
Effects of Preceding Anesthesia Protocols on Insulin and Glucagon Secretion from Isolated Perfused Rat Pancreas Preparations

 , ,
, ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Anesthetic Protocols
2.3. Assessment of Surgical Depth of Anesthesia
2.4. Isolated Perfused Rat Pancreas
2.5. Experimental Protocol
- (1)
- Glucose variations: The pancreas was exposed to varying glucose concentrations ranging from low (1.5 or 3.5 mM), intermediate (6.0 mM) or high (8.0 or 10 mM) in the perfusion buffer. Perfusion with each concentration lasted 15–20 min.
- (2)
- Repeated stimulations with GLP-1: During pancreas perfusion with a perfusion buffer containing 6.0 mM glucose, three stimulations with 1 nM GLP-1 (7-36) were performed. Each lasted 10 min and they were separated by a washout period of 10 or 40 min.
2.6. Test Substances
2.7. Hormone Analysis
3. Results
3.1. Assessment of Anesthetic Adequacy for Surgical General Anesthesia
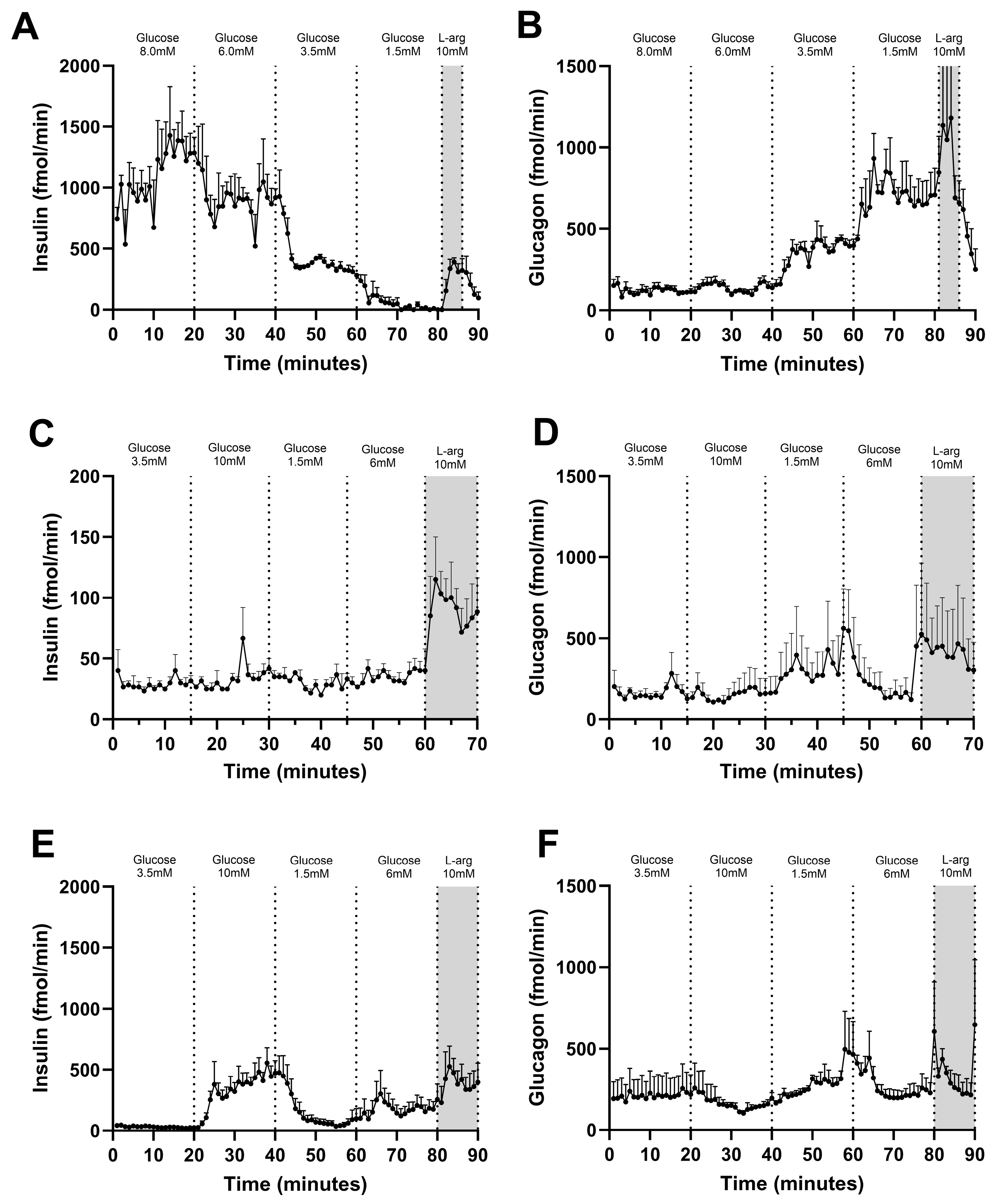
3.2. Glucose-Stimulated Insulin and Glucagon Secretion
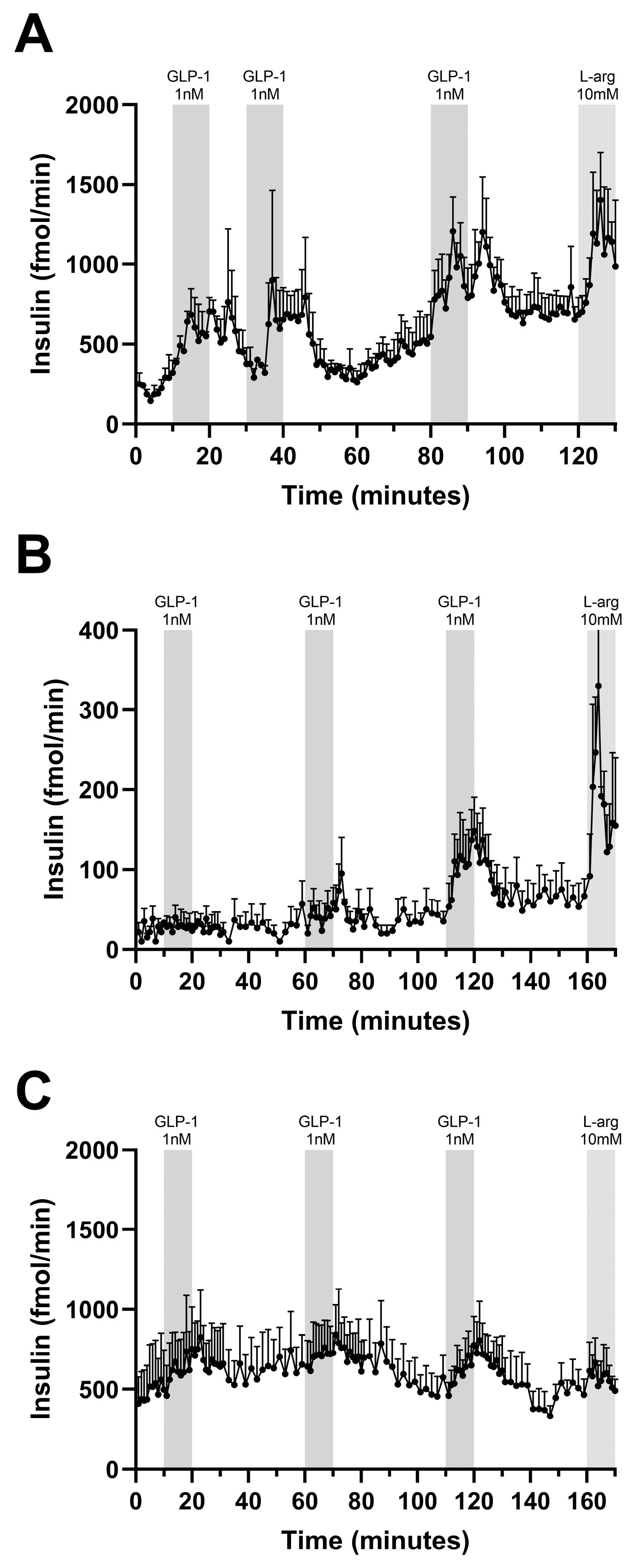
3.3. Incretin-Stimulated Insulin Secretion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GLP-1 | Glucagon-like peptide-1 |
| GSIS | Glucose-induced insulin secretion |
| NMDA | N-methyl-D-aspartate |
| FELASA | Federation of Laboratory Animal Science Associations |
| ACF | Animal Core Facility |
| RIA | Radioimmunoassay |
| cAMP | Cyclic adenosine monophosphate |
References
- Brown, L.; Panchal, S.K. Rodent Models for Metabolic Syndrome Research. J. Biomed. Biotechnol. 2010, 2011, 351982. [Google Scholar] [CrossRef]
- Ojha, A.; Ojha, U.; Mohammed, R.; Chandrashekar, A.; Ojha, H. Current Perspective on the Role of Insulin and Glucagon in the Pathogenesis and Treatment of Type 2 Diabetes Mellitus. Clin. Pharmacol. Adv. Appl. 2019, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Frías, J.P.; Davies, M.J.; Rosenstock, J.; Pérez Manghi, F.C.; Fernández Landó, L.; Bergman, B.K.; Liu, B.; Cui, X.; Brown, K. Tirzepatide versus Semaglutide Once Weekly in Patients with Type 2 Diabetes. N. Engl. J. Med. 2021, 385, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, A.M.; Aronne, L.J.; Ahmad, N.N.; Wharton, S.; Connery, L.; Alves, B.; Kiyosue, A.; Zhang, S.; Liu, B.; Bunck, M.C.; et al. Tirzepatide Once Weekly for the Treatment of Obesity. N. Engl. J. Med. 2022, 387, 205–216. [Google Scholar] [CrossRef]
- Holst, J.J. GLP-1 Physiology in Obesity and Development of Incretin-Based Drugs for Chronic Weight Management. Nat. Metab. 2024, 6, 1866–1885. [Google Scholar] [CrossRef]
