From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector?


{kind=link}
{kind=link}
Abstract
:1. Introduction
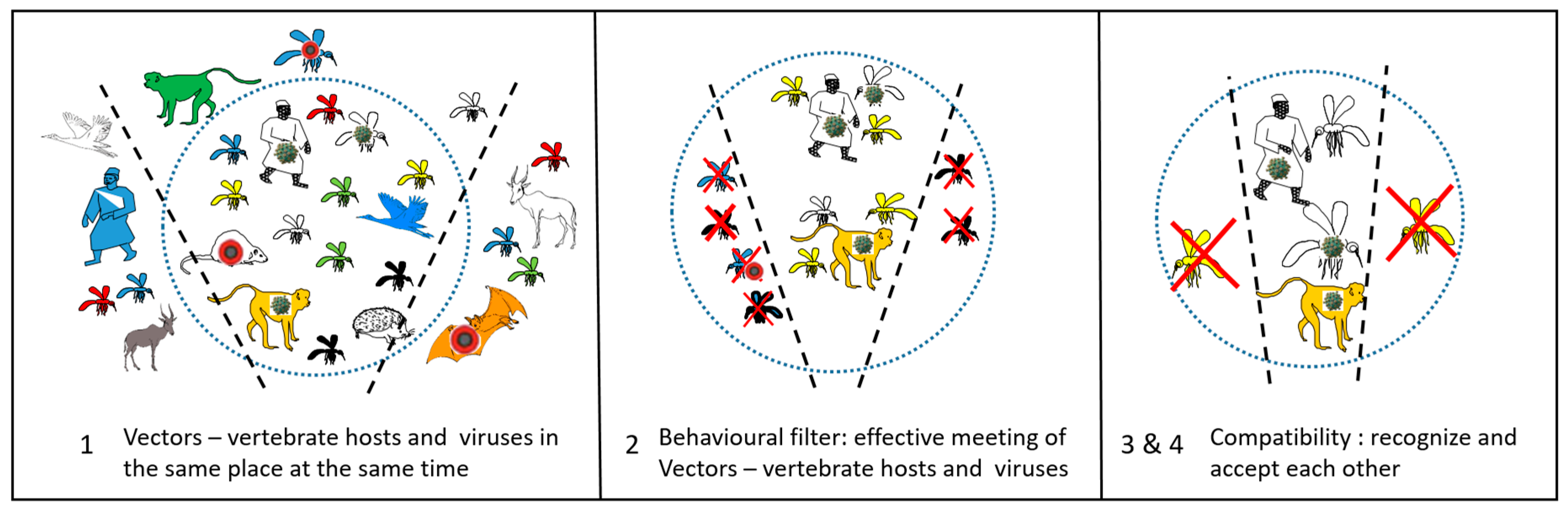
2. Vector or not yet Vector: Guilty or Presumed Innocent
3. Short History of Yellow Fever: Once Upon a Time in America
4. The Necessary Conditions
- The vector–host ratio (i.e. the vector density in relation to vertebrate host): m (the mosquito abundance);
- The human feeding rate: the number of human bites per mosquito, per day: a (mosquito - human contact);
- The daily survival rate (i.e. the probability of a mosquito surviving each day): p (mosquito longevity);
- The extrinsic development time, the time necessary for viruses to complete development from ingestion in midgut to the saliva: n;
- The infectiousness of the mosquito to the vertebrate host: b (largely dependent on virus titre in saliva)
- The susceptibility of the vertebrate host to the virus (e.g., immune state, age, health, etc): c;
- The vertebrate host infectious period: 1/r (how long the virus titre in the vertebrate remains at a level needed to infect a mosquito);
5. Aedes albopictus: From Local to Global Concern
6. Scenarios for the Future: The Worst Doesn’t Always Happen
- (1)
- Via primate-biting bridge vectors such as Ae. albopictus from forest edges in South America, Africa and Asia, forest Aedes of the Albopictus group in South East Asia, Haemagogus in South America, Stegomyia from forest galleries in Africa. In South America, Haemagogus and Sabethes are likely to transmit any new potential human viruses among primates and, as displayed by yellow fever, if these viruses are capable of transitioning to transmission in human habitats, it is likely to have already occurred.
