Collagen Peptide Upregulates Osteoblastogenesis from Bone Marrow Mesenchymal Stem Cells through MAPK- Runx2



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
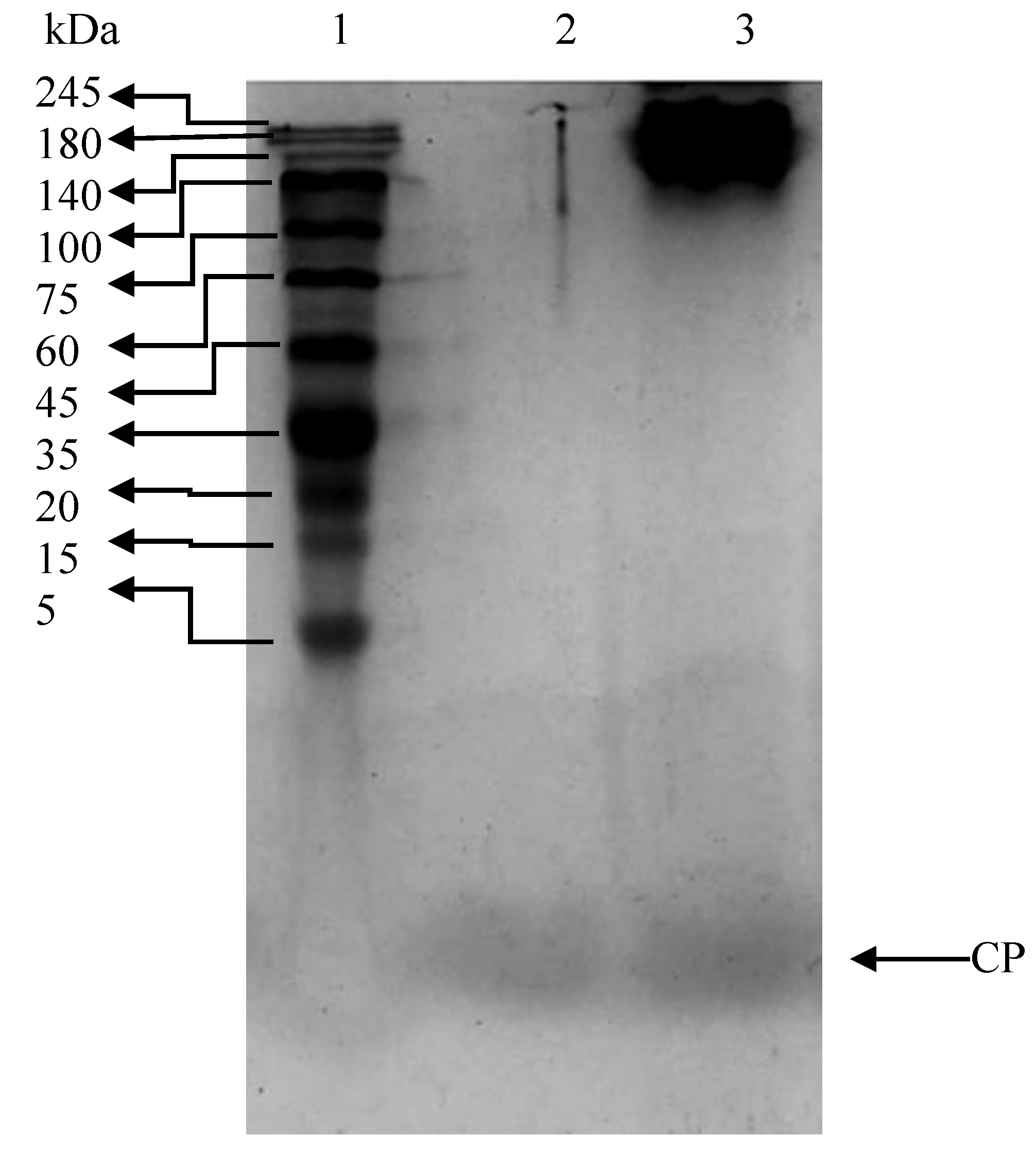
2.1. Molecular Weight Analysis
2.2. Effect of CP on the Proliferation of BMMS Cells
2.3. Effect of CP on the Differentiation of BMMS Cells
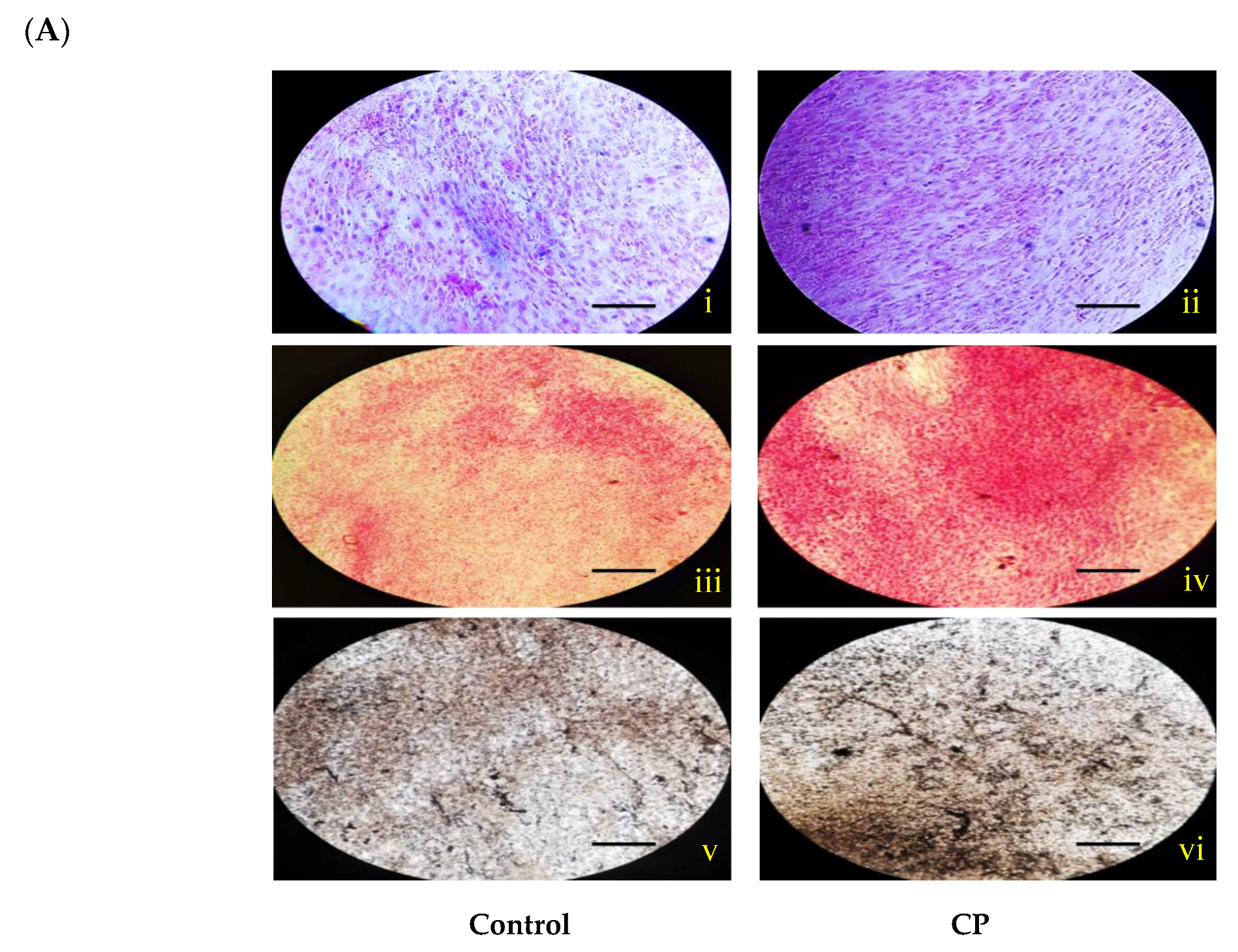
2.4. Histological Staining
2.5. Immunocytochemistry
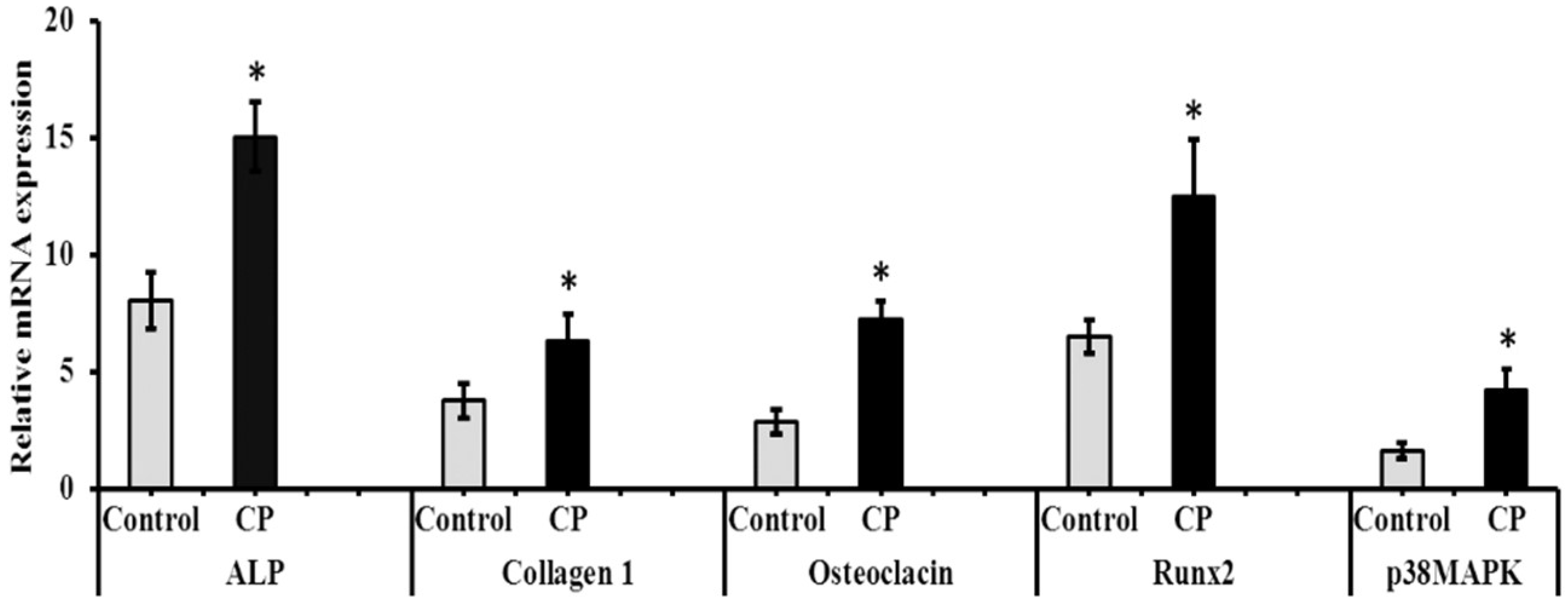
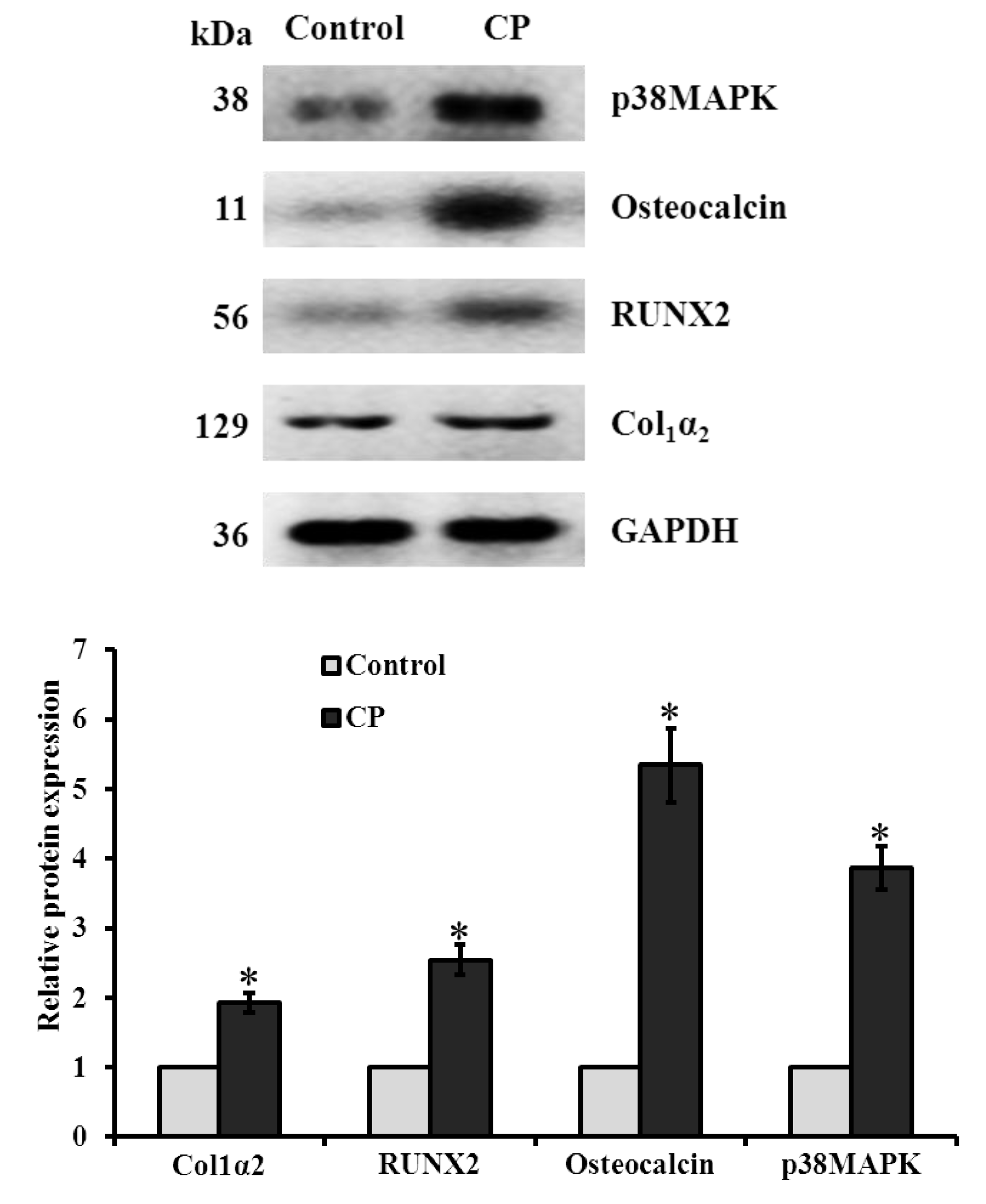
2.6. mRNA and Protein Expression of CP Treated BMMS Cells
3. Materials and Methods
3.1. Extraction of Fish Bone Collagen Peptide (CP)
3.2. Molecular Mass by Tricine SDS-PAGE
3.3. Cell Culture
3.4. Effect of CP on BMMS Cells Proliferation
3.5. Effect of CP on BMMS Cells Differentiation
3.6. Histological Staining for Osteogenic Evaluation
3.7. Cellular Alkaline Phosphatase
3.8. Immunocytochemistry
3.9. mRNA Expression
3.9.1. RNA Extractions
3.9.2. cDNA Synthesis and RT-PCR
3.10. Western Blot
3.11. Statistics
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (Hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Nagai, T.; Suzuki, N. Isolation of collagen from fish waste material - skin, bone and fins. Food Chem. 2000, 68, 277–281. [Google Scholar] [CrossRef]
- Muralidharan, N.; Jeya Shakila, R.; Sukumar, D.; Jeyasekaran, G. Skin, bone and muscle collagen extraction from the trash fish, leather jacket (Odonus niger) and their characterization. J. Food Sci. Technol. 2013, 50, 1106–1113. [Google Scholar] [CrossRef]
- Jeevithan, E.; Bao, B.; Bu, Y.; Zhou, Y.; Zhao, Q.; Wu, W.H. Type II collagen and gelatin from silvertip shark (Carcharhinus albimarginatus) cartilage: Isolation purification physicochemical and antioxidant properties. Mar. Drug 2014, 12, 3852–3873. [Google Scholar] [CrossRef] [PubMed]
