.; Wiborg, O.; Jensen, K. Hippocampal GABAergic dysfunction in a rat chronic mild stress model of depression. Hippocampus 2011, 21, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhang, M.; Czéh, B.; Zhang, W.; Flügge, G. Chronic restraint stress impairs endocannabinoid mediated suppression of GABAergic signaling in the hippocampus of adult male rats. Brain Res. Bull. 2011, 85, 374–379. [Google Scholar] [CrossRef]
- Reich, C.G.; Mihalik, G.R.; Iskander, A.N.; Seckler, J.C.; Weiss, M.S. Adolescent chronic mild stress alters hippocampal CB1 receptor-mediated excitatory neurotransmission and plasticity. Neuroscience 2013, 253, 444–454. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; An, S.C.; Ren, W.; Ma, X.M. Progressive alterations of hippocampal CA3-CA1 synapses in an animal model of depression. Behav. Brain Res. 2014, 275, 191–200. [Google Scholar] [CrossRef]
- Bannerman, D.M.; Sprengel, R.; Sanderson, D.J.; Mchugh, S.B.; Rawlins, J.N.P.; Monyer, H.; Seeburg, P.H. Hippocampal synaptic plasticity, spatial memory and anxiety. Nat. Rev. Neurosci. 2014, 15, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Fanselow, M.S.; Dong, H.W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [Green Version]
- Anacker, C.; Hen, R. Adult hippocampal neurogenesis and cognitive flexibility-linking memory and mood. Nat. Rev. Neurosci. 2017, 18, 335–346. [Google Scholar] [CrossRef]
- Delgado, Y.; Palacios, R.; Campo, A.; Henningsen, K.; Verhoye, M.; Poot, D.; Dijkstra, J.; Van Audekerke, J.; Benveniste, H.; Sijbers, J.; et al. Magnetic resonance imaging and spectroscopy reveal differential hippocampal changes in anhedonic and resilient subtypes of the chronic mild stress rat model. Biol. Psychiatry 2011, 70, 449–457. [Google Scholar] [CrossRef]
- Stein-Behrens, B.A.; Lin, W.J.; Sapolsky, R.M. Physiological Elevations of Glucocorticoids Potentiate Glutamate Accumulation in the Hippocampus. J. Neurochem. 1994, 63, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, R.; Novais, A.; Barrie‘re, D.A.; Marques, P.; Marques, F.; Sousa, J.C.; Cerqueira, J.J.; Cachia, A.; Jay, T.M.; Bottlaender, M.; et al. A resting-state functional MR imaging and spectroscopy study of the dorsal hippocampus in the chronic unpredictable stress rat model. J. Neurosci. 2019, 39, 3640–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapolsky, R.M. The possibility of neurotoxicity in the hippocampus in major depression: A primer on neuron death. Biol. Psychiatry 2000, 48, 755–765. [Google Scholar] [CrossRef]
- Dantzer, R.; Walker, A.K. Is there a role for glutamate-mediated excitotoxicity in inflammation-induced depression? J. Neural Transm. 2014, 121, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Simon, M.; Schmelting, B.; Hiemke, C.; Fuchs, E. Astroglial plasticity in the hippocampus is affected by chronic psychosocial stress and concomitant fluoxetine treatment. Neuropsychopharmacology 2006, 31, 1616–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, S.; Taniguchi, M.; Koyama, Y.; Shimizu, S.; Tanaka, T.; Yasuno, F.; Yamamoto, A.; Iida, H.; Kudo, T.; Katayama, T.; et al. Association between chronic stress-induced structural abnormalities in Ranvier nodes and reduced oligodendrocyte activity in major depression. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tynan, R.J.; Naicker, S.; Hinwood, M.; Nalivaiko, E.; Buller, K.M.; Pow, D.V.; Day, T.A.; Walker, F.R. Chronic stress alters the density and morphology of microglia in a subset of stress-responsive brain regions. Brain Behav. Immun. 2010, 24, 1058–1068. [Google Scholar] [CrossRef]
- Czéh, B.; Nagy, S.A. Clinical findings documenting cellular and molecular abnormalities of glia in depressive disorders. Front. Mol. Neurosci. 2018, 11, 56. [Google Scholar] [CrossRef]
- Li, L.F.; Yang, J.; Ma, S.P.; Qu, R. Magnolol treatment reversed the glial pathology in an unpredictable chronic mild stress-induced rat model of depression. Eur. J. Pharmacol. 2013, 711, 42–49. [Google Scholar] [CrossRef]
- Machado-Santos, A.R.; Alves, N.D.; Araújo, B.; Correia, J.S.; Patrício, P.; Mateus-Pinheiro, A.; Loureiro-Campos, E.; Bessa, J.M.; Sousa, N.; Pinto, L. Astrocytic plasticity at the dorsal dentate gyrus on an animal model of recurrent depression. Neuroscience 2019, S0306-4522(19)30724-9, [Epub ahead of print]. [Google Scholar] [CrossRef]
- Wang, Y.L.; Han, Q.Q.; Gong, W.Q.; Pan, D.H.; Wang, L.Z.; Hu, W.; Yang, M.; Li, B.; Yu, J.; Liu, Q. Microglial activation mediates chronic mild stress-induced depressive- and anxiety-like behavior in adult rats. J. Neuroinflammation 2018, 15, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferle, V.; Repouskou, A.; Aspiotis, G.; Raftogianni, A.; Chrousos, G.; Stylianopoulou, F.; Stamatakis, A. Synergistic effects of early life mild adversity and chronic social defeat on rat brain microglia and cytokines. Physiol. Behav. 2020, 215, 112791. [Google Scholar] [CrossRef] [PubMed]