- (2)
- By misfortune, as happened with the establishment of a sylvan cycle of yellow fever in Central and South America 500 years ago, new viruses may appear in transmission cycles that are heretofore unknown. The increase in trade and travel, and the establishment of invasive species (such as Ae. albopictus, Ae. koreicus, and Ae. japonicus, and even Ae. aegypti in Europe) suggest that this risk should not be overlooked.
- (3)
- Via increased human contacts with wild cycles due to deforestation and irrational forest exploitation. These ecological modifications favour the emergence of viruses from forest edges, and then to the human environment.
- (4)
- Via zoonotic cycles. For example, many viruses (West Nile, Japanese encephalitis, St Louis encephalitis virus, Murray valley, usutu) which are bird viruses, can be transmitted from birds (or mammals such as pigs) to humans, via vectors taking blood meals from both birds and humans, such as Cx. tritaeniorhynchus or Cx. pipiens, Cx. quinquefasciatus. It is very likely that some of these viruses, presently confined to wild cycles, will emerge in the coming years somewhere in the world, as a result of socio-ecological changes. Their shift from endemic to epidemic will be facilitated by close contact between humans and vertebrate hosts (urban commensal birds or rodents, farm animals).
- (5)
- By opening an ecological niche in urbanized areas. It is conceivable that local effective vector control by way of elimination of Ae. aegypti or Ae. albopictus would open their ecological niche in some areas, allowing a new species, such as Ae. malayensis in South-East Asia, already recognized as a vector of dengue and chikungunya viruses, to begin to colonize even closer to human populations. This scenario has not happened yet. Aedes albopictus may have replaced Ae. aegypti or vice versa, but for the moment no third species, with high vectorial capacity, has occupied their niches.
- (6)
- Via the evolution of viruses already known, but which have not yet found the conditions for emerging and spreading. Genetic changes in virus strains could lead to better adaptation to new vectors and a better transmissibility, as happened with the chikungunya virus and Ae. albopictus [52]. Viruses may also evolve resistance to drugs, when any are used, or human immune defences. Given the short generation time, large population size, and high mutation rate of RNA viruses (like yellow fever, dengue, and most pathogenic arboviruses), virus adaptation to efficient transmission by a human-preferring mosquito is rapid. That is, the virus more readily adapts to the mosquito (and vertebrate host), not the mosquito to the virus [53].
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nicolle, C. Destin des Maladies Infectieuses; Librairie Félix Alcan: Paris, France, 1933; p. 216. [Google Scholar]
- Grmek, M. Les Maladies à L’aube de la Civilisation Occidentale; PAYOT: Paris, France, 1983; p. 527. [Google Scholar]
- Weaver, S.C. Prediction and prevention of urban arbovirus epidemics: A challenge for the global virology community. Antivir. Res. 2018, 156, 80–84. [Google Scholar] [CrossRef]
- Brady, O.J.; Hay, S.I. The Global Expansion of Dengue: How Aedes aegypti Mosquitoes Enabled the First Pandemic Arbovirus. Ann. Rev. Entomol. 2020, 65, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Agramonte, A. An account of Dr. Louis-Daniel Beauperthuy, a pioneer in yellow fever research. Boston Med. Surg. J. 1908, 158, 927–930. [Google Scholar] [CrossRef] [Green Version]
- Finlay, C.J. The mosquito hypothetically considered as the agent of transmission of yellow fever. Presented at the Real Academia de Ciencias Medicas, Fısicas y Naturales de La Habana, Havana, Cuba, 14 August 1881. [Google Scholar]