- Jongjareonrak, A.; Benjakul, S.; Visessanguan, W.; Nagai, T.; Tanaka, M. Isolation and characterisation of acid and pepsin-solubilised collagens from the skin of brownstripe red snapper (Lutjanus vitta). Food Chem. 2005, 93, 475–484. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Jeevithan, E.; Zhao, Q.; Bin, B.; Wu, W. Biomedical and pharmaceutical application of fish collagen and gelatin: A Review. J. Nutr. Therapeut. 2013, 2, 218–227. [Google Scholar]
- Liu, D.; Nikoo, M.; Boran, G.; Zhou, P.; Regenstein, J.M. Collagen and gelatin. Ann. Rev. Food Sci. Technol. 2015, 6, 527–557. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Bao, B.; Wang, N.; Xie, J.; Wu, W.H. Oral administration of shark type II collagen suppresses complete freund’s adjuvant-induced rheumatoid arthritis in rats. Pharmaceutical 2012, 5, 339–352. [Google Scholar] [CrossRef]
- Xi, C.; Tan, L.; Sun, Y.; Liang, F.; Liu, N.; Xue, H.; Luo, Y.; Yuan, F.; Sun, Y.; Xi, Y. A novel recombinant peptide containing only two T-cell tolerance epitopes of chicken type-II collagen that suppresses collagen-induced arthritis. Mol. Immunol. 2009, 46, 729–737. [Google Scholar] [CrossRef]
- Schmid, T.M.; Conrad, H.E. A unique low molecular weight collagen secreted by cultured chick embryo chondrocytes. J. Biol. Chem. 1982, 257, 12444–12450. [Google Scholar] [PubMed]
- Myers, L.K.; Sakurai, Y.; Rosloniec, E.F.; Stuart, J.M.; Kang, A.H. An analog peptide that suppresses collagen-induced arthritis. Am. J. Med. Sci. 2004, 327, 212–216. [Google Scholar] [CrossRef]
- Ehrlich, H. Cartilage of marine vertebrates. Biol. Mater Marine Orig. 2015, 4, 69–89. [Google Scholar]