- Agerskov, R.H.; Nyeng, P. Innervation of the Pancreas in Development and Disease. Development 2024, 151, dev202254. [Google Scholar] [CrossRef]
- Chmelir, T.; Jarkovska, D.; Pandey, S.; Chottova Dvorakova, M. Neurogastroenterology: Current Insights into Gastrointestinal Innervation in Health and Disease. Auton. Neurosci. 2025, 261, 103339. [Google Scholar] [CrossRef]
- Hendrickx, J.F.A.; Eger, E.I.; Sonner, J.M.; Shafer, S.L. Is Synergy the Rule? A Review of Anesthetic Interactions Producing Hypnosis and Immobility. Anesth. Analg. 2008, 107, 494–506. [Google Scholar] [CrossRef]
- Johansen, O.; Vaaler, S.; Jorde, R.; Reikerås, O. Increased Plasma Glucose Levels after Hypnorm® Anaesthesia, but Not after Pentobarbital® Anaesthesia in Rats. Lab. Anim. 1994, 28, 244–248. [Google Scholar] [CrossRef]
- Lattermann, R.; Schricker, T.; Wachter, U.; Georgieff, M.; Goertz, A. Understanding the Mechanisms by Which Isoflurane Modifies the Hyperglycemic Response to Surgery. Anesth. Analg. 2001, 93, 121–127. [Google Scholar] [CrossRef]
- Saha, J.K.; Xia, J.; Grondin, J.M.; Engle, S.K.; Jakubowski, J.A. Acute Hyperglycemia Induced by Ketamine/Xylazine Anesthesia in Rats: Mechanisms and Implications for Preclinical Models. Exp. Biol. Med. 2005, 230, 777–784. [Google Scholar] [CrossRef]
- Tanaka, T.; Nabatame, H.; Tanifuji, Y. Insulin Secretion and Glucose Utilization Are Impaired under General Anesthesia with Sevoflurane as Well as Isoflurane in a Concentration-Independent Manner. J. Anesth. 2005, 19, 277–281. [Google Scholar] [CrossRef]
- Zuurbier, C.J.; Keijzers, P.J.M.; Koeman, A.; Van Wezel, H.B.; Hollmann, M.W. Anesthesia’s Effects on Plasma Glucose and Insulin and Cardiac Hexokinase at Similar Hemodynamics and without Major Surgical Stress in Fed Rats. Anesth. Analg. 2008, 106, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yamato Kodera, S.; Yoshida, M.; Dezaki, K.; Yada, T.; Murayama, T.; Kawakami, M.; Kakei, M. Inhibition of Insulin Secretion from Rat Pancreatic Islets by Dexmedetomidine and Medetomidine, Two Sedatives Frequently Used in Clinical Settings. Endocr. J. 2013, 60, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Kawano, T.; Eguchi, S.; Chi, H.; Iwata, H.; Yokoyama, M. Effects of Dexmedetomidine on Insulin Secretion from Rat Pancreatic β Cells. J. Anesth. 2014, 29, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Windeløv, J.A.; Pedersen, J.; Holst, J.J. Use of Anesthesia Dramatically Alters the Oral Glucose Tolerance and Insulin Secretion in C57Bl/6 Mice. Physiol. Rep. 2016, 4, e12824. [Google Scholar] [CrossRef]
