Abstract
Silvopastoral systems, integrating trees, forages, and livestock, are recognized as a sustainable approach to livestock production. This study evaluated the effect of shade vs. open-field conditions on the yield and nutritive value of Brachiaria decumbens in three systems—Inga edulis (guaba), Eucalyptus torrelliana (eucalyptus), and Cedrelinga cateniformis (tornillo)—in the Peruvian Amazon. A 3 × 2 factorial design with three replicates was used to analyze forage production (mass and mass by harvest), quality (NDF, protein, digestibility, metabolizable energy), and soil variables (bulk density, pH, organic matter, moisture, porosity, etc.). Most interactions were non-significant, except forage mass by harvest (FMH, p = 0.0328). B. decumbens in the guaba system under shade had the highest FMH (1406 kg DM ha−1), while the tornillo system showed elevated protein (10.63%). Protein was significantly higher under shade (9.55%) than in open field. Eucalyptus increased neutral detergent fiber (69.72%), whereas guaba provided the greatest metabolizable energy (8.08 MJ kg−1 DM). Soil analyses revealed improved moisture and CEC under guaba, while tornillo boosted soil phosphorus. Principal component analysis confirmed these associations (82.3% variance). B. decumbens grown under the shade of guaba or tornillo appears to enhance forage productivity and soil fertility, representing a promising strategy for sustainable tropical livestock management.
1. Introduction
Silvopastoral systems (SPS), which combine trees, forage grasses, and livestock within the same production area, are increasingly recognized for improving sustainability in tropical livestock systems [1,2]. These systems can boost animal welfare, conserve biodiversity, and sequester carbon [3,4,5]. The introduction of leguminous and non-leguminous trees frequently enhances nutrient cycling and microclimate regulation, reducing temperatures and increasing soil moisture [6,7].
In South America, silvopastoral systems have expanded considerably; for example, in Brazil it is estimated that more than 11.5 million hectares are managed under such systems [8], with extensive implementations also found in countries like Argentina and Colombia. In Peru, approximately 1.5 million hectares are managed using integrated silvopastoral practices [9,10,11]. Commonly used tree species in South America include Leucaena leucocephala, Gliricidia sepium, and Erythrina spp., which not only provide shade but also fix nitrogen—often in the range of 80–350 kg N ha−1 year−1—and serve as supplemental forage [11,12,13,14]. In the Peruvian Amazon, native species such as Inga edulis (guaba) and Cedrelinga cateniformis (tornillo) are being integrated into silvopastoral systems to enhance soil properties and improve animal performance [7,15] but also to provide timber, shade, and, in some cases, fruits or fuel [15,16]. In terms of N fixation, guaba may fix approximately 44 kg N ha−1 year−1 [17] or release 145 kg N ha−1 year−1 through leaf litter decomposition [18], while tornillo, as a leguminous species’ amount of N fixation is limited, however thereby enhancing soil fertility.
The variation in canopy architecture among these species—reflecting differences in crown density and form—plays a crucial role in determining the amount and distribution of shade. Excessive shade (>40%) may reduce forage biomass [19,20], whereas moderate shade (30–40%) can improve crude protein and digestibility (10–30%) whereas variations in NDF will depend on the specific interactions between the tree species and the environmental conditions [21,22]. Although specific quantitative data regarding the percentage of shade for species such as guaba, eucalyptus, or tornillo are limited, their distinct crown morphologies suggest that they provide varying amounts of shade. For instance, guaba generally exhibits a broad, dense crown that can generate significant or moderate shade, followed by tornillo with moderate shade, while eucalyptus often has a more open crown.
Species like Eucalyptus torrelliana can exert allelopathic effects that influence moisture and nutrient availability [23,24]. Dispersed trees in silvopastoral systems, however, offer several soil benefits. They promote an increase in soil carbon stocks—approximately 9.55% in the top 15 cm and 10.3% in the 15–30 cm layer [5]—and improve soil porosity while stimulating beneficial microbial activity [25,26]. This microbial community, which includes nitrogen-fixing bacteria (e.g., Rhizobium, Azospirillum, Bacillus, and Pseudomonas) and arbuscular mycorrhizal fungi (e.g., Glomus and Acaulospora), further enhances nutrient cycling and soil fertility [6]. These enhancements lead to better water retention and overall soil quality, ultimately supporting improved forage yield and nutritive value [16].
Brachiaria decumbens is a key forage grass in the Peruvian tropics and plays a central role in local silvopastoral systems. Along with Brachiaria brizantha, it represents one of the most predominant forage species used by livestock producers in the region [11,27]. Known for its high productivity, adaptability to low-fertility soils, and favorable nutritional profile, B. decumbens is well-suited for integration with tree species in these systems [28,29]. This integration not only optimizes forage yield but also improves microclimatic conditions and soil fertility, thereby supporting sustainable livestock production in the Peruvian Amazon [11,27]. The widespread adoption of these grasses highlights their importance in maintaining resilient and productive agroecosystems under tropical conditions.
Given the need for sustainable livestock in the Peruvian Amazon, this study aims to assess the effect of light conditions (shade vs. open field) on B. decumbens yield and quality in three silvopastoral systems with different tree species. We hypothesize that shade (regardless of the canopy cover percentage) from leguminous and non-leguminous trees will affect forage productivity and nutritive value, while also influencing soil characteristics. Specifically, we (1) evaluate forage mass and nutritional traits (crude protein, neutral detergent fiber, metabolizable energy, in vitro digestibility) under two light conditions; (2) assess soil physical and chemical parameters in each system; (3) determine which tree species offers better synergy with B. decumbens for sustainable forage production. Although this study focuses on agronomic and environmental parameters, it is recognized that silvopastoral systems can also enhance profitability and social cohesion in rural areas [30]; these aspects will be addressed in future studies.
2. Materials and Methods
2.1. Site Description
The study was conducted on three private farms in the Soritor district (Moyobamba province, San Martín region, Peru), each ranging from approximately 5 to 25 hectares in total area. Local soils are predominantly clay loam, characterized by moderate fertility and great water retention capacity. The district lies between 860 and 3000 m.a.s.l., gently sloping to hilly topography. The mean annual rainfall in 2017 was 178 mm, and the mean temperature was 28 °C, based on monthly records (Figure 1). Precipitation and temperature data from 2016 are also compared, illustrating the interannual variation in climate. These farms employ rotational stocking, typically allowing a 45-day rest period before regrazing the pastures.