- Xie, J.; Ye, H.Y.; Luo, X.F. An efficient preparation of chondroitin sulfate and collagen peptides from shark cartilage. Int. Food Res. J. 2014, 21, 1171–1175. [Google Scholar]
- Jeevithan, E.; Bao, B.; Zhang, J.; Hong, S.; Wu, W. Purification, characterization and antioxidant properties of low molecular weight collagenous polypeptide (37 kDa) prepared from whale shark cartilage (Rhincodon typus). J. Food Sci. Technol. 2015, 52, 6312–6322. [Google Scholar] [CrossRef]
- Wang, J.; Luo, D.; Liang, M.; Zhang, T.; Yin, X.; Zhang, Y.; Yang, X.; Liu, W. Spectrum-effect relationships between high-performance liquid chromatography (HPLC) fingerprints and the antioxidant and anti-inflammatory activities of collagen peptides. Molecule 2018, 23, 3257. [Google Scholar] [CrossRef]
- Chen, Q.; Gao, X.; Zhang, H.; Li, B.; Yu, G.; Li, B. Collagen peptides administration in early enteral nutrition intervention attenuates burn-induced intestinal barrier disruption: Effects on tight junction structure. J. Funct. Food. 2019, 55, 167–174. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. p38 MAPK signaling in osteoblast differentiation. Front Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef]
- Xu, X.; Han, J.; Ito, Y.; Bringas, P.; Deng, C.; Chai, Y. Ectodermal Smad4 and p38 MAPK are functionally redundant in mediating TGF-beta/BMP signaling during tooth and palate development. Dev. Cell 2008, 15, 322–329. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Diomede, F.; Merciaro, I.; Caputi, S.; Tartaro, A.; Guarnieri, S.; Trubiani, O. Endothelial committed oral stem cells as modelling in the relationship between periodontal and cardiovascular disease. J. Cell Physiol. 2018, 233, 6734–6747. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed]
