
Lymphovascular Tumoral Emboli in Inflammatory Breast Cancer Result from Haptotaxis-Mediated Encircling Lymphangiogenesis


Abstract
1. Introduction
2. Results
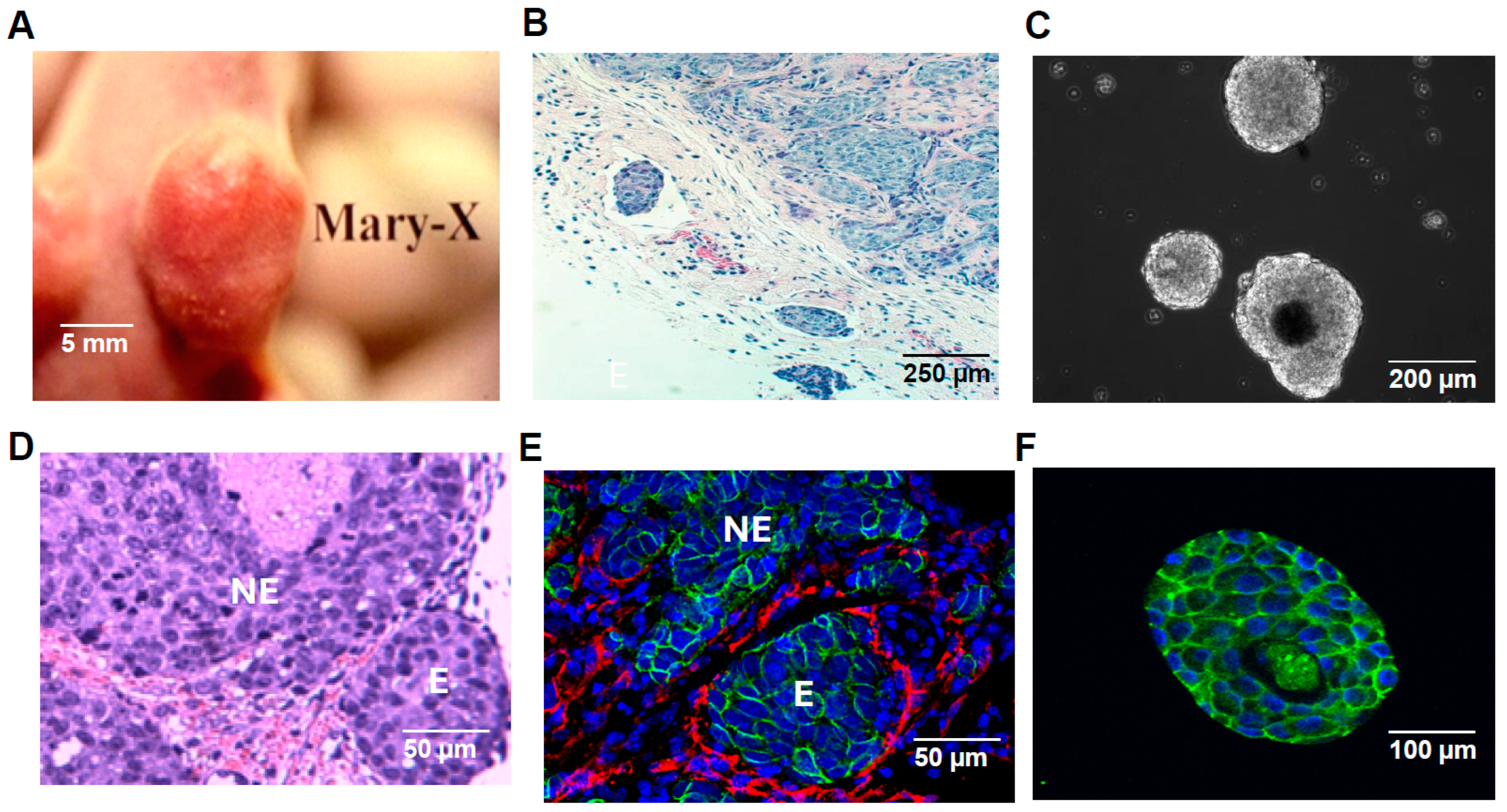
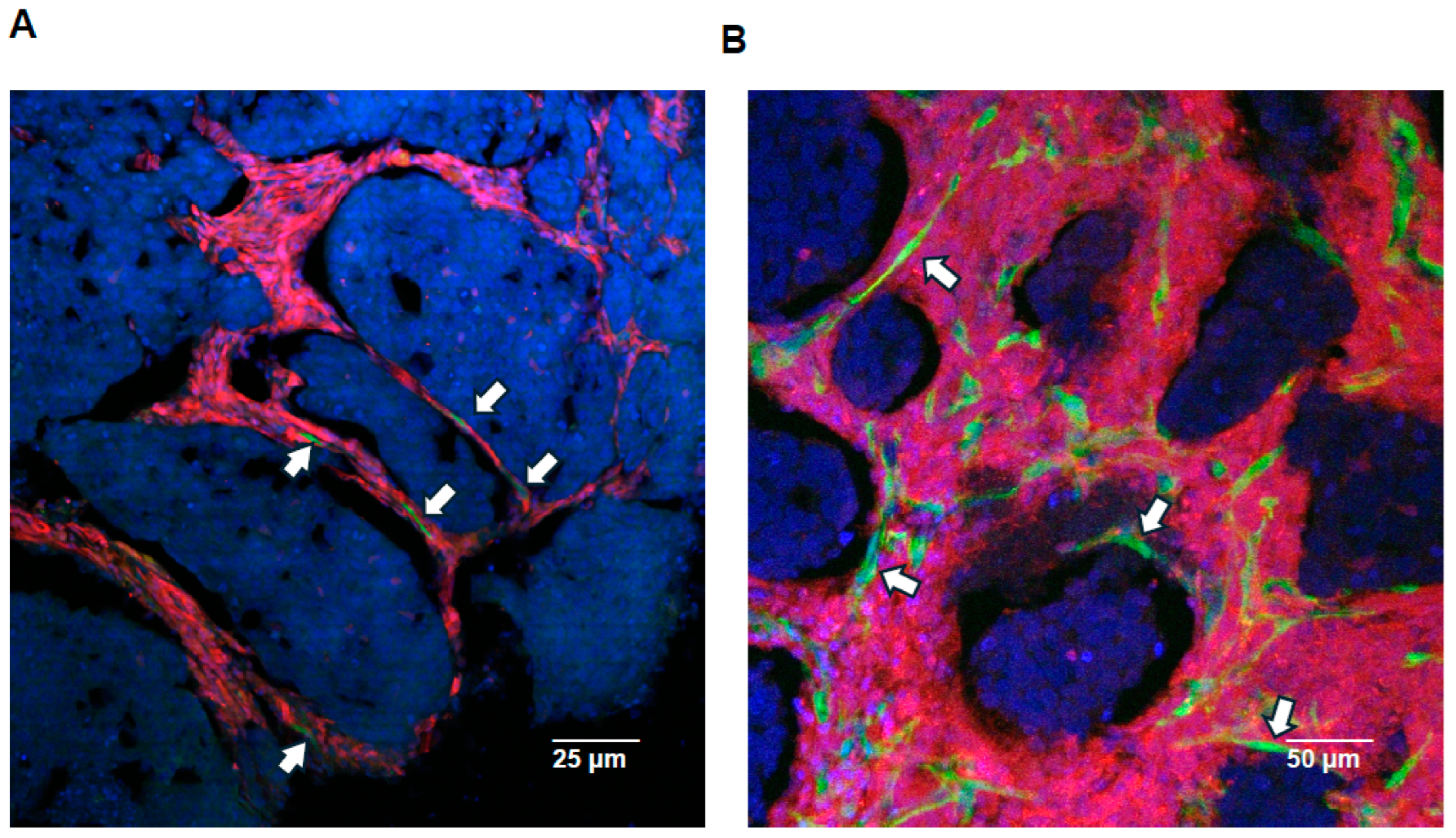
2.1. Formation and Characterization of IBC Emboli in Lymphatic Vessels
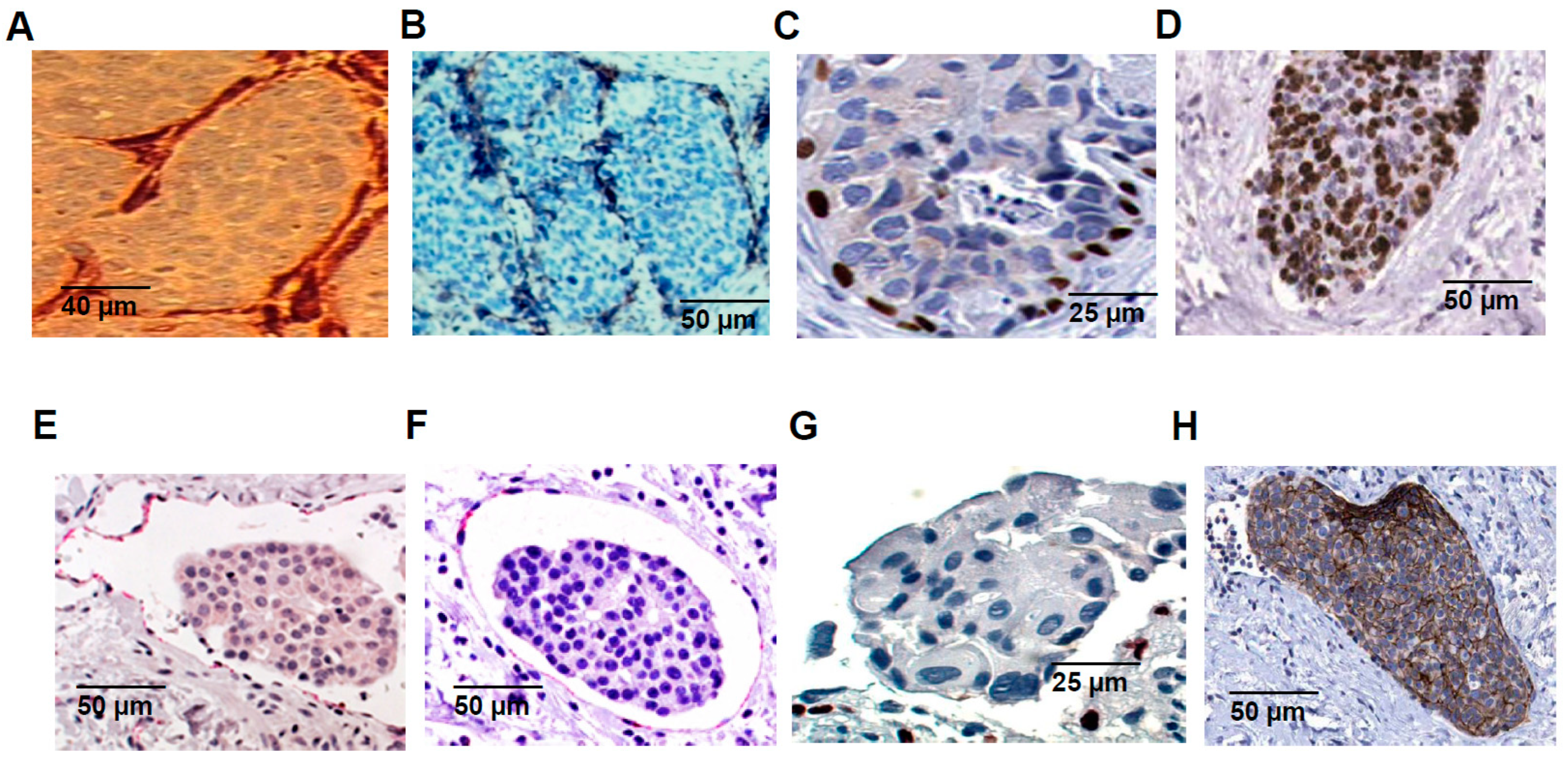