- Bouillon, J.; Duke, T.; Focken, A.P.; Snead, E.C.; Cosford, K.L. Effects of Dexmedetomidine on Glucose Homeostasis in Healthy Cats. J. Feline Med. Surg. 2019, 22, 344. [Google Scholar] [CrossRef]
- Murphy, K.L.; Roughan, J.V.; Baxter, M.G.; Flecknell, P.A. Anaesthesia with a Combination of Ketamine and Medetomidine in the Rabbit: Effect of Premedication with Buprenorphine. Vet. Anaesth. Analg. 2010, 37, 222–229. [Google Scholar] [CrossRef]
- LaTourette, P.C.; David, E.M.; Pacharinsak, C.; Jampachaisri, K.; Smith, J.C.; Marx, J.O. Effects of Standard and Sustained-Release Buprenorphine on the Minimum Alveolar Concentration of Isoflurane in C57BL/6 Mice. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2020, 59, 298. [Google Scholar] [CrossRef]
- Pathan, H.; Williams, J. Basic Opioid Pharmacology: An Update. Br. J. Pain 2012, 6, 11. [Google Scholar] [CrossRef]
- Flecknell, P.A.; Mitchell, M. Midazolam and Fentanyl-Fluanisone: Assessment of Anaesthetic Effects in Laboratory Rodents and Rabbits. Lab. Anim. 1984, 18, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Tremoleda, J.L.; Kerton, A.; Gsell, W. Anaesthesia and Physiological Monitoring During In Vivo Imaging of Laboratory Rodents: Considerations on Experimental Outcomes and Animal Welfare. EJNMMI Res. 2012, 2, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Flecknell, P.; Lofgren, J.L.S.; Dyson, M.C.; Marini, R.R.; Michael Swindle, M.; Wilson, R.P. Preanesthesia, Anesthesia, Analgesia, and Euthanasia. In Laboratory Animal Medicine: Third Edition; Academic Press: Cambridge, MA, USA, 2015; pp. 1135–1200. [Google Scholar] [CrossRef]
- Winters, W.D.; Ferrar-Allado, T.; Guzman-Flores, C.; Alcaraz, M. The Cataleptic State Induced by Ketamine: A Review of the Neuropharmacology of Anesthesia. Neuropharmacology 1972, 11, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, L.F.; Alegado, D.B.; Howard, E.B. Ketamine and Xylazine for Surgical Anesthesia in Rats. J. Am. Vet. Med. Assoc. 1977, 171, 842–844. [Google Scholar] [CrossRef]
- Green, C.J.; Knight, J.; Precious, S.; Simpkin, S. Ketamine Alone and Combined with Diazepam or Xylazine in Laboratory Animals: A 10 Year Experience. Lab. Anim. 1981, 15, 163–170. [Google Scholar] [CrossRef]
- Oh, S.S.; Narver, H.L. Mouse and Rat Anesthesia and Analgesia. Curr. Protoc. 2024, 4, e995. [Google Scholar] [CrossRef]