- Sciandra, M.; Marino, M.T.; Manara, M.C.; Guerzoni, C.; Grano, M.; Oranger, A.; Lucarelli, E.; Lollini, P.-L.; Dozza, B.; Pratelli, L.; et al. CD99 drives terminal differentiation of osteosarcoma cells by acting as a spatial regulator of ERK 1/2. J. Bone Miner Res. 2014, 29, 1295–1309. [Google Scholar] [CrossRef]
- Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Franceschi, R.T. Fibroblast growth factor 2 induction of the osteocalcin gene requires MAPK activity and phosphorylation of the osteoblast transcription factor, Cbfa1/Runx2. J. Biol. Chem. 2002, 277, 36181–36187. [Google Scholar] [CrossRef]
- Cuadrado, A.; Nebreda, A.R. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar] [CrossRef]
- Brancho, D.; Tanaka, N.; Jaeschke, A.; Ventura, J.J.; Kelkar, N.; Tanaka, Y.; Kyuuma, M.; Takeshita, T.; Flavell, R.A.; Davis, R.J. Mechanism of p38 MAP kinase activation in vivo. Genes Dev. 2003, 17, 1969–1978. [Google Scholar] [CrossRef]
- Schagger, H. Tricine-SDS-PAGE. Nature 2006, 1, 16–22. [Google Scholar] [CrossRef]
- Donzelli, E.; Salvadè, A.; Mimo, P.; Viganò, M.; Morrone, M.; Papagna, R.; Carini, F.; Zaopo, A.; Miloso, M.; Baldoni, M.; et al. Mesenchymal stem cells cultured on a collagen scaffold: In vitro osteogenic differentiation. Arch Oral. Biol. 2007, 52, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yin, S.; Xue, X.; Ji, J.; Tong, J.; Goltzman, D.; Miao, D. Transplantation of bone marrow-derived mesenchymal stem cells rescues partially rachitic phenotypes induced by 1, 25-Dihydroxyvitamin D deficiency in mice. Am. J. Transl. Res. 2016, 8, 4382–4393. [Google Scholar] [PubMed]