- Boda, E. Myelin and oligodendrocyte lineage cell dysfunctions: New players in the etiology and treatment of depression and stress-related disorders. Eur. J. Neurosci. 2019, ejn.14621, [Epub ahead of print]. [Google Scholar] [CrossRef] [PubMed]
- Cobb, J.A.; O’Neill, K.; Milner, J.; Mahajan, G.J.; Lawrence, T.J.; May, W.L.; Miguel-Hidalgo, J.; Rajkowska, G.; Stockmeier, C.A. Density of GFAP-immunoreactive astrocytes is decreased in left hippocampi in major depressive disorder. Neuroscience 2016, 316, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Innes, S.; Pariante, C.M.; Borsini, A. Microglial-driven changes in synaptic plasticity: A possible role in major depressive disorder. Psychoneuroendocrinology 2019, 102, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Jie, W.; Liu, J.-H.; Yang, J.-M.; Gao, T.-M. An astroglial basis of major depressive disorder? An overview. Glia 2017, 65, 1227–1250. [Google Scholar] [CrossRef]
- Murakami, S.; Imbe, H.; Morikawa, Y.; Kubo, C.; Senba, E. Chronic stress, as well as acute stress, reduces BDNF mRNA expression in the rat hippocampus but less robustly. Neurosci. Res. 2005, 53, 129–139. [Google Scholar] [CrossRef]
- Tornese, P.; Sala, N.; Bonini, D.; Bonifacino, T.; La Via, L.; Milanese, M.; Treccani, G.; Seguini, M.; Ieraci, A.; Mingardi, J.; et al. Chronic mild stress induces anhedonic behavior and changes in glutamate release, BDNF trafficking and dendrite morphology only in stress vulnerable rats. The rapid restorative action of ketamine. Neurobiol. Stress 2019, 10, 100160. [Google Scholar] [CrossRef]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [Green Version]
- Chattarji, S.; Tomar, A.; Suvrathan, A.; Ghosh, S.; Rahman, M.M. Neighborhood matters: Divergent patterns of stress-induced plasticity across the brain. Nat. Neurosci. 2015, 18, 1364–1375. [Google Scholar] [CrossRef]
- Christensen, T.; Bisgaard, C.F.; Wiborg, O. Biomarkers of anhedonic-like behavior, antidepressant drug refraction, and stress resilience in a rat model of depression. Neuroscience 2011, 196, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Groves, J.O. Is it time to reassess the BDNF hypothesis of depression? Mol. Psychiatry 2007, 12, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Martinowich, K.; Manji, H.; Lu, B. New insights into BDNF function in depression and anxiety. Nat. Neurosci. 2007, 10, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.A.; Aboharb, F.; Zeltner, N.; Studer, L. Pluripotent stem cells in neuropsychiatric disorders. Mol. Psychiatry 2017, 22, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Bardy, C.; Greenberg, Z.; Perry, S.W.; Licinio, J. Personalized psychiatry with human iPSCs and neuronal reprogramming. In Personalized Psychiatry; Elsevier: Amsterdam, The Netherlands, 2020; pp. 127–146. ISBN 9780128131763. [Google Scholar]
- Meijer, M.; Rehbach, K.; Brunner, J.W.; Classen, J.A.; Lammertse, H.C.A.; van Linge, L.A.; Schut, D.; Krutenko, T.; Hebisch, M.; Cornelisse, L.N.; et al. A Single-Cell Model for Synaptic Transmission and Plasticity in Human iPSC-Derived Neurons. Cell Rep. 2019, 27, 2199–2211.e6. [Google Scholar] [CrossRef] [Green Version]
- Arnsten, A.F.T. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Herman, J.P.; Prewitt, C.M.-F.; Cullinan, W.E. Neuronal Circuit Regulation of the Hypothalamo-Pituitary-Adrenocortical Stress Axis. Crit. Rev. Neurobiol. 1996, 10, 371–394. [Google Scholar] [CrossRef]
- Drevets, W.C.; Price, J.L.; Simpson, J.R.; Todd, R.D.; Reich, T.; Vannier, M.; Raichle, M.E. Subgenual prefrontal cortex abnormalities in mood disorders. Nature 1997, 386, 824–827. [Google Scholar] [CrossRef]
- Drevets, W.C. Neuroimaging studies of mood disorders. Biol. Psychiatry 2000, 48, 813–829. [Google Scholar] [CrossRef]
- Drevets, W.C. Functional anatomical abnormalities in limbic and prefrontal cortical structures in major depression. Prog. Brain Res. 2000, 126, 413–431. [Google Scholar] [PubMed]
- McEwen, B.S.; Morrison, J.H. The Brain on Stress: Vulnerability and Plasticity of the Prefrontal Cortex over the Life Course. Neuron 2013, 79, 16–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansell, E.B.; Rando, K.; Tuit, K.; Guarnaccia, J.; Sinha, R. Cumulative adversity and smaller gray matter volume in medial prefrontal, anterior cingulate, and insula regions. Biol. Psychiatry 2012, 72, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, G.L.; Bruss, J.; Denburg, N.L. Increased perceived stress is related to decreased prefrontal cortex volumes among older adults. J. Clin. Exp. Neuropsychol. 2017, 39, 313–325. [Google Scholar] [CrossRef]
- Savic, I.; Perski, A.; Osika, W. MRI Shows that Exhaustion Syndrome Due to Chronic Occupational Stress is Associated with Partially Reversible Cerebral Changes. Cereb. Cortex 2018, 28, 894–906. [Google Scholar] [CrossRef] [Green Version]
- Csabai, D.; Wiborg, O.; Czéh, B. Reduced synapse and axon numbers in the prefrontal cortex of rats subjected to a chronic stress model for depression. Front. Cell. Neurosci. 2018, 12, 1–15. [Google Scholar] [CrossRef]