- Lindh, E.; Meller, A.; Raekallio, M. Effects of Vatinoxan in Rats Sedated with a Combination of Medetomidine, Midazolam and Fentanyl. Acta Vet. Scand. 2024, 66, 23. [Google Scholar] [CrossRef]
- Grubb, T. Inhalant Anesthetics: He’s Got Gas! In Questions and Answers in Small Animal Anesthesia; Wiley: Hoboken, NJ, USA, 2015; pp. 93–99. [Google Scholar] [CrossRef]
- Jedlicka, J.; Groene, P.; Linhart, J.; Raith, E.; Mustapha, D.; Conzen, P. Inhalational Anaesthetics: An Update on Mechanisms of Action and Toxicity. J. Exp. Neurol. 2021, 2, 62–69. [Google Scholar] [CrossRef]
- Adam, C.; Kayal, C.; Ercole, A.; Contera, S.; Ye, H.; Jerusalem, A. Action of the General Anaesthetic Isoflurane Reveals Coupling between Viscoelasticity and Electrophysiological Activity in Individual Neurons. Commun. Phys. 2023, 6, 174. [Google Scholar] [CrossRef]
- Flecknell, P.A. Laboratory Animal Anaesthesia and Analgesia (Fifth Edition): Chapter 6: Anaesthesia of Common Laboratory Species: Special Considerations; Academic Press: Cambridge, MA, USA, 2023; pp. 215–282. [Google Scholar] [CrossRef]
- De Heer, J.; Holst, J.J. Sulfonylurea Compounds Uncouple the Glucose Dependence of the Insulinotropic Effect of Glucagon-Like Peptide 1. Diabetes 2007, 56, 438–443. [Google Scholar] [CrossRef]
- Christiansen, C.B.; Svendsen, B.; Holst, J.J. The VGF-Derived Neuropeptide TLQP-21 Shows No Impact on Hormone Secretion in the Isolated Perfused Rat Pancreas. Horm. Metab. Res. 2015, 47, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Parada, E.; Eisentraut, A.M.; Unger, R.H. Pancreatic Glucagon Secretion in Normal and Diabetic Subjects. Am. J. Med. Sci. 1969, 257, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Sener, A.; Lebrun, P.; Blachier, F.; Malaisse, W.J. Stimulus-Secretion Coupling of Arginine-Induced Insulin Release: Insulinotro Action of Agmatine. Biochem. Pharmacol. 1989, 38, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Brand, C.L.; Jorgensen, P.N.; Knigge, U.; Warberg, J.; Svendsen, I.; Kristensen, J.S.; Holst, J.J. Role of Glucagon in Maintenance of Euglycemia in Fed and Fasted Rats. Am. J. Physiol. Metab. 1995, 269, E469–E477. [Google Scholar] [CrossRef]
- Ørskov, C.; Jeppesen, J.; Madsbad, S.; Holst, J.J. Proglucagon Products in Plasma of Noninsulin-Dependent Diabetics and Nondiabetic Controls in the Fasting State and after Oral Glucose and Intravenous Arginine. J. Clin. Investig. 1991, 87, 415–423. [Google Scholar] [CrossRef]
- Smith, W. Responses of Laboratory Animals to Some Injectable Anaesthetics. Lab. Anim. 1993, 27, 30–39. [Google Scholar] [CrossRef]