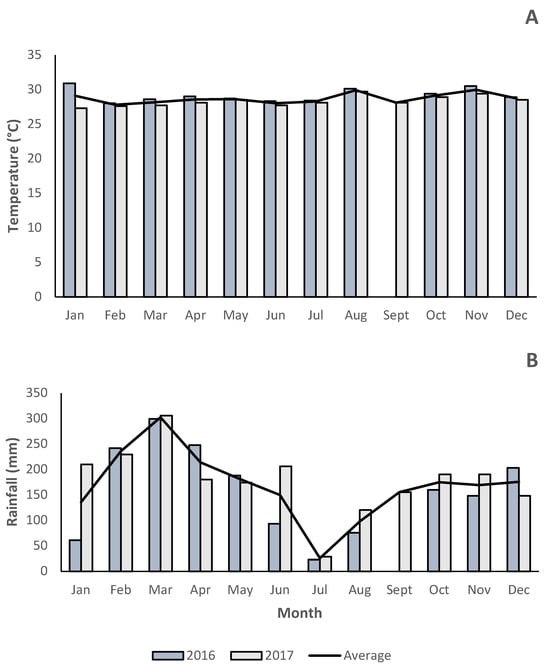
Figure 1.
Average precipitation, maximum and minimum long-term temperatures (2016–2017). (A): temperature, (B): rainfall [31].
2.2. Treatments and Sampling
Three silvopastoral systems (SPS) were compared: (1) dispersed Inga edulis (guaba) trees (5 years old, ~7.7 m tall, 5–7 trees ha−1); (2) living fences of Eucalyptus torrelliana (5 years old, 12 m tall, five trees per 10 linear meters); and (3) dispersed Cedrelinga cateniformis (tornillo) trees (~35–40 years old, ~23 m tall, ~6–8 trees ha−1). Each system was evaluated under two light conditions—open-field (full sunlight with no canopy shade) and shaded (with canopy)—yielding six treatments arranged in a 3 × 2 factorial design with three replicates per farm. The shading factor was defined solely by the observed effect of shade relative to open-field conditions, without considering the specific percentage of shade provided by each species.
Brachiaria decumbens was selected based on previous studies identifying it as one of the predominant grasses in the region [11,27] while planting densities and pasture management followed local producers’ practices. Previous studies at the location [32,33] of this study describe the management practices as follows: an average of 83 head per farm, a rotational grazing cycle of 45 days, NPK fertilization, and the genotype consisted of a crossbreed of Gyr × Brown Swiss and others.
Two separate evaluations were conducted. For initial forage mass (FM) and forage quality analysis, samples were collected after a 45-day rest period (prior to grazing) under a rotational stocking regimen, with forage clipped at approximately 5 cm above ground level. Seven replicate samples were taken per treatment using a 0.50 × 0.50 m wooden frame arranged in a zigzag pattern throughout each paddock. The entire biomass within the frame was weighed, and a 200 g subsample was dried at 55 °C to determine dry matter, which was scaled up to yield (kg DM ha−1). A composite sample from replicates of each SPS was also prepared for laboratory quality analysis.
For forage mass by harvest (FMH), representing forage mass at harvest frequencies of 30, 45, 60, and 75 days, samples were obtained from 3 × 6 m2 plots under both light conditions. Exclusion cages of identical dimensions were installed to prevent grazing before sampling, and forage was clipped at approximately 5 cm above ground level. Four replicate samples were collected per treatment, with a 200 g subsample from each used to calculate yield. Nutritional quality parameters—including dry matter (DM), crude protein (CP) and neutral detergent fiber (NDF) were analyzed according to the methodology described by AOAC [34], and in vitro dry matter digestibility (IVDMD)—were determined using the technique described by Tilley and Terry [35] following ANKOM [36] procedures. Metabolizable energy (ME) was estimated using the equation proposed by Indah et al. [37].
where ME is metabolizable energy, NDF is neutral detergent fiber.
ME (MJ kg−1 DM) = 46.93 — 0.52 × NDF
Soil sampling and analysis were conducted at a depth of 0–15 cm in each SPS, measuring soil moisture, bulk density [38], porosity [39], pH, organic matter (OM) [40], phosphorus (P), cation exchange capacity (CEC) [41], and macronutrients (Ca, Mg, Na, K, Al). The procedures followed standard protocols, and the analyses were carried out at the Soil Analysis Laboratory of the National Agrarian University La Molina. These analyses were performed in a previous study, and the results can be verified by Diaz et al. [33].
2.3. Statistical Analysis
This study conducted two distinct evaluations—each with its own statistical approach. The first assessment simultaneously measured FM and forage quality using a completely randomized design arranged in a 3 × 2 factorial scheme with three replications. The second evaluation focused on FM at different harvest intervals (FMH) using a repeated measures design, with harvest frequency as the repeated factor (30, 45, 60, and 75 days).
For both evaluations, SPS and light conditions were treated as fixed effects, while replicate was considered a random effect. Analyses were performed using PROC MIXED in SAS (version 9.4; SAS Institute), with LSMEANS compared via the PDIFF option and pairwise comparisons adjusted by Tukey’s test (p < 0.05). Soil property data (e.g., CEC, pH, OM, P) were exclusively utilized for principal component analysis (PCA), which was executed in R using the prcomp function (stats package) to explore relationships among forage quality parameters (CP, NDF, IVDMD) and soil characteristics. Residual normality and variance homogeneity were verified using the Shapiro–Wilk and Levene’s tests, respectively, with log transformations applied when necessary to meet model assumptions. Model selection was based on the Akaike Information Criterion (AIC), ensuring the simplest model that adequately explained data variability.