- Banasr, M.; Valentine, G.W.; Li, X.Y.; Gourley, S.L.; Taylor, J.R.; Duman, R.S. Chronic Unpredictable Stress Decreases Cell Proliferation in the Cerebral Cortex of the Adult Rat. Biol. Psychiatry 2007, 62, 496–504. [Google Scholar] [CrossRef]
- Czéh, B.; Perez-Cruz, C.; Fuchs, E.; Flügge, G. Chronic stress-induced cellular changes in the medial prefrontal cortex and their potential clinical implications: Does hemisphere location matter? Behav. Brain Res. 2008, 190, 1–13. [Google Scholar] [CrossRef]
- Bachis, A.; Cruz, M.I.; Nosheny, R.L.; Mocchetti, I. Chronic unpredictable stress promotes neuronal apoptosis in the cerebral cortex. Neurosci. Lett. 2008, 442, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Dias-Ferreira, E.; Sousa, J.C.; Melo, I.; Morgado, P.; Mesquita, A.R.; Cerqueira, J.J.; Costa, R.M.; Sousa, N. Chronic stress causes frontostriatal reorganization and affects decision-making. Science (80-. ) 2009, 325, 621–625. [Google Scholar] [CrossRef]
- Kafetzopoulos, V.; Kokras, N.; Sotiropoulos, I.; Oliveira, J.F.; Leite-Almeida, H.; Vasalou, A.; Sardinha, V.M.; Papadopoulou-Daifoti, Z.; Almeida, O.F.X.; Antoniou, K.; et al. The nucleus reuniens: A key node in the neurocircuitry of stress and depression. Mol. Psychiatry 2017, 23, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Liu, R.J.; Dwyer, J.M.; Banasr, M.; Lee, B.; Son, H.; Li, X.Y.; Aghajanian, G.; Duman, R.S. Glutamate N-methyl-D-aspartate receptor antagonists rapidly reverse behavioral and synaptic deficits caused by chronic stress exposure. Biol. Psychiatry 2011, 69, 754–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueira, J.J.; Taipa, R.; Uylings, H.B.M.; Almeida, O.F.X.; Sousa, N. Specific configuration of dendritic degeneration in pyramidal neurons of the medial prefrontal cortex induced by differing corticosteroid regimens. Cereb. Cortex 2007, 17, 1998–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemanth Kumar, B.S.; Mishra, S.K.; Rana, P.; Singh, S.; Khushu, S. Neurodegenerative evidences during early onset of depression in CMS rats as detected by proton magnetic resonance spectroscopy at 7T. Behav. Brain Res. 2012, 232, 53–59. [Google Scholar] [CrossRef]
- McKlveen, J.M.; Moloney, R.D.; Scheimann, J.R.; Myers, B.; Herman, J.P. “Braking” the Prefrontal Cortex: The Role of Glucocorticoids and Interneurons in Stress Adaptation and Pathology. Biol. Psychiatry 2019, 86, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, S.; Hare, B.D.; Duman, R.S. Prefrontal cortex GABAergic deficits and circuit dysfunction in the pathophysiology and treatment of chronic stress and depression. Curr. Opin. Behav. Sci. 2017, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- McKlveen, J.M.; Morano, R.L.; Fitzgerald, M.; Zoubovsky, S.; Cassella, S.N.; Scheimann, J.R.; Ghosal, S.; Mahbod, P.; Packard, B.A.; Myers, B.; et al. Chronic Stress Increases Prefrontal Inhibition: A Mechanism for Stress-Induced Prefrontal Dysfunction. Biol. Psychiatry 2016, 80, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Gilabert-Juan, J.; Castillo-Gomez, E.; Guirado, R.; Moltó, M.D.; Nacher, J. Chronic stress alters inhibitory networks in the medial prefrontal cortex of adult mice. Brain Struct. Funct. 2013, 218, 1591–1605. [Google Scholar] [CrossRef]
- Shepard, R.; Page, C.E.; Coutellier, L. Sensitivity of the prefrontal GABAergic system to chronic stress in male and female mice: Relevance for sex differences in stress-related disorders. Neuroscience 2016, 332, 1–12. [Google Scholar] [CrossRef]
- Shepard, R.; Coutellier, L. Changes in the Prefrontal Glutamatergic and Parvalbumin Systems of Mice Exposed to Unpredictable Chronic Stress. Mol. Neurobiol. 2018, 55, 2591–2602. [Google Scholar] [CrossRef] [PubMed]
- Page, C.E.; Shepard, R.; Heslin, K.; Coutellier, L. Prefrontal parvalbumin cells are sensitive to stress and mediate anxiety-related behaviors in female mice. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Maguire, J. Neuroactive Steroids and GABAergic Involvement in the Neuroendocrine Dysfunction Associated With Major Depressive Disorder and Postpartum Depression. Front. Cell. Neurosci. 2019, 13, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.J.; Voleti, B.; Hajszan, T.; Rajkowska, G.; Stockmeier, C.A.; Licznerski, P.; Lepack, A.; Majik, M.S.; Jeong, L.S.; Banasr, M.; et al. Decreased expression of synapse-related genes and loss of synapses in major depressive disorder. Nat. Med. 2012, 18, 1413–1417. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.E.; Scheinost, D.; Finnema, S.J.; Naganawa, M.; Davis, M.T.; DellaGioia, N.; Nabulsi, N.; Matuskey, D.; Angarita, G.A.; Pietrzak, R.H.; et al. Lower synaptic density is associated with depression severity and network alterations. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banasr, M.; Duman, R.S. Glial Loss in the Prefrontal Cortex Is Sufficient to Induce Depressive-like Behaviors. Biol. Psychiatry 2008, 64, 863–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banasr, M.; Chowdhury, G.M.I.; Terwilliger, R.; Newton, S.S.; Duman, R.S.; Behar, K.L.; Sanacora, G. Glial pathology in an animal model of depression: Reversal of stress-induced cellular, metabolic and behavioral deficits by the glutamate-modulating drug riluzole. Mol. Psychiatry 2010, 15, 501–511. [Google Scholar] [CrossRef]