- Jeevithan, E.; Sanchez, C.; de Val, J.M.E.S.; Henrotin, Y.; Wang, S.; Motaung, K.S.C.M.; Regenstein, J.M.; Bao, B.; Wu, W.H. Cross-talk between primary osteocytes and bone marrow macrophages for osteoclastogenesis upon collagen treatment. Sci. Rep. 2018, 8, 5318. [Google Scholar]
- Kim, H.K.; Kim, M.G.; Leem, K.H. Osteogenic activity of collagen peptide via ERK/MAPK pathway mediated boosting of collagen synthesis and its therapeutic efficacy in osteoporotic bone by back-scattered electron imaging and microarchitecture analysis. Molecule 2013, 18, 15474–15489. [Google Scholar] [CrossRef]
- Guillerminet, F.; Beaupied, H.; Fabien-Soule, V.; Tome, D.; Benhamou, C.L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone metabolism and biomechanical parameters in ovariectomized mice: An in vitro and in vivo study. Bone 2010, 46, 827–834. [Google Scholar] [CrossRef]
- Daneault, A.; Coxam, V.; Fabien Soulé, V.; Wittrant, Y. Hydrolyzed collagen contributes to osteoblast differentiation in vitro and subsequent bone health in vivo. Presented at the OARSI World Congress, Paris, France, 24–27 April 2014. [Google Scholar]
- Fathi-Najafi, M.; Vahedi, F.; Ahmadi, S.; Madani, R.; Mehrvarz, M. Effect of collagen type I (Rat tail) on cell proliferation and adhesion of BHK-21. 4th Kuala Lumpur. Int. Conf. Biomed. Eng. 2008, 806–809. [Google Scholar]
- Gao, C.; Harvey, E.J.; Chua, M.; Chen, B.P.; Jiang, F.; Liu, Y.; Li, A.; Wang, H.; Henderson, J.E. MSC-seeded dense collagen scaffolds with a bolus dose of VEGF promote healing of large bone defects. Euro. Cells Mat. 2013, 26, 195–207. [Google Scholar] [CrossRef]