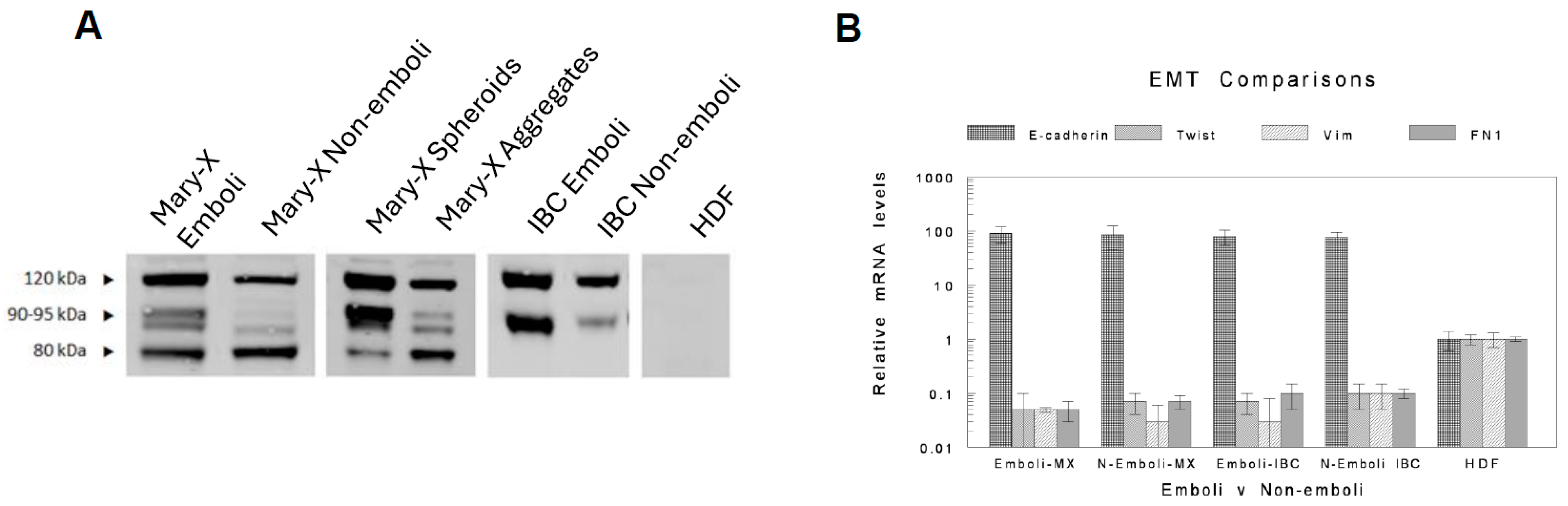
2.2. In Vivo Embolic vs. Non-Embolic Comparisons of E-Cadherin Expression
2.3. In Vitro Lymphovasculogenesis Growth Factor Comparisons