- Hinwood, M.; Morandini, J.; Day, T.A.; Walker, F.R. Evidence that Microglia Mediate the Neurobiological Effects of Chronic Psychological Stress on the Medial Prefrontal Cortex. Cereb. Cortex 2012, 22, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Hinwood, M.; Tynan, R.J.; Charnley, J.L.; Beynon, S.B.; Day, T.A.; Rohan Walker, F. Chronic Stress Induced Remodeling of the Prefrontal Cortex: Structural Re-Organization of Microglia and the Inhibitory Effect of Minocycline. Cereb. Cortex 2013, 23, 1784–1797. [Google Scholar] [CrossRef] [Green Version]
- Tynan, R.J.; Beynon, S.B.; Hinwood, M.; Johnson, S.J.; Nilsson, M.; Woods, J.J.; Walker, F.R. Chronic stress-induced disruption of the astrocyte network is driven by structural atrophy and not loss of astrocytes. Acta Neuropathol. 2013, 126, 75–91. [Google Scholar] [CrossRef]
- Bollinger, J.L.; Bergeon Burns, C.M.; Wellman, C.L. Differential effects of stress on microglial cell activation in male and female medial prefrontal cortex. Brain. Behav. Immun. 2016, 52, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.L.; Weigel, T.K.; Elkahloun, A.G.; Herkenham, M. Chronic social defeat reduces myelination in the mouse medial prefrontal cortex. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wohleb, E.S.; Terwilliger, R.; Duman, C.H.; Duman, R.S. Stress-Induced Neuronal Colony Stimulating Factor 1 Provokes Microglia-Mediated Neuronal Remodeling and Depressive-like Behavior. Biol. Psychiatry 2018, 83, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Cathomas, F.; Azzinnari, D.; Bergamini, G.; Sigrist, H.; Buerge, M.; Hoop, V.; Wicki, B.; Goetze, L.; Soares, S.; Kukelova, D.; et al. Oligodendrocyte gene expression is reduced by and influences effects of chronic social stress in mice. Genes Brain Behav. 2019, 18, e12475. [Google Scholar] [CrossRef] [PubMed]
- Rajkowska, G.; Miguel-Hidalgo, J.J.; Wei, J.; Dilley, G.; Pittman, S.D.; Meltzer, H.Y.; Overholser, J.C.; Roth, B.L.; Stockmeier, C.A. Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression∗∗See accompanying Editorial, in this issue. Biol. Psychiatry 1999, 45, 1085–1098. [Google Scholar] [CrossRef]
- Rajkowska, G. Postmortem studies in mood disorders indicate altered numbers of neurons and glial cells. Biol. Psychiatry 2000, 48, 766–777. [Google Scholar] [CrossRef]
- Bender, C.L.; Calfa, G.D.; Molina, V.A. Astrocyte plasticity induced by emotional stress: A new partner in psychiatric physiopathology? Prog. Neuro Psychopharmacol. Biol. Psychiatry 2016, 65, 68–77. [Google Scholar] [CrossRef]
- Liu, J.; Dietz, K.; Hodes, G.E.; Russo, S.J.; Casaccia, P. Widespread transcriptional alternations in oligodendrocytes in the adult mouse brain following chronic stress. Dev. Neurobiol. 2018, 78, 152–162. [Google Scholar] [CrossRef]
- Canbeyli, R. Sensorimotor modulation of mood and depression: An integrative review. Behav. Brain Res. 2010, 207, 249–264. [Google Scholar] [CrossRef]
- Buyukdura, J.S.; McClintock, S.M.; Croarkin, P.E. Psychomotor retardation in depression: Biological underpinnings, measurement, and treatment. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2011, 35, 395–409. [Google Scholar] [CrossRef] [Green Version]
- Bracht, T.; Federspiel, A.; Schnell, S.; Horn, H.; Höfle, O.; Wiest, R.; Dierks, T.; Strik, W.; Müller, T.J.; Walther, S. Cortico-Cortical White Matter Motor Pathway Microstructure Is Related to Psychomotor Retardation in Major Depressive Disorder. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Wang, M.; Wang, Z.; Xie, C.; Zhang, H.; Zhang, H.; Zhang, Z.; Yuan, Y. Decreased cerebral blood flow in the primary motor cortex in major depressive disorder with psychomotor retardation. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 81, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, L.; Wu, M.; Chen, Z.; Hu, X.; Chen, Y.; Zhu, H.; Jia, Z.; Gong, Q. Brain gray matter alterations in first episodes of depression: A meta-analysis of whole-brain studies. Neurosci. Biobehav. Rev. 2016, 60, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, S.; Yamashita, H.; Rytsälä, H.; Suominen, K.; Ahveninen, J.; Isometsä, E. Dysfunction in early auditory processing in major depressive disorder revealed by combined MEG and EEG. J Psychiatry Neurosci. 2007, 32, 316–322. [Google Scholar] [PubMed]
- Smiley, J.F.; Hackett, T.A.; Bleiwas, C.; Petkova, E.; Stankov, A.; Mann, J.J.; Rosoklija, G.; Dwork, A.J. Reduced GABA neuron density in auditory cerebral cortex of subjects with major depressive disorder. J. Chem. Neuroanat. 2016, 76, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Grieve, S.M.; Korgaonkar, M.S.; Koslow, S.H.; Gordon, E.; Williams, L.M. Widespread reductions in gray matter volume in depression. NeuroImage Clin. 2013, 3, 332–339. [Google Scholar] [CrossRef] [Green Version]