- Hennessy, K.M.; Pollot, B.E.; Clem, W.C.; Phipps, M.C.; Sawyer, A.A.; Culpepper, K.; Bellis, S.L. The effect of collagen I mimetic peptides on mesenchymal stem cell adhesion and differentiation, and on bone formation at hydroxyapatite surfaces. Biomaterial 2009, 30, 1898–1909. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.H.; Lai, W.F.; Chang, S.F.; Wong, C.C.; Fan, C.Y.; Fang, C.L. The effect of type II collagen on MSC osteogenic differentiation and bone defect repair. Biomaterial 2014, 35, 2680–2691. [Google Scholar] [CrossRef]
- Daneault, A.; Prawitt, J.; Fabien Soulé, V.; Coxam, V.; Wittrant, Y. Biological effect of hydrolyzed collagen on bone metabolism. Crit. Rev. Food Sci. Nutr. 2017, 57, 1922–1937. [Google Scholar] [CrossRef]
- Bernhardt, A.; Lode, A.; Boxberger, S.; Pompe, W.; Gelinsky, M. Mineralised collagen-An artificial, extracellular bone matrix—Improves osteogenic differentiation of bone marrow stromal cells. J. M. Sci. Mat. Med. 2008, 19, 269–275. [Google Scholar] [CrossRef]
- Tsuruoka, N.; Yamato, R.; Sakai, Y.; Yoshitake, Y.; Yonekura, H. Promotion by collagen tripeptide of type I collagen gene expression in human osteoblastic cells and fracture healing of rat femur. Biosci. Biotechnol. Biochem. 2007, 71, 2680–2687. [Google Scholar] [CrossRef]
- Boskey, A.L.; Robey, P.G. Chapter 11—The regulatory role of matrix proteins in mineralization of bone. Osteopores (Fourth Edition) 2013, 235–255. [Google Scholar] [CrossRef]
- Mizuno, M.; Kuboki, Y. Osteoblast-related gene expression of bone marrow cells during the osteoblastic differentiation induced by type I collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef]
- Jeevithan, E.; Jung, W.L.; Shujun, W.; Yves, H.; José, E.M.S.V.; Regenstein, J.M.; Bao, B.; Wu, W.H. Evaluation of differentiated bone cells proliferation by blue shark skin collagen via biochemical for bone tissue engineering. Mar. Drug 2018, 16, 350. [Google Scholar]