3. Results
3.1. Forage Mass
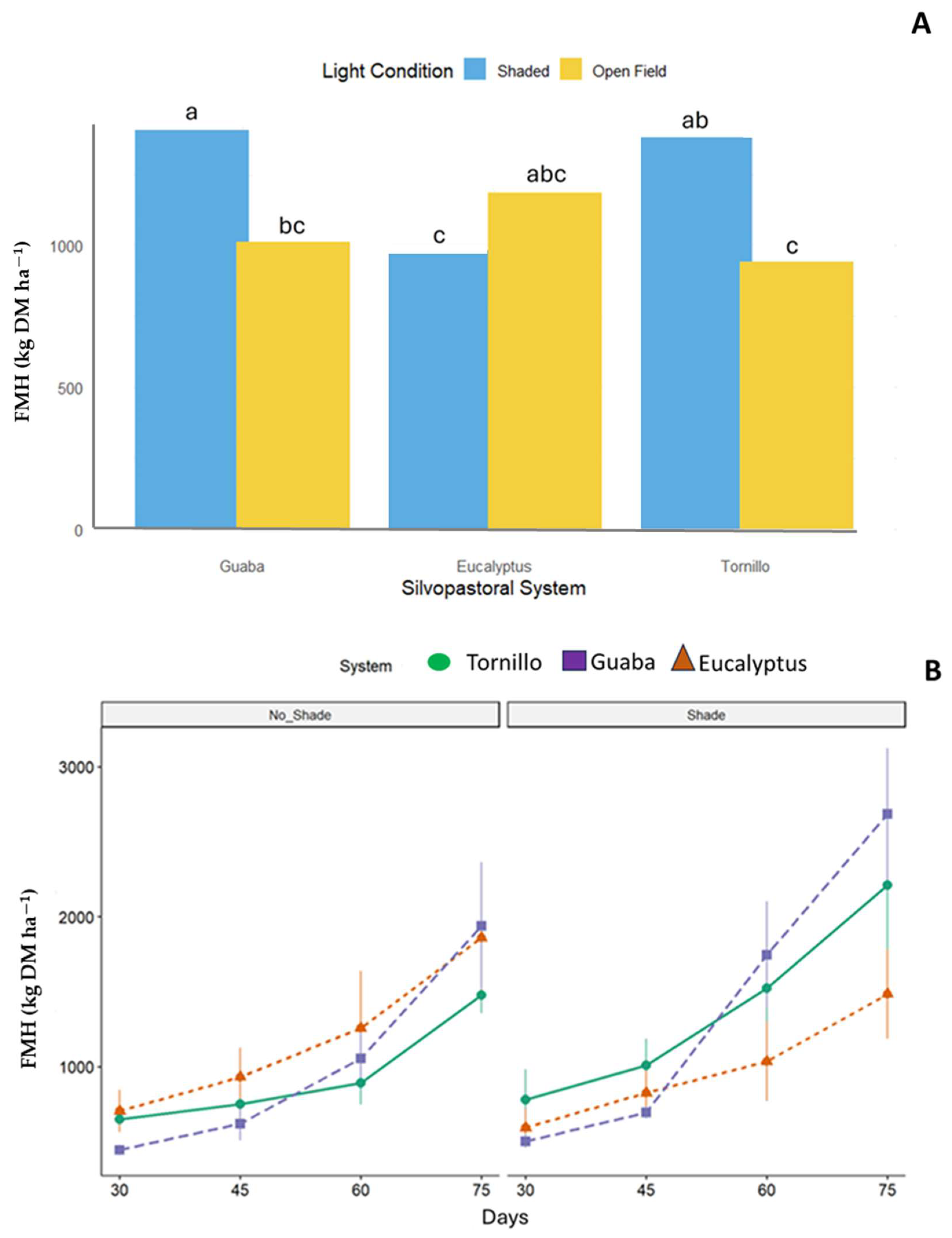
No significant interactions (p > 0.05) were found between SPS and light conditions for forage mass (FM), protein (P), NDF, IVDMD, ME, or productive efficiency, except for forage availability by harvest (FAH) (p = 0.0328) (Table 1, Figure 2). A significant SPS × light condition interaction was detected for FMH. B. decumbens in guaba systems under shade produced the highest yield (1406 kg DM ha−1), exceeding both eucalyptus under shade (983 kg DM ha−1) and tornillo in open-field conditions (942 kg DM ha−1, Figure 2). The greatest FMH values were recorded at the 75-day cutting interval.

Table 1.
Forage mass and quality in SPS under different light conditions.
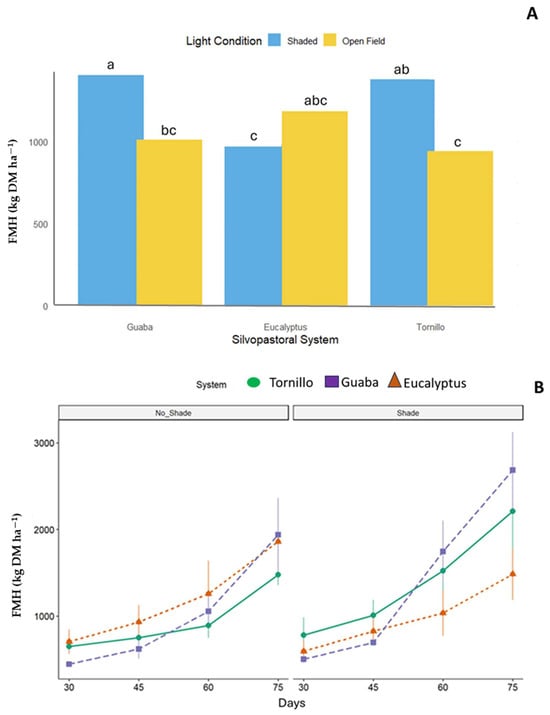
Figure 2.
Forage mass by harvest (FMH) of Brachiaria decumbens (kg DM ha−1) × three silvopastoral systems (Guaba, Eucalyptus, Tornillo) and two light conditions (Shaded, Open Field), p = 0.0328. Bars followed by different lowercase letters are significantly different at the 5% probability level. (A) = SPS × Interaction, (B) = Forage mass by harvest.
3.2. Nutritive Value (Protein, NDF, IVDMD, ME)
Protein content was higher (p = 0.0045) in the system with tornillo compared to the systems with eucalyptus and guaba. This represented an approximate 48.2% increase in protein content compared to eucalyptus and 24% compared to guaba. Additionally, shaded conditions have significantly greater (p = 0.0431) protein content (18.9%) compared to direct light conditions (p < 0.05). As NDF, it was highest in the system with eucalyptus (69.72%) and lowest in the system with guaba (p = 0.0215), while light conditions had no significant effect. ME was higher (p = 0.0214) in the systems with guaba (8.08 MJ kg−1 DM) and tornillo (7.77 MJ kg−1 DM) and lower in eucalyptus systems. Regarding IVDMD ranged from 37.6% (eucalyptus) to 41.9% (guaba), with no significant light effect (p > 0.05).