- Järnum, H.; Eskildsen, S.F.; Steffensen, E.G.; Lundbye-Christensen, S.; Simonsen, C.W.; Thomsen, I.S.; Fründ, E.-T.; Théberge, J.; Larsson, E.-M. Longitudinal MRI study of cortical thickness, perfusion, and metabolite levels in major depressive disorder. Acta Psychiatr. Scand. 2011, 124, 435–446. [Google Scholar] [CrossRef]
- Magalhães, R.; Barrière, D.A.; Novais, A.; Marques, F.; Marques, P.; Cerqueira, J.; Sousa, J.C.; Cachia, A.; Boumezbeur, F.; Bottlaender, M.; et al. The dynamics of stress: A longitudinal MRI study of rat brain structure and connectome. Mol. Psychiatry 2018, 23, 1998–2006. [Google Scholar] [CrossRef]
- Maren, S.; Yap, S.A.; Goosens, K.A. The amygdala is essential for the development of neuronal plasticity in the medial geniculate nucleus during auditory fear conditioning in rats. J. Neurosci. 2001, 21, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gong, Q.; Wu, Q.; Scarpazza, C.; Lui, S.; Jia, Z.; Marquand, A.; Huang, X.; McGuire, P.; Mechelli, A. Prognostic prediction of therapeutic response in depression using high-field MR imaging. Neuroimage 2011, 55, 1497–1503. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Du, M.Y.; Huang, X.Q.; Lui, S.; Chen, Z.Q.; Liu, J.; Luo, Y.; Wang, X.L.; Kemp, G.J.; Gong, Q.Y. Brain grey matter abnormalities in medication-free patients with major depressive disorder: A meta-analysis. Psychol. Med. 2014, 44, 2927–2937. [Google Scholar] [CrossRef] [Green Version]
- Roozendaal, B.; McEwen, B.S.; Chattarji, S. Stress, memory and the amygdala. Nat. Rev. Neurosci. 2009, 10, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Bylsma, L.M.; Morris, B.H.; Rottenberg, J. A meta-analysis of emotional reactivity in major depressive disorder. Clin. Psychol. Rev. 2008, 28, 676–691. [Google Scholar] [CrossRef]
- Hamilton, J.P.; Siemer, M.; Gotlib, I.H. Amygdala volume in major depressive disorder: A meta-analysis of magnetic resonance imaging studies. Mol. Psychiatry 2008, 13, 993–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheline, Y.I.; Barch, D.M.; Donnelly, J.M.; Ollinger, J.M.; Snyder, A.Z.; Mintun, M.A. Increased amygdala response to masked emotional faces in depressed subjects resolves with antidepressant treatment: An fMRI study. Biol. Psychiatry 2001, 50, 651–658. [Google Scholar] [CrossRef]
- Vyas, A.; Mitra, R.; Shankaranarayana Rao, B.S.; Chattarji, S. Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons. J. Neurosci. 2002, 22, 6810–6818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, A.; Pillai, A.G.; Chattarji, S. Recovery after chronic stress fails to reverse amygdaloid neuronal hypertrophy and enhanced anxiety-like behavior. Neuroscience 2004, 128, 667–673. [Google Scholar] [CrossRef]
- Roozendaal, B.; McReynolds, J.R.; McGaugh, J.L. The Basolateral Amygdala Interacts with the Medial Prefrontal Cortex in Regulating Glucocorticoid Effects on Working Memory Impairment. J. Neurosci. 2004, 24, 1385–1392. [Google Scholar] [CrossRef]
- Wilson, M.A.; Grillo, C.A.; Fadel, J.R.; Reagan, L.P. Stress as a one-armed bandit: Differential effects of stress paradigms on the morphology, neurochemistry and behavior in the rodent amygdala. Neurobiol. Stress 2015, 1, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Réus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The role of inflammation and microglial activation in the pathophysiology of psychiatric disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Franklin, T.; Iwata, M.; Duman, R.S. Integrating neuroimmune systems in the neurobiology of depression. Nat. Rev. Neurosci. 2016, 17, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Bourgin, J.; Cachia, A.; Boumezbeur, F.; Djemaï, B.; Bottlaender, M.; Duchesnay, E.; Mériaux, S.; Jay, T.M. Hyper-responsivity to stress in rats is associated with a large increase in amygdala volume. A 7T MRI study. Eur. Neuropsychopharmacol. 2015, 25, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Shors, T.J. Stress, anxiety, and dendritic spines: What are the connections? Neuroscience 2013, 251, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Sibille, E.; Wang, Y.; Joeyen-Waldorf, J.; Gaiteri, C.; Surget, A.; Oh, S.; Belzung, C.; Tseng, G.C.; Lewis, D.A. A molecular signature of depression in the amygdala. Am. J. Psychiatry 2009, 166, 1011–1024. [Google Scholar] [CrossRef] [Green Version]
- Savalli, G.; Diao, W.; Schulz, S.; Todtova, K.; Pollak, D.D. Diurnal oscillation of Amygdala clock gene expression and loss of synchrony in a mouse model of depression. Int. J. Neuropsychopharmacol. 2015, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bora, E.; Harrison, B.J.; Davey, C.G.; Yücel, M.; Pantelis, C. Meta-analysis of volumetric abnormalities in cortico-striatal-pallidal- thalamic circuits in major depressive disorder. Psychol. Med. 2012, 42, 671–681. [Google Scholar] [CrossRef]
- Pizzagalli, D.A.; Holmes, A.J.; Dillon, D.G.; Goetz, E.L.; Birk, J.L.; Bogdan, R.; Dougherty, D.D.; Iosifescu, D.V.; Rauch, S.L.; Fava, M. Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder. Am. J. Psychiatry 2009, 166, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Khundakar, A.; Morris, C.; Oakley, A.; Thomas, A.J. Morphometric analysis of neuronal and glial cell pathology in the caudate nucleus in late-life depression. Am. J. Geriatr. Psychiatry 2011, 19, 132–141. [Google Scholar] [CrossRef]