- Mizuno, M.; Fujisawa, R.; Kuboki, Y. Type I collagen-induced osteoblastic differentiation of bone-marrow cells mediated by collagen-alpha2beta1 integrin interaction. J. Cell Physiol. 2000, 184, 207–213. [Google Scholar] [CrossRef]
- Xu, Y.; Han, X.; Li, Y. Effect of marine collagen peptides on long bone development in growing rats. J. Sci. Food Agric. 2010, 90, 1485–1491. [Google Scholar] [CrossRef]
- Wu, J.; Fujioka, M.; Sugimoto, K.; Mu, G.; Ishimi, Y. Assessment of effectiveness of oral administration of collagen peptide on bone metabolism in growing and mature rats. J. Bone Miner Metab. 2004, 22, 547–553. [Google Scholar] [CrossRef]
- Takeda, S.; Park, J.H.; Kawashima, E.; Ezawa, I.; Omi, N. Hydrolyzed collagen intake increases bone mass of growing rats trained with running exercise. J. Int. Soc. Sports Nutr. 2013, 10, 35. [Google Scholar] [CrossRef]
- Watanabe-Kamiyama, M.; Shimizu, M.; Kamiyama, S.; Taguchi, Y.; Sone, H.; Morimatsu, F.; Komai, M. Absorption and effectiveness of orally administered low molecular weight collagen hydrolysate in rats. J. Agric. Food Chem. 2010, 58, 835–841. [Google Scholar] [CrossRef]
- Guillerminet, F.; Fabien-Soule, V.; Even, P.C.; Tome, D.; Benhamou, C.L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone status and prevents bone loss in ovariectomized C3H/HeN mice. Osteoporos Int. 2012, 23, 1909–1919. [Google Scholar] [CrossRef]
- Tuckwell, D.S.; Ayad, S.; Grant, M.E.; Takigawa, M.; Humphries, M.J. Conformation dependence of integrin-type II collagen binding Inability of collagen peptides to support alpha 2 beta 1 binding, and mediation of adhesion to denatured collagen by a novel alpha 5 beta 1-fibronectin bridge. J. Cell Sci. 1994, 107, 993–1005. [Google Scholar]