3.3. Soil and Forage Correlations (PCA)
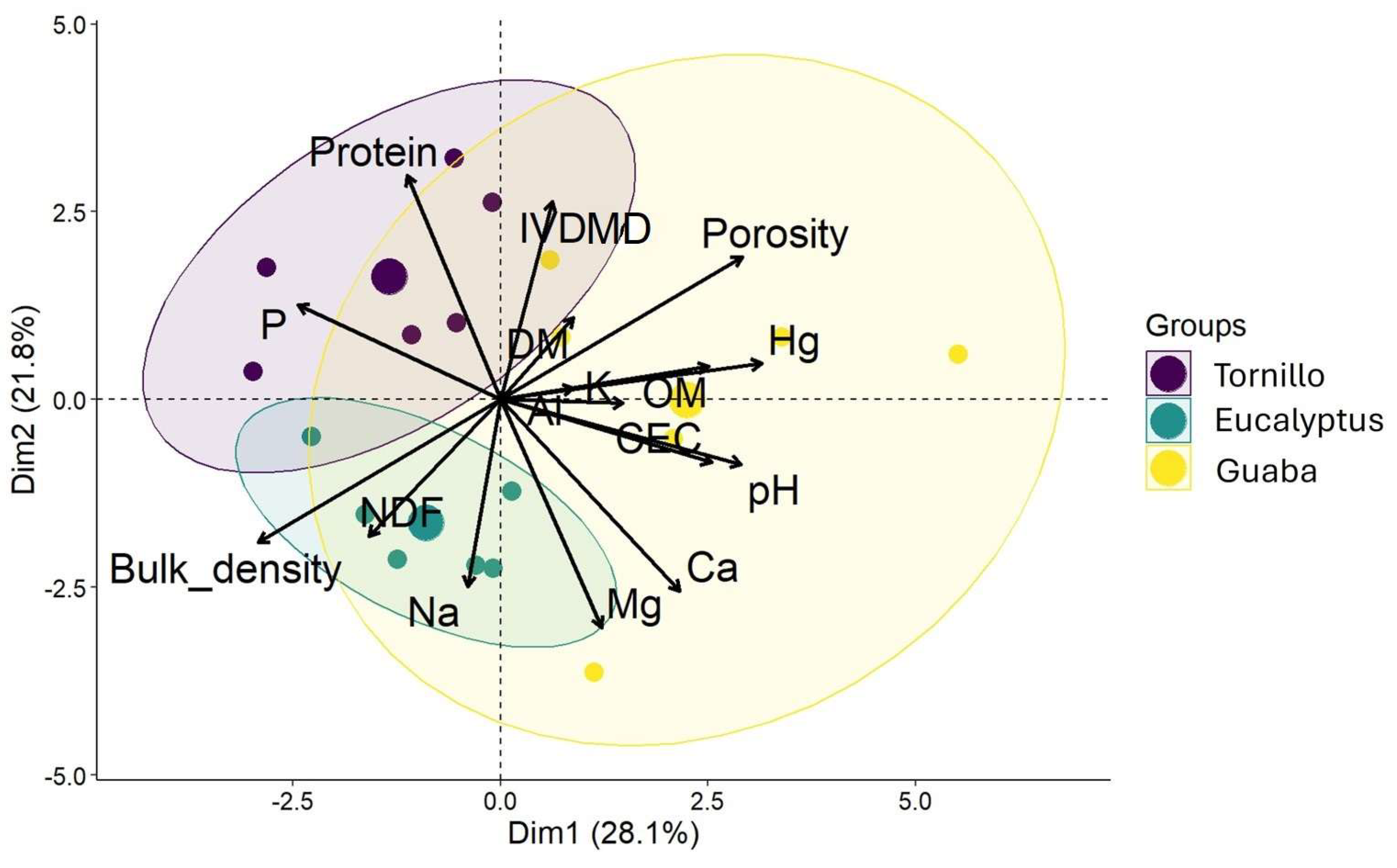
Figure 3 displays the PCA associating forage quality (protein, NDF, IVDMD, DM, ME) and soil parameters (moisture, OM, pH, CEC, P, Ca, Mg, Na, K, Al) with the three SPS (guaba, eucalyptus, tornillo). PC1 and PC2 accounted for 28.1% and 21.8% of the total variance, respectively (49.9% combined). Guaba correlated strongly with soil moisture, CEC, OM, and certain forage variables (e.g., IVDMD). Eucalyptus correlated with higher NDF and potential Na accumulation. Tornillo showed a positive association with forage protein and soil P. Although 50% of the variability is explained in these first two dimensions, further components carry additional information on the system’s complexity.
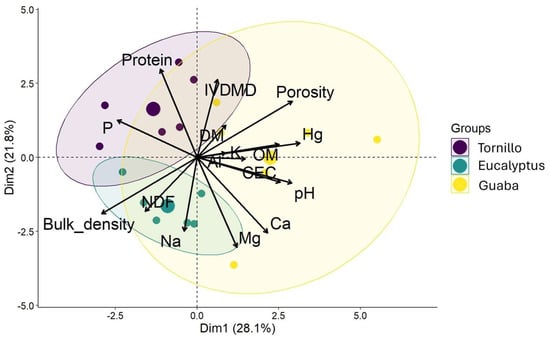
Figure 3.
Principal component analysis (PCA) of the forage quality of Brachiaria decunbens and soil characteristics in different SPS and under different light conditions. Forage parameters; Protein: protein, IVDMD: in vitro dry matter digestibility, DM: dry matter, NDF: neutral detergent fiber; Soil parameters, Hg: soil moisture, OM: organic matter, Mg: magnesium, Ca: calcium, CEC: cation exchange capacity, Na: sodium, P: phosphorus, K: potassium.
Table 2 provides a clearer numerical breakdown of the principal component analysis (PCA), allowing for a more precise interpretation of how each variable contributes to the principal components. The PCA of variables in different silvopastoral systems showed that the first five components explain 82.3% of the total variance, highlighting key factors that differentiate these systems. PC1 (28.1%) is primarily associated with soil properties, including porosity, pH, organic matter, and cation exchange capacity, while negatively correlated with bulk density and phosphorus. PC2 (21%) relates to forage quality, with high contributions from crude protein and in vitro dry matter digestibility (IVDMD) and a negative association with magnesium and calcium. PC3 (15.6%) captures variations in soil fertility, being positively linked to potassium, CEC, and aluminum, but negatively to organic matter and NDF. PC4 (10.7%) highlights soil acidity, with strong negative associations with aluminum and CEC, while PC5 (6.9%) reflects soil structure and forage composition, with dry matter and bulk density contributing positively and sodium and potassium negatively. Compared to the biplot, which visually illustrates group clustering and variable relationships, Table 2 offers a more detailed and quantifiable representation of the PCA results, making it easier to interpret the influence of each variable in distinguishing silvopastoral systems.
Table 2.
Eigenvalues of principal components, obtained from the analysis of the variables correlated in different silvopastoral systems.