- Hickie, I.; Ward, P.; Scott, E.; Haindl, W.; Walker, B.; Dixon, J.; Turner, K. Neo-striatal rCBF correlates of psychomotor slowing in patients with major depression. Psychiatry Res. Neuroimaging 1999, 92, 75–81. [Google Scholar] [CrossRef]
- Delgado y Palacios, R.; Verhoye, M.; Henningsen, K.; Wiborg, O.; Van der Linden, A. Diffusion Kurtosis Imaging and High-Resolution MRI Demonstrate Structural Aberrations of Caudate Putamen and Amygdala after Chronic Mild Stress. PLoS ONE 2014, 9, e95077. [Google Scholar] [CrossRef] [Green Version]
- Hemanth Kumar, B.S.; Mishra, S.K.; Trivedi, R.; Singh, S.; Rana, P.; Khushu, S. Demyelinating evidences in CMS rat model of depression: A DTI study at 7T. Neuroscience 2014, 275, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Van Velzen, L.S.; Kelly, S.; Isaev, D.; Aleman, A.; Aftanas, L.I.; Bauer, J.; Baune, B.T.; Brak, I.V.; Carballedo, A.; Connolly, C.G.; et al. White matter disturbances in major depressive disorder: A coordinated analysis across 20 international cohorts in the ENIGMA MDD working group. Mol. Psychiatry 2019, 1–15, [Epub ahead of print]. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillon, D.G.; Gonenc, A.; Belleau, E.; Pizzagalli, D.A. Depression is associated with dimensional and categorical effects on white matter pathways. Depress. Anxiety 2018, 35, 440–447. [Google Scholar] [CrossRef]
- Bracht, T.; Horn, H.; Strik, W.; Federspiel, A.; Schnell, S.; Höfle, O.; Stegmayer, K.; Wiest, R.; Dierks, T.; Müller, T.J.; et al. White matter microstructure alterations of the medial forebrain bundle in melancholic depression. J. Affect. Disord. 2014, 155, 186–193. [Google Scholar] [CrossRef]
- Kamiya, K.; Okada, N.; Sawada, K.; Watanabe, Y.; Irie, R.; Hanaoka, S.; Suzuki, Y.; Koike, S.; Mori, H.; Kunimatsu, A.; et al. Diffusional kurtosis imaging and white matter microstructure modeling in a clinical study of major depressive disorder. NMR Biomed. 2018, 31, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fieremans, E.; Jensen, J.H.; Helpern, J.A. White matter characterization with diffusional kurtosis imaging. Neuroimage 2011, 58, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.; Jespersen, S.N. Recent developments in fast kurtosis imaging. Front. Phys. 2017, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lawson, R.P.; Nord, C.L.; Seymour, B.; Thomas, D.L.; Dayan, P.; Pilling, S.; Roiser, J.P. Disrupted habenula function in major depression. Mol. Psychiatry 2017, 22, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Savitz, J.B.; Nugent, A.C.; Bogers, W.; Roiser, J.P.; Bain, E.E.; Neumeister, A.; Zarate, C.A.; Manji, H.K.; Cannon, D.M.; Marrett, S.; et al. Habenula volume in bipolar disorder and major depressive disorder: A high-resolution magnetic resonance imaging study. Biol. Psychiatry 2011, 69, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zhou, T.; Liao, L.; Yang, Z.; Wong, C.; Henn, F.; Malinow, R.; Yates, J.R.; Hu, H. βCaMKII in lateral habenula mediates core symptoms of depression. Science (80-.) 2013, 341, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Christensen, T.; Jensen, L.; Bouzinova, E.V.; Wiborg, O. Molecular profiling of the lateral habenula in a rat model of depression. PLoS ONE 2013, 8, e80666. [Google Scholar] [CrossRef]
- Shen, X.F.; Yuan, H.B.; Wang, G.Q.; Xue, H.; Liu, Y.F.; Zhang, C.X. Role of DNA hypomethylation in lateral habenular nucleus in the development of depressive-like behavior in rats. J. Affect. Disord. 2019, 252, 373–381. [Google Scholar] [CrossRef]
- Faron-Górecka, A.; Kuśmider, M.; Kolasa, M.; Zurawek, D.; Szafran-Pilch, K.; Gruca, P.; Pabian, P.; Solich, J.; Papp, M.; Dziedzicka-Wasylewska, M. Chronic mild stress alters the somatostatin receptors in the rat brain. Psychopharmacology (Berlin) 2016, 233, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Ågren, H.; Lundqvist, G. Low levels of somatostatin in human CSF mark depressive episodes. Psychoneuroendocrinology 1984, 9, 233–248. [Google Scholar] [CrossRef]
- Lin, L.C.; Sibille, E. Somatostatin, neuronal vulnerability and behavioral emotionality. Mol. Psychiatry 2015, 20, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Fee, C.; Banasr, M.; Sibille, E. Somatostatin-Positive Gamma-Aminobutyric Acid Interneuron Deficits in Depression: Cortical Microcircuit and Therapeutic Perspectives. Biol. Psychiatry 2017, 82, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Carlezon, W.A.; Thomas, M.J. Biological substrates of reward and aversion: A nucleus accumbens activity hypothesis. Neuropharmacology 2009, 56, 122–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, J.M.; Morais, M.; Marques, F.; Pinto, L.; Palha, J.A.; Almeida, O.F.X.; Sousa, N. Stress-induced anhedonia is associated with hypertrophy of medium spiny neurons of the nucleus accumbens. Transl. Psychiatry 2013, 3, e266–e267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, R.W.; Edgar, N.; Gillman, A.G.; Hoffman, D.; Zhu, X.; McClung, C.A. Chronic Stress Induces Brain Region-Specific Alterations of Molecular Rhythms that Correlate with Depression-like Behavior in Mice. Biol. Psychiatry 2015, 78, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress. Anxiety 2000, 12, 2–19. [Google Scholar] [CrossRef]