- Zhang, Z.; Zhou, S.; Mei, Z.; Zhang, M. Inhibition of p38MAPK potentiates mesenchymal stem cell therapy against myocardial infarction injury in rats. Mol. Med. Rep. 2017, 16, 3489–3493. [Google Scholar] [CrossRef][Green Version]
- Kawaguchi, T.; Nanbu, P.N.; Kurokawa, M. Distribution of prolylhydroxyproline and its metabolites after oral administration in rats. Biol. Pharm. Bull. 2012, 35, 422–427. [Google Scholar] [CrossRef]
- Kimira, Y.; Odaira, H.; Nomura, K.; Taniuchi, Y.; Inoue, N.; Nakatani, S.; Shimizu, J.; Wada, M.; Mano, H. Collagen-derived dipeptide prolylhydroxyproline promotes osteogenic differentiation through Foxg1. Cell Mol. Biol. Lett. 2017, 22, 27. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elango, J.; Robinson, J.; Zhang, J.; Bao, B.; Ma, N.; de Val, J.E.M.S.; Wu, W. Collagen Peptide Upregulates Osteoblastogenesis from Bone Marrow Mesenchymal Stem Cells through MAPK- Runx2. Cells 2019, 8, 446. https://doi.org/10.3390/cells8050446
Elango J, Robinson J, Zhang J, Bao B, Ma N, de Val JEMS, Wu W. Collagen Peptide Upregulates Osteoblastogenesis from Bone Marrow Mesenchymal Stem Cells through MAPK- Runx2. Cells. 2019; 8(5):446. https://doi.org/10.3390/cells8050446
Chicago/Turabian StyleElango, Jeevithan, Jeyashakila Robinson, Jingyi Zhang, Bin Bao, Nan Ma, José Eduardo Maté Sánchez de Val, and Wenhui Wu. 2019. "Collagen Peptide Upregulates Osteoblastogenesis from Bone Marrow Mesenchymal Stem Cells through MAPK- Runx2" Cells 8, no. 5: 446. https://doi.org/10.3390/cells8050446
APA StyleElango, J., Robinson, J., Zhang, J., Bao, B., Ma, N., de Val, J. E. M. S., & Wu, W. (2019). Collagen Peptide Upregulates Osteoblastogenesis from Bone Marrow Mesenchymal Stem Cells through MAPK- Runx2. Cells, 8(5), 446. https://doi.org/10.3390/cells8050446