4. Discussion
4.1. Forage Yield Under Shade Conditions
While overall forage mass (FM) did not differ significantly among the silvopastoral systems, B. decumbens under the guaba system showed a trend toward higher FM compared to those in the eucalyptus and tornillo systems. According to Díaz et al. [33], preliminary analyses on the same farms revealed that the guaba system exhibited notably improved soil properties. Similar positive effects on soil properties from the implementation of guaba were also reported by Freitas et al. [42], Alegre et al. [43], Nichols et al. [44], and da Silva et al. [45]. In this system, soils demonstrated a substantially higher cation exchange capacity (around 25.5 cmolc/kg), which enhances nutrient retention, even though they present lower available phosphorus (approximately 0.93 mg/kg) and elevated aluminum levels. These attributes, together with a moderate organic matter content (about 2.9%), are likely linked to a larger crown volume and increased litter production that promote better moisture retention and more efficient nutrient recycling, thereby accelerating regrowth.
In contrast, the tornillo system showed soils with higher available phosphorus (approximately 3.4 mg/kg) but lower cation exchange capacity (around 9.1 cmolc/kg) and reduced levels of essential cations (Ca2+ and Mg2+), suggesting that its nutrient retention capacity is more limited. Meanwhile, the eucalyptus system exhibited the highest organic matter content (nearly 3.8%) along with elevated calcium and magnesium levels and virtually no aluminum, creating a fertile soil environment despite its cation exchange capacity being similar to that of the tornillo system.
Furthermore, a significant interaction between light conditions and silvopastoral systems was observed in forage mass by harvest (FMH), with shaded conditions yielding higher FMH in both the Guaba and tornillo systems than in the eucalyptus systems. This likely reflects the ability of moderate shade to mitigate evapotranspiration and thermal stress, thereby creating more favorable microclimatic conditions for regrowth [10,21,22]. Additionally, analyses of FM across different cutting frequencies under shade indicate that regrowth dynamics are particularly pronounced in the Guaba and tornillo systems, suggesting that optimal harvest management under moderate shade can enhance both forage quality and yield. These findings, consistent with previous agroforestry studies, highlight that while overall biomass differences may not be statistically significant, the regrowth responses under shaded conditions reveal important agronomic benefits linked to the specific soil fertility attributes of each system as demonstrated by Díaz et al. [33].
4.2. Forage Quality (Protein, NDF, Digestibility, ME)
Tornillo—although a legume with limited data on its nitrogen-fixing capacity—exhibited notably higher protein content than the other systems. This suggests that even modest nitrogen fixation by tornillo may enhance soil fertility and, in turn, improve forage nutritional quality. Protein content was especially elevated under shade in tornillo-based SPS, possibly due to the older tree age and the establishment of stable symbiotic relationships with nitrogen-fixing bacteria [46]. Several studies have demonstrated that beneficial rhizobacteria, including members of the genera Rhizobium, Bacillus, and Pseudomonas, can progressively increase soil nitrogen accumulation, which has been linked to enhanced protein content in pastures [47]. This enhanced protein level is crucial for ruminants, as it can improve microbial protein synthesis in the rumen and thereby positively influence animal performance. Eucalyptus-based SPS displayed higher neutral detergent fiber (NDF) content, which could be related to allelopathic effects and resource competition [48]. The phenolic compounds released by Eucalyptus can disrupt enzymes that remodel cell walls, leading to thicker, more lignified tissues. As a result, the grass directs more energy towards building structural carbohydrates rather than growth, which raises its NDF content. Additionally, this shift may reduce nutrient uptake efficiency, further contributing to the higher fiber levels observed [49]. Furthermore, partial shading often induces morphological changes in grasses, such as increased stem elongation and a reduced leaf/stem ratio, which can elevate NDF content [50]. Since NDF levels above 65–70% are generally considered high for ruminants, exceeding this range could limit intake and reduce overall digestibility [49,51]. In summary, we postulate that the combined effects of reduced thermal stress and enhanced organic inputs from trees positively modulate the nutritional quality of the forage, as evidenced by improvements in parameters such as protein content and digestibility. Although specific measurements of leaf/stem ratio and canopy height were not obtained in this study, incorporating these parameters in future research would help clarify the exact mechanisms behind the higher NDF observed under certain shade conditions.
The in vitro dry matter digestibility (IVDMD) was not affected by SPS and light conditions; however, in general terms improving digestibility could potentially increase metabolizable energy (ME). Higher IVDMD means that ruminants can more efficiently extract nutrients, directly influencing weight gain or milk production. However, if shading is too intense, the reduction in total biomass could offset these nutritional gains, as forage availability remains a critical factor in grazing systems [24,25]. Variations in light availability can alter chlorophyll synthesis and enzymatic activity, thereby reducing the formation of structural compounds and enhancing nutrient availability for protein synthesis [52]. Our findings suggest that species that provide balanced levels of shading, such as tornillo or guaba, can maintain or even improve forage quality characteristics (protein, EM) while still producing acceptable biomass levels. This happens probably because a reduction in light intensity decreases oxidative stress, thereby enhancing photosynthetic efficiency and promoting the accumulation of secondary metabolites that improve forage quality [53].
4.3. Soil–Forage Interactions from the PCA
Principal component analysis (PCA) showed that each tree species influences specific soil parameters, affecting forage quality. Guaba is associated with improved soil structure, moisture, and nutrient retention, contributing to better digestibility and overall forage quality. Previous studies highlight that leaf litter decomposition from Inga edulis can boost organic matter content, raising CEC and enhancing water retention [42,54]. For instance, literature in similar silvopastoral systems suggests that forage biomass production may increase by approximately 10–20% under optimal conditions, organic matter decomposition may result in a 15–20% turnover over a 60-day period, water retention in soil could improve by around 15–25%, and cation exchange capacity (CEC) might increase by roughly 10–20% compared to conventional systems. Tornillo, owing to its leguminous nature and deep-rooting system, enhances soil phosphorus levels and forage protein content. Its extensive roots not only access nutrients from deeper soil layers but also exude organic acids that solubilize phosphorus bound to soil minerals [55], making it more available to adjacent forages. Additionally, many fabaceae species stimulate beneficial soil microbial communities, which further promote nutrient cycling and the release of phosphorus. Although specific quantitative data for tornillo are limited, studies have shown that leguminous trees can improve soil fertility through these mechanisms [56,57]. This enhancement in phosphorus availability is particularly significant for B. decumbens, as phosphorus deficiency is a common constraint in tropical forage production.