- Reed, W.; Carroll, J.; Agramonte, A.; Lazear, J.W. The etiology of yellow fever: A preliminary note. Philad. Med. J. 1900, 6, 790–796. [Google Scholar]
- Christophers, S.R. Aedes aegypti (L.), the Yellow Fever Mosquito. In Its Life History, Bionomics, and Structure; Cambridge University Press: New York, NY, USA, 1960. [Google Scholar]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Hawley, A.H. The biology of Aedes albopictus. J. Am. Mosq. Control Assoc. 1988, 1, 1–39. [Google Scholar]
- Kotsakiozi, P.; Richardson, J.B.; Pichler, V.; Favia, G.; Martins, A.J.; Urbanelli, S.; Armbruster, P.A.; Caccone, A. Population genomics of the Asian tiger mosquito, Aedes albopictus: Insights into the recent worldwide invasion. Ecol. Evol. 2017, 7, 10143–10157. [Google Scholar] [CrossRef] [Green Version]
- WHO. Prevention and Control of Yellow Fever in Africa; World Health Organization: Geneva, Switzerland, 1986; p. 96. [Google Scholar]
- Powell, J.R. Mosquito-Borne Human Viral Diseases: Why Aedes aegypti? Am. J. Trop. Med. Hyg. 2018, 98, 1563–1565. [Google Scholar] [CrossRef]
- Powell, J.R.; Gloria-Soria, A.; Kotsakiozi, P. Recent history of Aedes aegypti: Vector genomics and epidemiology records. Bioscience 2018, 68, 854–860. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.; Chippaux, A. Yellow fever in Africa and the Americas: A historical and epidemiological perspective. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 20. [Google Scholar] [CrossRef] [Green Version]
- de Figueiredo, M.L.; de C Gomes, A.; Amarilla, A.A.; de S Leandro, A.; de S Orrico, A.; de Araujo, R.F.; do S M Castro, J.; Durigon, E.L.; Aquino, V.H.; Figueiredo, L.T. Mosquitoes infected with dengue viruses in Brazil. Virol. J. 2010, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.S.; Bersot, M.I.; Castro, M.G.; Telleria, E.L.; Ferreira-de-Brito, A.; Raphael, L.M.; Bonaldo, M.C.; Lourenço-de-Oliveira, R. Low vector competence in sylvatic mosquitoes limits Zika virus to initiate an enzootic cycle in South America. Sci. Rep. 2019, 9, 20151. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, L.T.M. Human Urban Arboviruses Can Infect Wild Animals and Jump to Sylvatic Maintenance Cycles in South America. Front. Cell. Infect. Microbiol. 2019, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Jácome, R.; Carrasco-Hernández, R.; Campillo-Balderas, J.A.; López-Vidal, Y.; Lazcano, A.; Wenzel, R.P.; Ponce de León, S. A yellow flag on the horizon: The looming threat of yellow fever to North America. Int. J. Infect. Dis. 2019, 87, 143–150. [Google Scholar] [CrossRef] [Green Version]
- McNeill, J.R. Mosquito Empires. In Ecology and War in the Greater Caribbean, 1620–1914; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Soper, F.L. The elimination of urban yellow fever in the Americas through eradication of Aedes aegypti. Am. J. Publ. Health 1963, 53, 7–16. [Google Scholar] [CrossRef]
- Webb, J.L., Jr. Aedes aegypti suppression in the Americas: Historical perspectives. Lancet 2016, 388, 556–557. [Google Scholar] [CrossRef]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Compos, S.S.; de Albuquerque Motta, M.; Barreto dos Santos, F.; Vazeille, M.; Vasconcelos, P.F.; Lourenco-de-Oliveira, R.; Failloux, A.B. Potential risk of re-emergence of urban transmission of yellow fever virus in Brazil facilitated by competent Aedes populations. Sci. Rep. 2017, 7, 4848. [Google Scholar] [CrossRef]
- Moreira-Soto, A.; Torres, M.C.; Lima de Mendonça, M.C.; Mares-Guia, M.A.; Dos Santos Rodrigues, C.D.; Fabri, A.A.; Dos Santos, C.C.; Machado Araújo, E.S.; Fischer, C.; Ribeiro Nogueira, R.M.; et al. Evidence for multiple sylvatic transmission cycles during the 2016–2017 yellow fever virus outbreak, Brazil. Clin. Microbiol. Infect. 2018, 24, 1019. [Google Scholar] [CrossRef] [Green Version]