- Buhr, P.; Kolstrup, S.; Nikolajsen, L.L.; Bollen, P. The Anaesthetic Effects of Ketamine/Xylazine/Midazolam in C57Bl/6JRj Mice. Scand. J. Lab. Anim. Sci. 2023, 49, 21–27. [Google Scholar] [CrossRef]
- Wang, L.; Holland, L.; Fong, R.; Khokhar, S.; Fox, A.P.; Xie, Z. A Pilot Study Showing That Repeated Exposure to Stress Produces Alterations in Subsequent Responses to Anesthetics in Rats. PLoS ONE 2019, 14, e0214093. [Google Scholar] [CrossRef]
- Chemali, J.J.; Kenny, J.D.; Olutola, O.; Taylor, N.E.; Kimchi, E.Y.; Purdon, P.L.; Brown, E.N.; Solt, K. Ageing Delays Emergence from General Anaesthesia in Rats by Increasing Anaesthetic Sensitivity in the Brain. BJA Br. J. Anaesth. 2015, 115, i58. [Google Scholar] [CrossRef]
- Hendrick, G.K.; Gjinovci, A.; Baxter, L.A.; Mojsov, S.; Wollheim, C.B.; Habener, J.F.; Weir, G.C. Glucagon-like Peptide-I-(7–37) Suppresses Hyperglycemia in Rats. Metabolism 1993, 42, 1–6. [Google Scholar] [CrossRef]
- Galsgaard, K.D.; Abba, V.; Kuhre, R.E.; Holst, J.J.; Hartmann, B. In Vivo Inhibition of Dipeptidyl Peptidase 4 and Neprilysin Activity Enables Measurement of GLP-1 Secretion in Male Rats. J. Endocr. Soc. 2025, 9, bvaf173. [Google Scholar] [CrossRef]
- Zander, M.; Madsbad, S.; Madsen, J.L.; Holst, J.J. Effect of 6-Week Course of Glucagon-like Peptide 1 on Glycaemic Control, Insulin Sensitivity, and β-Cell Function in Type 2 Diabetes: A Parallel-Group Study. Lancet 2002, 359, 824–830. [Google Scholar] [CrossRef] [PubMed]
- Svane, M.S.; Bojsen-Møller, K.N.; Martinussen, C.; Dirksen, C.; Madsen, J.L.; Reitelseder, S.; Holm, L.; Rehfeld, J.F.; Kristiansen, V.B.; van Hall, G.; et al. Postprandial Nutrient Handling and Gastrointestinal Hormone Secretion After Roux-En-Y Gastric Bypass vs Sleeve Gastrectomy. Gastroenterology 2019, 156, 1627–1641.e1. [Google Scholar] [CrossRef] [PubMed]
- Quddusi, S.; Vahl, T.P.; Hanson, K.; Prigeon, R.L.; D’Alessio, D.A. Differential Effects of Acute and Extended Infusions of Glucagon-Like Peptide-1 on First- and Second-Phase Insulin Secretion in Diabetic and Nondiabetic Humans. Diabetes Care 2003, 26, 791–798. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Angel, I.; Bidet, S.; Langer, S.Z. Pharmacological Characterization of the Hyperglycemia Induced by Alpha-2 Adrenoceptor Agonists. J. Pharmacol. Exp. Ther. 1988, 246, 1098–1103. [Google Scholar] [CrossRef]