Eucalyptus, however, appears to increase soil bulk density and the fiber content in the grass, a phenomenon that may be linked to its configuration as a live fence with an approximate density of five trees per linear meter. This concentrated arrangement could exacerbate root competition and limit soil porosity, thereby elevating fiber content in the adjacent forage. These factors suggest that careful management of Eucalyptus in silvopastoral systems is essential to mitigate potential negative impacts on forage productivity and quality. In addition, because eucalyptus species can have strong taproots and extensive lateral root systems, soil compaction and shifts in microbial communities may occur, leading to lower nutrient availability for grasses [25]. In addition, any allelopathic compounds released by eucalyptus leaf litter or root exudates might suppress beneficial soil microbes, thereby diminishing nutrient mineralization rates [24]. Consequently, producers who choose eucalyptus in their silvopastoral designs may require periodic liming, strategic fertilization, or careful canopy pruning to mitigate these effects.
The PCA findings confirm the interconnectedness between tree-soil interactions and forage nutritive traits, demonstrating that tree selection should be carefully aligned with site conditions and production goals. Future studies could extend this research by examining how tree age, stand density, and thinning/pruning regimes affect long-term soil properties and forage composition. Additionally, assessing microbial community shifts under different tree species would provide deeper insights into the mechanisms driving soil fertility changes in silvopastoral systems.
In summary, the PCA results reinforce the discussion by providing evidence of how tree species influence key soil and forage quality parameters. By understanding these interactions, silvopastoral system producers can make informed decisions to optimize both productivity and ecological benefits.
5. Conclusions
Integrating Brachiaria decumbens with different tree species in silvopastoral systems significantly influences forage yield, nutritional quality, and soil properties. In our study, the Inga edulis (guaba) system—characterized by relatively high canopy cover—resulted in the highest forage mass under shaded conditions. In contrast, the Eucalyptus system, with its taller yet more open canopy, was associated with increased neutral detergent fiber, which may limit forage digestibility. Meanwhile, the Cedrelinga cateniformis (tornillo) system, with its intermediate canopy cover, was linked to improved forage protein content and enhanced soil phosphorus levels. These findings suggest that leguminous species such as guaba and tornillo can promote soil nutrient cycling and improve forage quality, possibly through mechanisms related to deep rooting and effective nutrient uptake. Although some microclimatic parameters were not directly measured, the observed differences in forage performance provide valuable insights into the benefits of integrating trees into tropical livestock systems. Future research should focus on long-term evaluations of soil and microclimatic dynamics to further refine management practices for sustainable silvopastoral systems.
Author Contributions
Conceptualization: J.A., C.G. (Carlos Gómez), and M.D.; methodology: J.A., C.G. (Carlos Gómez), and M.D.; software: C.A.-H.; validation: C.A.-H. and M.D.; formal analysis: C.A.-H.; investigation: M.D.; resources: C.G. (Carlos Gómez); data curation: C.A.-H. and J.A.; writing—original draft preparation: C.G. (Carlos Garcia) and M.D.; writing—review and editing: C.G. (Carlos Garcia), J.A., C.G. (Carlos Gómez), C.A.-H., and M.D.; visualization: M.D. and C.G. (Carlos Garcia); supervision: C.G. (Carlos Gómez) and J.A.; project administration: C.G.(Carlos Gómez) and J.A.; funding acquisition: C.G. (Carlos Gómez). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by UNALM-PNIA Project “Innovación en la evaluación de sistemas silvopastoriles de selva alta peruana como estrategia de adaptación y mitigación al cambio climático”, grant number 010-2015, started from January 2016 to August 2019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data can be requested by email: maryondiaz@hotmail.com.
Acknowledgments
The authors would like to thank the UNALM-PNIA for their financial support, which enabled the execution of this research. They also express gratitude to the farmers in San Martin Region for providing the field to collect data for this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Huertas, S.M.; Bobadilla, P.E.; Alcántara, I.; Akkermans, E.; van Eerdenburg, F.J.C.M. Benefits of silvopastoral systems for keeping beef cattle. Animals 2021, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Gomes da Silva, I.A.; Dubeux, J.C.B., Jr.; De Melo, A.C.L.; Da Cunha, M.V.; Dos Santos, M.V.F.; Apolinário, V.X.O.; de Freitas, E. Tree legume enhances livestock performance in a silvopasture system. Agron. J. 2021, 113, 358–369. [Google Scholar] [CrossRef]
- Lemes, A.P.; Garcia, A.R.; Pezzopane, J.R.M.; Brandão, F.Z.; Watanabe, Y.F.; Cooke, R.F.; Sponchiado, M.; de Paz, C.C.P.; Camplesi, A.C.; Binelli, M.; et al. Silvopastoral system is an alternative to improve animal welfare and productive performance in meat production systems. Sci. Rep. 2021, 11, 14092. [Google Scholar] [CrossRef] [PubMed]
- Paiva, I.G.; Auad, A.M.; Veríssimo, B.A.; Silveira, L.C.P. Differences in the insect fauna associated to a monocultural pasture and a silvopasture in Southeastern Brazil. Sci. Rep. 2020, 10, 12112. [Google Scholar] [CrossRef] [PubMed]
- Salazar, R.; Alegre, J.; Pizarro, D.; Duff, A.J.; García, C.; Gómez, C. Soil carbon stock potential in pastoral and silvopastoral systems in the Peruvian Amazon. Agrofor. Syst. 2024, 98, 2157–2167. [Google Scholar] [CrossRef]
- Dagar, J.C.; Gupta, S.R.; Teketay, D. Silvopasture options for enhanced biological productivity of degraded pasture/grazing lands: An overview. In Agroforestry for Degraded Landscapes: Recent Advances and Emerging Challenges; Springer: Singapore, 2020; Volume 2, pp. 163–227. [Google Scholar] [CrossRef]
- Navarro, E.F.; Gómez, C.; Pizarro, D.; Alegre, J.; Castillo, M.S.; Vela, J.; Vásquez, H. Development of silvopastoral systems in the Peruvian Amazon. In Silvopastoral Systems of Meso America and Northern South America; Springer: Cham, Switzerland, 2023; pp. 135–154. [Google Scholar] [CrossRef]
- Dos Reis, J.C.; Kamoi, M.Y.T.; Michetti, M.; Wruck, F.J.; Rodrigues-Filho, S. Sistema de integração lavoura-pecuária-floresta como estratégia de desenvolvimento sustentável no estado de Mato Grosso. In Repositório de Casos sobre o Big Push para a Sustentabilidade no Brasil; CEPAL: Santiago, Chile, 2020; Available online: https://biblioguias.cepal.org/bigpushparaasustentabilidade (accessed on 21 March 2025).