- Lemos, P.S.; Monteiro, H.A.O.; Castro, F.C.; Lima, C.P.S.; Silva, D.E.A.; Vasconcelos, J.M.; Oliveira, L.F.; Silva, S.P.; Cardoso, J.F.; Vianez-Júnior, J.L.S.G.; et al. Caracterização do genoma mitocondrial de Haemagogus janthinomys (Diptera: Culicidae). DNA Mitocondrial 2017, 28, 50–51. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Wallis, G.P.; Aitken, T.H.G.; Miller, B.R.; Amato, G.D.; Lorenz, L.; Powell, J.R.; Beaty, B.R. Oral infection of Aedes aegypti with yellow fever virus: Geographical variation and genetic considerations. Am. J. Trop. Med. Hyg. 1985, 34, 1219–1224. [Google Scholar] [CrossRef]
- Wasserman, S.; Tambyah, P.A.; Lim, P.L. Yellow fever cases in Asia: Primed for an epidemic. Int. J. Infect. Dis. 2016, 48, 98–103. [Google Scholar] [CrossRef] [Green Version]
- Brey, P.T.; Fontenille, D.; Tang, H. Re-evaluate yellow fever risk in Asia-Pacific region. Nature 2018, 554, 31. [Google Scholar] [CrossRef]
- Macdonald, G. The Epidemiology and Control of Malaria; Oxford University Press: Oxford, UK, 1957. [Google Scholar]
- Cohuet, A.; Harris, C.; Robert, V.; Fontenille, D. Evolutionary forces on Anopheles: What makes a malaria vector? Trends Parasitol. 2010, 26, 130–136. [Google Scholar] [CrossRef]
- Azar, S.R.; Weaver, S.C. Vector competence: What has Zika virus taught us. Viruses 2019, 11, 867. [Google Scholar] [CrossRef] [Green Version]
- Houé, V.; Gabiane, G.; Dauga, C.; Suez, M.; Madec, Y.; Mousson, L.; Marconcini, M.; Yen, P.S.; de Lamballerie, X.; Bonizzoni, M.; et al. Evolution and biological significance of flaviviral elements in the genome of the arboviral vector Aedes albopictus. Emerg. Microbes Infect. 2019, 8, 1265–1279. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.R. An update on mosquito cell expressed dengue virus receptor proteins. Insect Mol. Biol. 2012, 21, 1–7. [Google Scholar] [CrossRef]
- Combes, C. Parasitism: The Ecology and Evolution of Intimate Interactions; University of Chicago Press: Chicago, IL, USA, 2001; p. 728. [Google Scholar]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes albopictusas an epidemic vector of Chikungunya virus: Another emerging problem? Lancet Inf. Dis. 2006, 6, 463–464. [Google Scholar] [CrossRef]
- Delatte, H.; Desvars, A.; Bouetard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, vector of chikungunya on La Reunion. Vector Borne Zoonotic Dis. 2008, 8, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.M. Contributions to the mosquito fauna of Southeast Asia. XIV. The subgenus Stegomyia of Aedes in Southeast Asia. I. The scuterallis group of species. Contrib. Am. Entomol. Inst. 1972, 9, 1–109. [Google Scholar]
- Mogi, M.; Armbruster, P.A.; Tuno, N.; Aranda, C.; Yong, H.S. The climate range expansion of Aedes albopictus (Diptera: Culicidae) in Asia inferred from the distribution of Albopictus subgroup species of Aedes (Stegomyia). J. Med. Entomol. 2017, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Miot, E. Potential of the mosquito Aedes malayensis as an arbovirus vector in South East Asia. Ph.D. Thesis, Sorbonne University, Paris, France, 20 December 2019. [Google Scholar]
- Tangena, J.A.; Thammavong, P.; Malaithong, N.; Inthavong, T.; Ouanesamon, P.; Brey, P.T.; Lindsay, S.W. Diversity of mosquitoes (Diptera: Culicidae) attracted to human subjects in rubber plantations, secondary forests, and villages in Luang Prabang Province, northern Lao PDR. J. Med. Entomol. 2017, 54, 1589–1604. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Wasserman, R.J.; Gan, J.Y.; Wilson, R.F.; Rahman, F.; Yek, S.H. Human activities attract harmful mosquitoes in a tropical urban landscape. EcoHealth 2020, 17, 52–63. [Google Scholar] [CrossRef]