- Schildkraut, J.J. The catecholamine hypothesis of affective disorders: A review of supporting evidence. Am. J. Psychiatry 1965, 122, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Schildkraut, J.J.; Kety, S.S. Biogenic amines and emotion. Science (80-. ) 1967, 156, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Massart, R.; Mongeau, R.; Lanfumey, L. Beyond the monoaminergic hypothesis: Neuroplasticity and epigenetic changes in a transgenic mouse model of depression. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, K.; Chen, H.S.; Gao, S.Q.; Xia, Z.X.; Zhang, J.T.; Wang, F.; Chen, J.G. Dorsal raphe projection inhibits the excitatory inputs on lateral habenula and alleviates depressive behaviors in rats. Brain Struct. Funct. 2018, 223, 2243–2258. [Google Scholar] [CrossRef] [PubMed]
- Germain, A.; Kupfer, D.J. Circadian rhythm disturbances in depression. Hum. Psychopharmacol. Clin. Exp. 2008, 23, 571–585. [Google Scholar] [CrossRef] [Green Version]
- Bunney, B.G.; Li, J.Z.; Walsh, D.M.; Stein, R.; Vawter, M.P.; Cartagena, P.; Barchas, J.D.; Schatzberg, A.F.; Myers, R.M.; Watson, S.J.; et al. Circadian dysregulation of clock genes: Clues to rapid treatments in major depressive disorder. Mol. Psychiatry 2015, 20, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Bunney, W.E.; Bunney, B.G. Molecular clock genes in man and lower animals: Possible implications for circadian abnormalities in depression. Neuropsychopharmacology 2000, 22, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Slack, F.J. microRNAs: Small molecules with big roles -C. elegans to human cancer. Biol. Cell 2008, 100, 71–81. [Google Scholar] [CrossRef]
- Lopez, J.P.; Kos, A.; Turecki, G. Major depression and its treatment: MicroRNAs as peripheral biomarkers of diagnosis and treatment response. Curr. Opin. Psychiatry 2018, 31, 7–16. [Google Scholar] [CrossRef]
- Ferrúa, C.P.; Giorgi, R.; da Rosa, L.C.; do Amaral, C.C.; Ghisleni, G.C.; Pinheiro, R.T.; Nedel, F. MicroRNAs expressed in depression and their associated pathways: A systematic review and a bioinformatics analysis. J. Chem. Neuroanat. 2019, 100, 101650. [Google Scholar] [CrossRef]
- Fries, G.R.; Zhang, W.; Benevenuto, D.; Quevedo, J. MicroRNAs in Major Depressive Disorder. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2019; Volume 1118, pp. 175–190. [Google Scholar]
- Cogswell, J.; Ward, J.; Taylor, I.; Waters, M.; Shi, Y.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; Chen, C.; et al. Identification of miRNA Changes in Alzheimer’s.pdf. J. Alzheimer’s Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, N.J.; Cairns, M.J. MicroRNA dysregulation in schizophrenia. Neurobiol. Dis. 2012, 46, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos, K.; Malenica, I.; Metpally, R.; Courtright, A.; Rakela, B.; Beach, T.; Shill, H.; Adler, C.; Sabbagh, M.; Villa, S.; et al. Profiles of extracellular miRNA in cerebrospinal fluid and serum from patients with Alzheimer’s and Parkinson’s diseases correlate with disease status and features of pathology. PLoS ONE 2014, 9, e94839. [Google Scholar] [CrossRef]
- Moreau, M.P.; Bruse, S.E.; David-Rus, R.; Buyske, S.; Brzustowicz, L.M. Altered MicroRNA expression profiles in postmortem brain samples from individuals with schizophrenia and bipolar disorder. Biol. Psychiatry 2011, 69, 188–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, H.R.; Schoenfeld, L.W.; Ruby, J.G.; Auyeung, V.C.; Spies, N.; Baek, D.; Johnston, W.K.; Russ, C.; Luo, S.; Babiarz, J.E.; et al. Mammalian microRNAs: Experimental evaluation of novel and previously annotated genes. Genes Dev. 2010, 24, 992–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, B.M.; Heimberg, A.M.; Moy, V.N.; Sperling, E.A.; Holstein, T.W.; Heber, S.; Peterson, K.J. The deep evolution of metazoan microRNAs. Evol. Dev. 2009, 11, 50–68. [Google Scholar] [CrossRef]
- Meerson, A.; Cacheaux, L.; Goosens, K.A.; Sapolsky, R.M.; Soreq, H.; Kaufer, D. Changes in brain MicroRNAs contribute to cholinergic stress reactions. J. Mol. Neurosci. 2010, 40, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, A.; Vincenti, S.; De Vito, F.; Bozzoni, I.; Oliverio, A.; Presutti, C.; Fragapane, P.; Mele, A. Stress induces region specific alterations in microRNAs expression in mice. Behav. Brain Res. 2010, 208, 265–269. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, M.; Wang, X.; Liu, K.; Wan, Y.Q.; Li, M.; Liu, L.; Zhang, C. Abnormal Expression of MicroRNAs Induced by Chronic Unpredictable Mild Stress in Rat Hippocampal Tissues. Mol. Neurobiol. 2018, 55, 917–935. [Google Scholar] [CrossRef]