- Arevalo, L.A.; Alegre, J.C.; Bandy, D.E.; Szott, L.T. The effect of cattle grazing on soil physical and chemical properties in a silvopastoral system in the Peruvian Amazon. Agrofor. Syst. 1998, 40, 109–124. [Google Scholar] [CrossRef]
- Valqui, L.; Lopez, E.L.; Lopez, C.A.; Valqui-Valqui, L.; Bobadilla, L.G.; Vigo, C.N.; Vásquez, H.V. Influence of the arboreal component in the productive and nutritional parameters of Brachiaria mutica grass in northeastern Peru. Environ. Sci. Proc. 2022, 22, 69. [Google Scholar] [CrossRef]
- Pizarro, D.; Vásquez, H.; Bernal, W.; Fuentes, E.; Alegre, J.; Castillo, M.S.; Gómez, C. Assessment of silvopasture systems in the northern Peruvian Amazon. Agrofor. Syst. 2020, 94, 173–183. [Google Scholar] [CrossRef]
- Sanginga, N.; Mulongoy, K.; Ayanaba, A. Nitrogen fixation of field-inoculated Leucaena leucocephala (Lam.) de Wit estimated by the 15N and the difference methods. Plant Soil 1989, 117, 269–274. [Google Scholar] [CrossRef]
- Liyanage, M.d.S.; Danso, S.K.A.; Jayasundara, H.P.S. Biological nitrogen fixation in four Gliricidia sepium genotypes. Plant Soil 1994, 161, 267–274. [Google Scholar] [CrossRef]
- Cervantes, C.A. Nitrogen management in coffee–legume agroforestry systems in Costa Rica. In Management of Agroforestry Systems for Enhancing Resource Use Efficiency and Crop Productivity; International Atomic Energy Agency: Vienna, Austria, 2008; p. 175. [Google Scholar]
- Chará-Serna, A.M.; Chará, J. Efecto de los sistemas silvopastoriles sobre la biodiversidad y la provisión de servicios ecosistémicos en agropaisajes tropicales. Livest. Res. Rural Dev. 2020, 32, 184. Available online: http://www.lrrd.org/lrrd32/11/ana32184.html (accessed on 22 February 2025).
- Vásquez, H.V.; Valqui, L.; Bobadilla, L.G.; Arbizu, C.I.; Alegre, J.C.; Maicelo, J.L. Influence of arboreal components on the physical-chemical characteristics of the soil under four silvopastoral systems in northeastern Peru. Heliyon 2021, 7, e07725. [Google Scholar] [CrossRef]
- Leblanc, H.A.; McGraw, R.L.; Nygren, P.; Roux, C.L. Neotropical legume tree Inga edulis forms N2-fixing symbiosis with fast-growing Bradyrhizobium strains. Plant Soil 2005, 275, 123–133. [Google Scholar] [CrossRef]
- Leblanc, H.A.; Nygren, P.; McGraw, R.L. Green mulch decomposition and nitrogen release from leaves of two Inga spp. in an organic alley-cropping practice in the humid tropics. Soil Biol. Biochem. 2006, 38, 349–358. [Google Scholar] [CrossRef]
- Hernández, A.P.; Bautista, I.B.; Chávez, L.M.; Ramírez, Á.E.C.; Mateo, A.B. Sistemas silvopastoriles para la producción de rumiantes en pastoreo. Braz. J. Dev. 2023, 9, 30956–30972. [Google Scholar] [CrossRef]
- Angadi, S.V.; Umesh, M.R.; Begna, S.; Gowda, P. Light interception, agronomic performance, and nutritive quality of annual forage legumes as affected by shade. Field Crops Res. 2022, 275, 108358. [Google Scholar] [CrossRef]
- Pang, K.; Van Sambeek, J.W.; Navarrete-Tindall, N.E.; Lin, C.H.; Jose, S.; Garrett, H.E. Responses of legumes and grasses to non-, moderate, and dense shade in Missouri, USA. I. Forage yield and its species-level plasticity. Agrofor. Syst. 2019, 93, 11–24. [Google Scholar] [CrossRef]
- Mercier, K.M.; Teutsch, C.D.; Fike, J.H.; Munsell, J.F.; Tracy, B.F.; Strahm, B.D. Impact of increasing shade levels on the dry-matter yield and botanical composition of multispecies forage stands. Grass Forage Sci. 2020, 75, 291–302. [Google Scholar] [CrossRef]
- De Oliveira, A.F.; Menezes, G.L.; Goncalves, L.C.; de Araujo, V.E.; Ramirez, M.A.; Júnior, R.G.; Jayme, D.G.; Lana, A.M.Q. Pasture traits and cattle performance in silvopastoral systems with Eucalyptus and Urochloa: Systematic review and meta-analysis. Livest. Sci. 2022, 262, 104973. [Google Scholar] [CrossRef]
- Sarmin, I.J.; Rahman, M.S.; Amin, M.H.; Ahmed, K. Effects of bark and stem exudates of eucalyptus on three crop plants. J. Sci. Technol. 2020, 17, 24. [Google Scholar]
- Bosi, C.; Pezzopane, J.R.M.; Sentelhas, P.C.; Santos, P.M.; Nicodemo, M.L.F. Productivity and biometric characteristics of signal grass in a silvopastoral system. Pesqui. Agropecu. Bras. 2014, 49, 449–456. [Google Scholar] [CrossRef]
- Stewart, A.; Coble, A.; Contosta, A.R.; Orefice, J.N.; Smith, R.G.; Asbjornsen, H. Forest conversion to silvopasture and open pasture: Effects on soil hydraulic properties. Agrofor. Syst. 2020, 94, 869–879. [Google Scholar] [CrossRef]
- Fuentes, E.; Gómez, C.; Pizarro, D.; Alegre, J.; Castillo, M.; Vela, J.; Huaman, E.; Vásquez, H. A review of silvopastoral systems in the Peruvian Amazon region. Trop. Grassl.-Forraj. Trop. 2022, 10, 78–88. [Google Scholar] [CrossRef]
- Lima, M.A.; Paciullo, D.S.; Morenz, M.J.; Gomide, C.A.; Rodrigues, R.A.; Chizzotti, F.H. Productivity and nutritive value of Brachiaria decumbens and performance of dairy heifers in a long-term silvopastoral system. Grass Forage Sci. 2019, 74, 160–170. [Google Scholar] [CrossRef]
- Guamán-Rivera, S.A.; Herrera-Feijoo, R.J.; Velepucha-Caiminagua, H.J.; Avalos-Peñafiel, V.G.; Aguilar-Miranda, G.J.; Melendres-Medina, E.M.; Baquero-Tapia, M.F.; Cajamarca Carrazco, D.I.; Fernández-Vinueza, D.F.; Montero-Arteaga, A.A.; et al. Silvopastoral systems as a tool for recovering degraded pastures and improving animal thermal comfort indexes in Northern Ecuador. Braz. J. Biol. 2024, 84, e286137. [Google Scholar] [CrossRef] [PubMed]
- Sunariyo, S.; Firdausi, R.Z. Community-based agro-silvopastoral systems: Integrating forestry, agriculture, and livestock for sustainable rural development in forest regions. Serunai 2024, 4, 45–57. [Google Scholar] [CrossRef]
- SENAMHI. Servicio Nacional de Meteorología e Hidrología del Perú. 2018. Available online: https://www.senamhi.gob.pe/?p=pronostico-meteorologico (accessed on 21 March 2025).