- Pereira Dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef] [Green Version]
- Lounibos, L.P.H. Invasions by Insect Vectors of Human Disease. Ann. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef]
- Lounibos, L.P. Habitat segregation among African treehole mosquitoes. Ecol. Entomol. 1981, 6, 129–154. [Google Scholar] [CrossRef]
- Delatte, H.; Bagny, L.; Brengue, C.; Bouetard, A.; Paupy, C.; Fontenille, D. The invaders: Phylogeography of dengue and chikungunya viruses Aedes vectors, on the South West islands of the Indian Ocean. Infect. Genet. Evol. 2011, 11, 1769–1781. [Google Scholar] [CrossRef]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef]
- Hadfield, J.; Brito, A.F.; Swetnam, D.M.; Vogels, C.B.F.; Tokarz, R.E.; Andersen, K.G.; Smith, R.C.; Bedford, T.; Grubaugh, N.D. Twenty years of West Nile virus spread and evolution in the Americas visualized by Nextstrain. PLoS Pathog. 2019, 15, e1008042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, F.A.; Calisher, C.H.; Tesh, R.B.; Walker, D.H. In memoriam: Robert Ellis Shope (1929–2004). Emerg. Infect. Dis. 2004, 10, 762–765. [Google Scholar] [CrossRef]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome Microevolution of Chikungunya Viruses Causing the Indian Ocean Outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.R. An evolutionary perspective on vector-borne diseases. Front. Gent. 2019, 10, 1266. [Google Scholar] [CrossRef] [PubMed]
- Abad-Franch, F.; Grimmer, G.H.; de Paula, V.S.; Figueiredo, L.T.; Braga, W.S.M.; Luz, S.L.B. Mayaro Virus Infection in Amazonia: A Multimodel Inference Approach to Risk Factor Assessment. PLoS Negl. Trop. Dis. 2012, 6, e1846. [Google Scholar] [CrossRef] [PubMed]
- Pezzi, L.; A Diallo, M.; Rosa-Freitas, M.G.; Vega-Rua, A.; Ng, L.F.P.; Boyer, S.; Drexler, J.F.; Vasilakis, N.; Lourenço-de-Oliveira, R.; Weaver, S.C.; et al. GloPID-R report on chikungunya, o’nyong-nyong and Mayaro virus, part 5: Entomological aspects. Antivir. Res. 2020, 174, 104670. [Google Scholar] [CrossRef] [PubMed]
- Haddow, A.D.; Nasar, F.; Guzman, H.; Ponlawat, A.; Jarman, R.G.; Tesh, R.B.; Weaver, S.C. Genetic Characterization of Spondweni and Zika Viruses and Susceptibility of Geographically Distinct Strains of Aedes aegypti, Aedes albopictus and Culex quinquefasciatus (Diptera: Culicidae) to Spondweni Virus. PLoS Negl. Trop. Dis. 2016, 10, e0005083. [Google Scholar] [CrossRef] [Green Version]
- White, S.K.; Lednicky, J.A.; Okech, B.A.; Morris, J.G., Jr.; Dunford, J.C. Spondweni Virus in Field-Caught Culex quinquefasciatus Mosquitoes, Haiti, 2016. Emerg. Infect. Dis. 2018, 24, 1765–1767. [Google Scholar] [CrossRef] [Green Version]
- WHO. International Health Regulations, 3rd ed.; World Health Organization: Geneva, Switzerland, 2005; p. 74. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontenille, D.; Powell, J.R. From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector? Pathogens 2020, 9, 265. https://doi.org/10.3390/pathogens9040265
Fontenille D, Powell JR. From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector? Pathogens. 2020; 9(4):265. https://doi.org/10.3390/pathogens9040265
Chicago/Turabian StyleFontenille, Didier, and Jeffrey R. Powell. 2020. "From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector?" Pathogens 9, no. 4: 265. https://doi.org/10.3390/pathogens9040265
APA StyleFontenille, D., & Powell, J. R. (2020). From Anonymous to Public Enemy: How Does a Mosquito Become a Feared Arbovirus Vector? Pathogens, 9(4), 265. https://doi.org/10.3390/pathogens9040265