- Buran, İ.; Etem, E.Ö.; Tektemur, A.; Elyas, H. Treatment with TREK1 and TRPC3/6 ion channel inhibitors upregulates microRNA expression in a mouse model of chronic mild stress. Neurosci. Lett. 2017, 656, 51–57. [Google Scholar] [CrossRef]
- Dwivedi, Y. Evidence demonstrating role of microRNAs in the etiopathology of major depression. J. Chem. Neuroanat. 2011, 42, 142–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocchio-Chiavetto, L.; Maffioletti, E.; Bettinsoli, P.; Giovannini, C.; Bignotti, S.; Tardito, D.; Corrada, D.; Milanesi, L.; Gennarelli, M. Blood microRNA changes in depressed patients during antidepressant treatment. Eur. Neuropsychopharmacol. 2013, 23, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Zurawek, D.; Kusmider, M.; Faron-Gorecka, A.; Gruca, P.; Pabian, P.; Solich, J.; Kolasa, M.; Papp, M.; Dziedzicka-Wasylewska, M. Reciprocal MicroRNA Expression in Mesocortical Circuit and Its Interplay with Serotonin Transporter Define Resilient Rats in the Chronic Mild Stress. Mol. Neurobiol. 2017, 54, 5741–5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurawek, D.; Kusmider, M.; Faron-Gorecka, A.; Gruca, P.; Pabian, P.; Kolasa, M.; Solich, J.; Szafran-Pilch, K.; Papp, M.; Dziedzicka-Wasylewska, M. Time-dependent miR-16 serum fluctuations together with reciprocal changes in the expression level of miR-16 in mesocortical circuit contribute to stress resilient phenotype in chronic mild stress - An animal model of depression. Eur. Neuropsychopharmacol. 2016, 26, 23–36. [Google Scholar] [CrossRef]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.M.; Kellermann, O. MiR-16 targets the serotonin transporter: A new facet for adaptive responses to antidepressants. Science (80-.) 2010, 329, 1537–1541. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, F.; Uchida, S.; Yamagata, H.; Abe-Higuchi, N.; Hobara, T.; Hara, K.; Kobayashi, A.; Shintaku, T.; Itoh, Y.; Suzuki, T.; et al. Hippocampal microRNA-124 enhances chronic stress resilience in mice. J. Neurosci. 2016, 36, 7253–7267. [Google Scholar] [CrossRef]
- Yu, H.; Fan, C.; Yang, L.; Yu, S.; Song, Q.; Wang, P.; Mao, X. Ginsenoside Rg1 Prevents Chronic Stress-Induced Depression-Like Behaviors and Neuronal Structural Plasticity in Rats. Cell. Physiol. Biochem. 2018, 48, 2470–2482. [Google Scholar] [CrossRef]
- Bahi, A.; Chandrasekar, V.; Dreyer, J.L. Selective lentiviral-mediated suppression of microRNA124a in the hippocampus evokes antidepressants-like effects in rats. Psychoneuroendocrinology 2014, 46, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Schratt, G.M.; Tuebing, F.; Nigh, E.A.; Kane, C.G.; Sabatini, M.E.; Kiebler, M.; Greenberg, M.E. A brain-specific microRNA regulates dendritic spine development. Nature 2006, 439, 283–289. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | miRNA | Analysis Method | Species | Brain Area | Behavioral Parameter | |
|---|---|---|---|---|---|---|
| Upregulated | Downregulated | |||||
| Meerson et al., 2010 [232] | miR-134 | Spotted array, qRT-PCR | Adult rat | Hippocampal CA1, central nucleus of the amygdala | Chronic restraint stress | |
| Rinaldi et al., 2010 [233] | let-7a, miR-9, miR-26a/b | Microarray, Northern blot | CD1 mice | PFC | Acute and chronic restraint stress | |
| Zurawek et al., 2016 [239] | miR-16 | qRT-PCR | Male Wistar Han rat | VTA | Chronic mild stress | |
| miR-16 | medial PFC | |||||
| Higuchi et al., 2016 [241] | miR-124 | qRT-PCR, Northern blot | BALB/c mice | Hippocampus | Chronic ultra-mild stress | |
| Buran et al., 2017 [235] | miR-9-5p, miR-128-1-5p, miR-382-5p | miR-16-5p, miR-129-5p, miR-219a-5p | qRT-PCR | BALB/c mice | PFC | Chronic mild stress |
| Zurawek et al., 2017 [238] | miR-18a-5p, miR-34a-5p, miR-135a-5p, miR-195-5p, miR-320-3p, miR-674-3p, miR-872-5p | qRT-PCR | Wistar Han rat | VTA | Chronic mild stress | |
| miR-18a-5p, miR-34a-5p, miR-135a-5p, miR-195-5p, miR-320-3p, miR-674-3p, miR-872-5p | PFC | |||||
| Yu et al., 2018 [242] | miR-134 | qRT-PCR | Wistar rat | BLA | Chronic unpredictable mild stress | |
| Zhou et al., 2018 [234] | miR-382-3p, miR-183-5p, miR-3573-5p, miR-202-3p, miR-493-3p | miR-370-3p | Microarray, qRT-PCR | SPF Sprague-Dawley rat | Hippocampus | Chronic unpredictable mild stress |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.R.; Geiger, L.; Wiborg, O.; Czéh, B. Stress-Induced Morphological, Cellular and Molecular Changes in the Brain—Lessons Learned from the Chronic Mild Stress Model of Depression. Cells 2020, 9, 1026. https://doi.org/10.3390/cells9041026
Khan AR, Geiger L, Wiborg O, Czéh B. Stress-Induced Morphological, Cellular and Molecular Changes in the Brain—Lessons Learned from the Chronic Mild Stress Model of Depression. Cells. 2020; 9(4):1026. https://doi.org/10.3390/cells9041026
Chicago/Turabian StyleKhan, Ahmad Raza, Lili Geiger, Ove Wiborg, and Boldizsár Czéh. 2020. "Stress-Induced Morphological, Cellular and Molecular Changes in the Brain—Lessons Learned from the Chronic Mild Stress Model of Depression" Cells 9, no. 4: 1026. https://doi.org/10.3390/cells9041026