- Kjems, L.L.; Holst, J.J.; Vølund, A.; Madsbad, S. The Influence of GLP-1 on Glucose-Stimulated Insulin Secretion Effects on β-Cell Sensitivity in Type 2 and Nondiabetic Subjects. Diabetes 2003, 52, 380–386. [Google Scholar] [CrossRef]
- Vaughan, K.L.; Szarowicz, M.D.; Herbert, R.L.; Mattison, J.A. Comparison of Anesthesia Protocols for Intravenous Glucose Tolerance Testing in Rhesus Monkeys. J. Med. Primatol. 2014, 43, 162–168. [Google Scholar] [CrossRef]
- Fagerholm, V.; Haaparanta, M.; Scheinin, M. α 2-Adrenoceptor Regulation of Blood Glucose Homeostasis. Basic Clin. Pharmacol. Toxicol. 2011, 108, 365–370. [Google Scholar] [CrossRef]
- Samols, E.; Weir, G.C. Adrenergic Modulation of Pancreatic A, B, and D Cells Alpha-Adrenergic Suppression and Beta-Adrenergic Stimulation of Somatostatin Secretion, Alpha-Adrenergic Stimulation of Glucagon Secretion in the Perfused Dog Pancreas. J. Clin. Investig. 1979, 63, 230–238. [Google Scholar] [CrossRef]
- Saito, M.; Saitoh, T.; Inoue, S. Alpha 2-Adrenergic Modulation of Pancreatic Glucagon Secretion in Rats. Physiol. Behav. 1992, 51, 1165–1171. [Google Scholar] [CrossRef]
- Kawano, T.; Tanaka, K.; Chi, H.; Eguchi, S.; Yamazaki, F.; Kitamura, S.; Kumagai, N.; Yokoyama, M. Biophysical and Pharmacological Properties of Glucagon-like Peptide-1 in Rats under Isoflurane Anesthesia. Anesth. Analg. 2012, 115, 62–69. [Google Scholar] [CrossRef]
- Rorsman, P.; Ramracheya, R.; Rorsman, N.J.G.; Zhang, Q. ATP-Regulated Potassium Channels and Voltage-Gated Calcium Channels in Pancreatic Alpha and Beta Cells: Similar Functions but Reciprocal Effects on Secretion. Diabetologia 2014, 57, 1749–1761. [Google Scholar] [CrossRef]
- MacDonald, P.E.; Marinis, Y.Z.D.; Ramracheya, R.; Salehi, A.; Ma, X.; Johnson, P.R.V.; Cox, R.; Eliasson, L.; Rorsman, P. A KATP Channel-Dependent Pathway within α Cells Regulates Glucagon Release from Both Rodent and Human Islets of Langerhans. PLoS Biol. 2007, 5, e143. [Google Scholar] [CrossRef]
- Wewer Albrechtsen, N.J.; Kuhre, R.E.; Windeløv, J.A.; Ørgaard, A.; Deacon, C.F.; Kissow, H.; Hartmann, B.; Holst, J.J. Dynamics of Glucagon Secretion in Mice and Rats Revealed Using a Validated Sandwich ELISA for Small Sample Volumes. Am. J. Physiol. Metab. 2016, 311, E302–E309. [Google Scholar] [CrossRef]
- Yu, Q.; Li, J.; Dai, C.L.; Li, H.; Iqbal, K.; Liu, F.; Gong, C.X. Anesthesia with Sevoflurane or Isoflurane Induces Severe Hypoglycemia in Neonatal Mice. PLoS ONE 2020, 15, e0231090. [Google Scholar] [CrossRef]