- Irigoin, V.; Pizarro, D.; Fuentes, E.; García, C.; Wattiaux, M.; Picasso, V.; Arango, J.; Romero, G.; Gómez-Bravo, C. Biotechnical, economic, and environmental assessment of dairy systems in the Peruvian Amazon utilizing the CLEANED tool. Agrofor. Syst. 2025, 99, 32. [Google Scholar] [CrossRef]
- Díaz Pablo, M.E.; Alegre Orihuela, J.C.; Gómez Bravo, C.A.; Mendoza Tamani, P.; Arévalo-Hernández, C.O. Reservas de carbono en tres sistemas silvopastoriles de la Amazonía peruana. Manglar 2024, 21, 305–311. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2001; Methods 950.46, 2001.11. [Google Scholar]
- Tilley, J.M.A.; Terry, D.R. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- ANKOM. In Vitro Digestibility using the Daisy II Incubator. 2017. Available online: https://www.ankom.com/sites/default/files/documentfiles/Method_3_Invitro_D200_D200I.pdf (accessed on 21 March 2025).
- Indah, A.S.; Permana, I.G.; Despal, D. Correlation and determination of the metabolizable energy (ME) of tropical forage with nutrient content for ruminants. Aceh J. Anim. Sci. 2023, 8, 34–38. [Google Scholar] [CrossRef]
- Blake, G.R. Bulk density. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Properties, Including Statistics of Measurement and Sampling; American Society of Agronomy: Madison, WI, USA, 1965; Volume 9, pp. 374–390. [Google Scholar]
- Danielson, R.E.; Sutherland, P.L. Porosity. In Methods of Soil Analysis: Part 1 Physical and Mineralogical Methods; American Society of Agronomy: Madison, WI, USA, 1986; Volume 5, pp. 443–461. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Rhoades, J.D. Cation exchange capacity. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Ed.; ASA and SSSA: Madison, WI, USA, 1982; Volume 9, pp. 149–157. [Google Scholar] [CrossRef]
- De Freitas, A.F.; Junqueira Carneiro, J.; Venturin, N.; de Souza Moreira, F.M.; Guimaraes Ferreira, A.I.; de Oliveira Lara, G.P.; Cardoso, I.M. Inga edulis Mart. intercropped with pasture improves soil quality without compromising forage yields. Agrofor. Syst. 2020, 94, 2355–2366. [Google Scholar] [CrossRef]
- Alegre, J.C.; Weber, J.C.; Bandy, D.E. The potential of Inga species for improved woody fallows and multistrata agroforest in the Peruvian Amazon Basin. In The Genus Inga—Utilization; Pennington, T.D., Fernández, E.C.M., Eds.; The Royal Botanic Garden: Kew, UK, 1998; Chapter 6; pp. 87–100. [Google Scholar]
- Nichols, J.D.; Carpenter, F.L. Interplanting Inga edulis yields nitrogen benefits to Terminalia amazonia. For. Ecol. Manag. 2006, 233, 344–351. [Google Scholar] [CrossRef]
- da Silva, E.R.; da Costa Ayres, M.I.; Neves, A.L.; Uguen, K.; de Oliveira, L.A.; Alfaia, S.S. Organic fertilization with residues of cupuassu (Theobroma grandiflorum) and inga (Inga edulis) for improving soil fertility in central Amazonia. In New Generation of Organic Fertilizers; IntechOpen: London, UK, 2021. [Google Scholar]
- Soper, F.M.; Sparks, J.P. Estimating ecosystem nitrogen addition by a leguminous tree: A mass balance approach using a woody encroachment chronosequence. Ecosystems 2017, 20, 1164–1178. [Google Scholar] [CrossRef]
- Van Chuong, N. Effect of three different nitrogen rates and three rhizosphere N2-fixing bacteria on growth, yield and quality of peanuts. Trends Sci. 2024, 21, 7281. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic interactions and allelochemicals: New possibilities for sustainable weed management. Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Harper, K.J.; McNeill, D.M. The role iNDF in the regulation of feed intake and the importance of its assessment in subtropical ruminant systems (the role of iNDF in the regulation of forage intake). Agriculture 2015, 5, 778–790. [Google Scholar] [CrossRef]
- Baldissera, T.C.; da Silveira Pontes, L.; Giostri, A.F.; Barro, R.S.; Lustosa, S.B.C.; de Moraes, A.; de Faccio Carvalho, P.C. Sward structure and relationship between canopy height and light interception for tropical C₄ grasses growing under trees. Crop Pasture Sci. 2016, 67, 1199–1207. [Google Scholar] [CrossRef]
- Banakar, P.S.; Anand Kumar, N.; Shashank, C.G. Physically effective fiber in ruminant nutrition. J. Pharmacogn. Phytochem. 2018, 7, 303–308. [Google Scholar]
- Reinbothe, S.; Reinbothe, C. The regulation of enzymes involved in chlorophyll biosynthesis. Eur. J. Biochem. 1996, 237, 323–343. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lojka, B.; Preininger, D.; Van Damme, P.; Rollo, A.; Banout, J. Use of the Amazonian tree species Inga edulis for soil regeneration and weed control. J. Trop. For. Sci. 2012, 24, 89–101. [Google Scholar]
- Zhang, X.; Zhao, Q.; Wei, L.M.; Sun, Q.Y.; Zeng, D.H. Tree roots exert greater impacts on phosphorus fractions than aboveground litter in mineral soils under a Pinus sylvestris var. mongolica plantation. For. Ecol. Manag. 2023, 545, 121242. [Google Scholar] [CrossRef]
- Gaind, S. Phosphorus mobilization strategies of grain legumes: An overview. J. Adv. Microbiol. 2017, 3, 1–15. [Google Scholar] [CrossRef]
- Lebrazi, S.; Fikri-Benbrahim, K. Potential of tree legumes in agroforestry systems and soil conservation. In Advances in Legumes for Sustainable Intensification; Academic Press: Cambridge, MA, USA, 2022; pp. 461–482. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).