- Nummela, A.J.; Laaksonen, L.T.; Laitio, T.T.; Kallionpää, R.E.; Långsjö, J.W.; Scheinin, J.M.; Vahlberg, T.J.; Koskela, H.T.; Aittomäki, V.; Valli, K.J.; et al. Effects of Dexmedetomidine, Propofol, Sevoflurane and S-Ketamine on the Human Metabolome: A Randomised Trial Using Nuclear Magnetic Resonance Spectroscopy. Eur. J. Anaesthesiol. 2022, 39, 521–532. [Google Scholar] [CrossRef]
- Tharp, W.G.; Breidenstein, M.W.; Friend, A.F.; Bender, S.P.; Raftery, D. The Neuroendocrine Stress Response Compensates for Suppression of Insulin Secretion by Volatile Anesthetic Agents: An Observational Study. Physiol. Rep. 2023, 11, e15603. [Google Scholar] [CrossRef]
- Russell, W.; Burch, R. The Principles of Humane Experimental Technique; Methuen & Co. Limited: London, UK, 1959. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Anesthetic Regimen | Surgical Depth Achieved? | Stability of Preparation | Hormone Assays? |
|---|---|---|---|
| Protocol 1: Fentanyl/fluanisone/midazolam | Yes | Stable | Yes |
| Protocol 2: Ketamine/xylazine | No | Inadequate depth | No |
| Protocol 3: Ketamine/xylazine/midazolam | No | Inadequate depth | No |
| Protocol 4: Fentanyl/medetomidine/midazolam | No | Inadequate depth | No |
| Protocol 5: Ketamine/dexmedetomidine | Variable | Occasionally some reflexes | Yes |
| Protocol 6: Isoflurane/buprenorphine | Yes | Stable | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Abba, V.; Nielsen, A.B.E.; Buhr, P.; Hammelev, K.P.; Holst, J.J.; Lobato, C.B. Effects of Preceding Anesthesia Protocols on Insulin and Glucagon Secretion from Isolated Perfused Rat Pancreas Preparations. Anesth. Res. 2026, 3, 6. https://doi.org/10.3390/anesthres3010006
Abba V, Nielsen ABE, Buhr P, Hammelev KP, Holst JJ, Lobato CB. Effects of Preceding Anesthesia Protocols on Insulin and Glucagon Secretion from Isolated Perfused Rat Pancreas Preparations. Anesthesia Research. 2026; 3(1):6. https://doi.org/10.3390/anesthres3010006
Chicago/Turabian StyleAbba, Valentina, Amalie B. E. Nielsen, Petra Buhr, Karsten Pharao Hammelev, Jens J. Holst, and Carolina B. Lobato. 2026. "Effects of Preceding Anesthesia Protocols on Insulin and Glucagon Secretion from Isolated Perfused Rat Pancreas Preparations" Anesthesia Research 3, no. 1: 6. https://doi.org/10.3390/anesthres3010006
APA StyleAbba, V., Nielsen, A. B. E., Buhr, P., Hammelev, K. P., Holst, J. J., & Lobato, C. B. (2026). Effects of Preceding Anesthesia Protocols on Insulin and Glucagon Secretion from Isolated Perfused Rat Pancreas Preparations. Anesthesia Research, 3(1), 6. https://doi.org/10.3390/anesthres3010006