2.4. In Vitro Vascular and Lymphatic Endothelial Haptotaxis/Chemotaxis Comparisons of Mary-X Spheroid Conditioned Media (CM) vs. Other Breast Carcinoma Lines
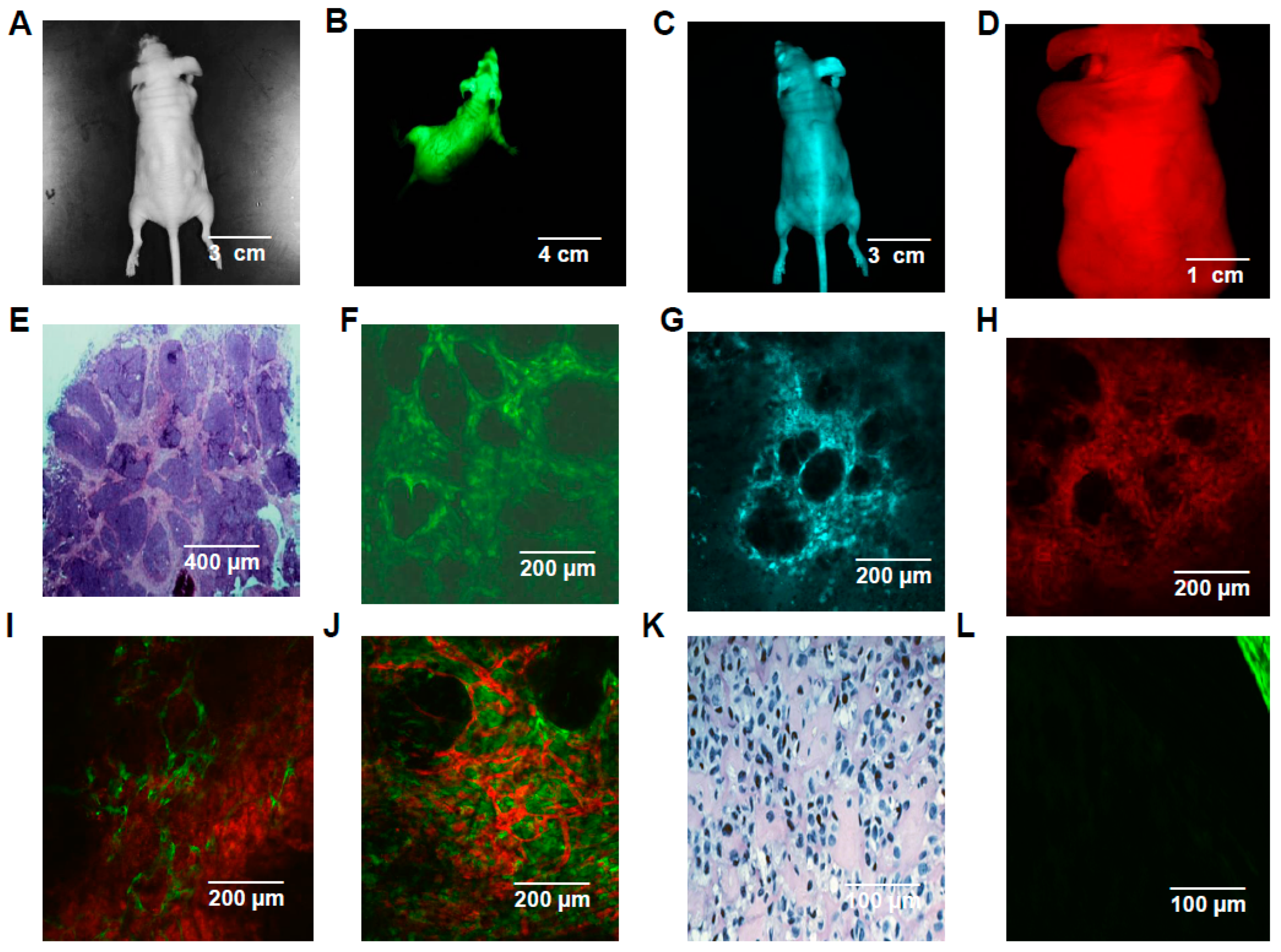
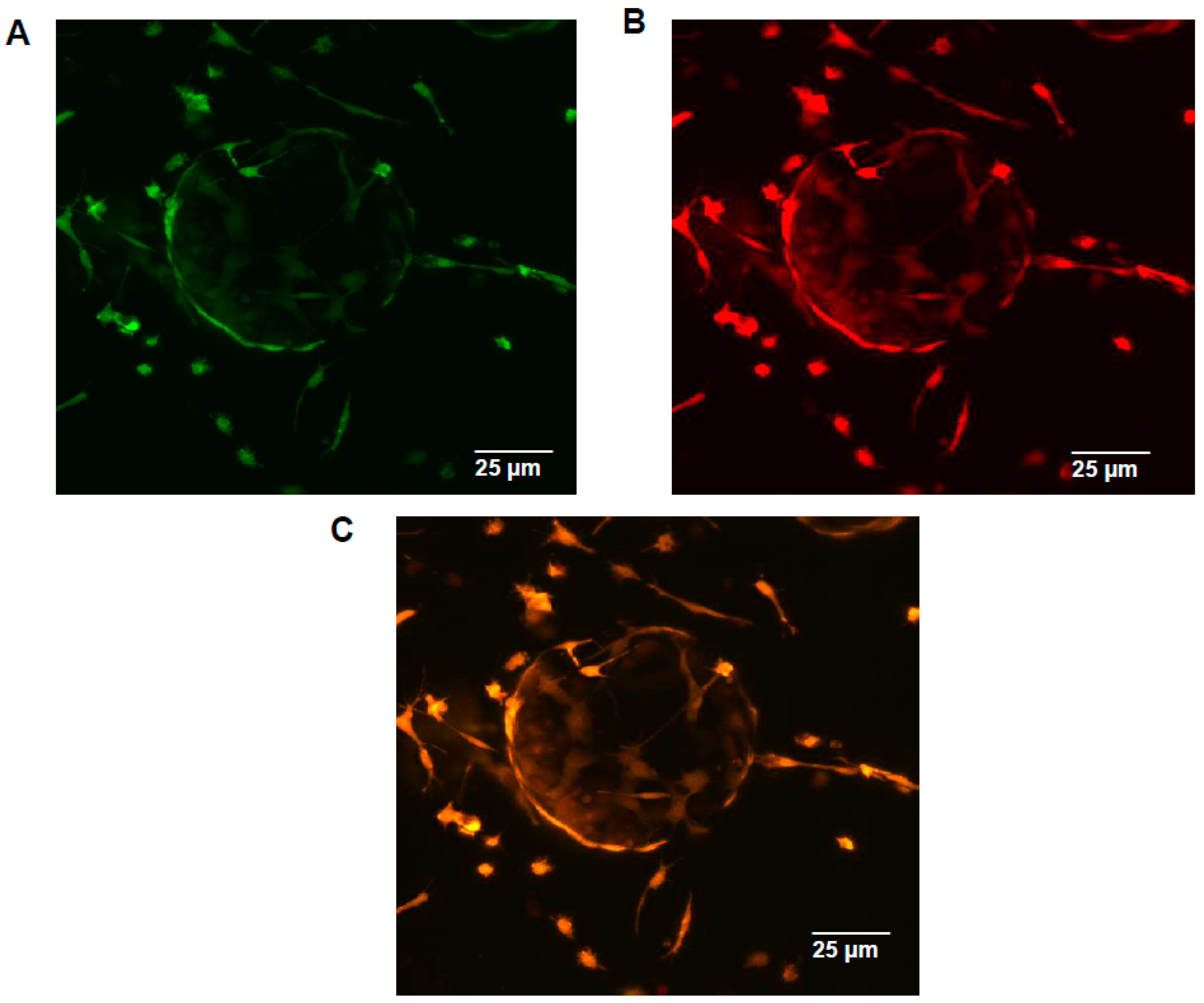
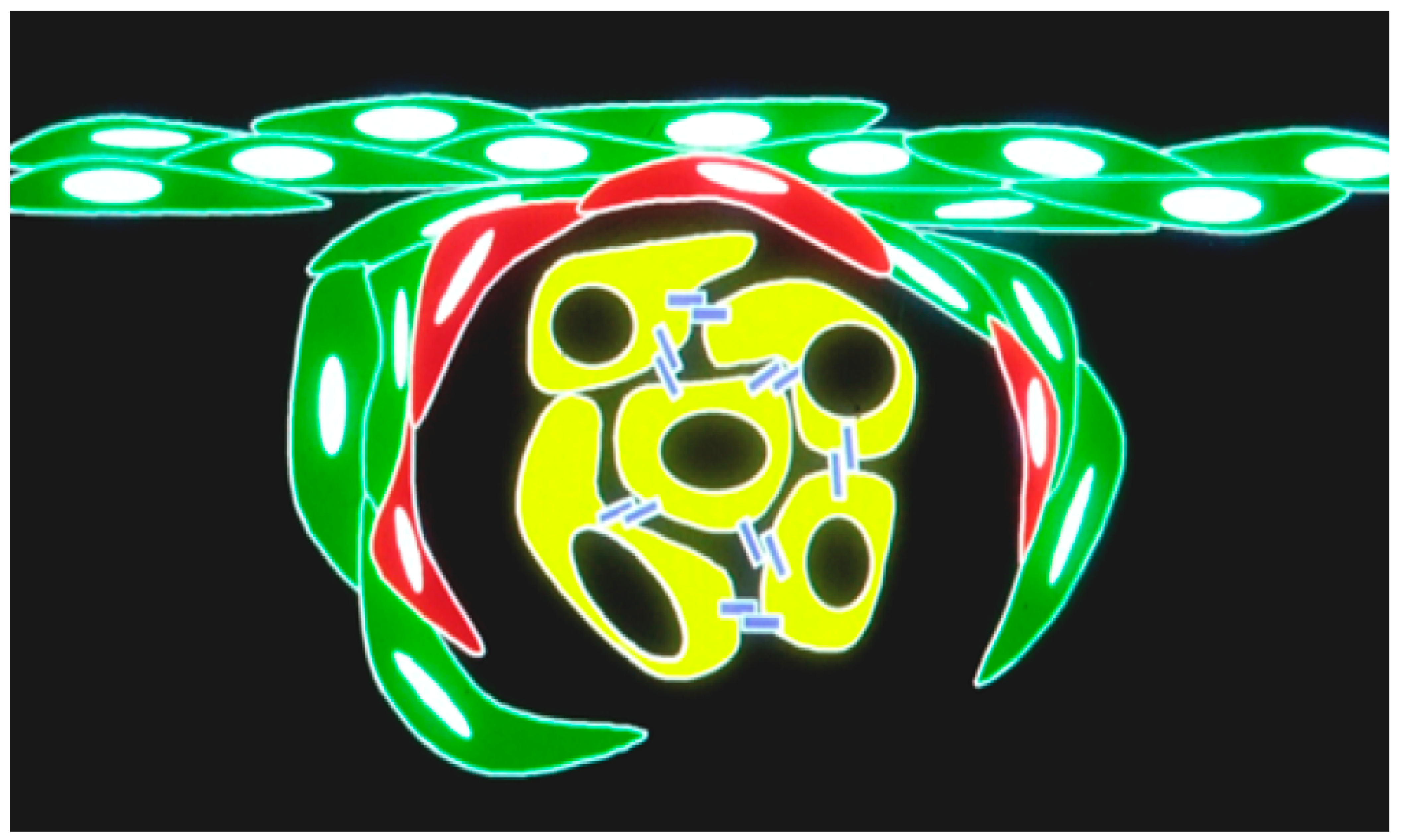
2.5. In Vivo/In Vitro Spectrally Distinct Transgenic Reporter Gene Studies
3. Discussion
4. Materials and Methods
4.1. Institutional Approvals
4.2. Emboli vs. Non-Emboli Comparisons
Immunohistochemistry
4.3. ATCC Patent Deposits and Cell Identification
4.4. Confocal Single and Double Label Immunofluorescence Experiments on Mary-X and Mary-X Spheroids
4.5. Laser Capture Microdissection
4.6. Preparation of Protein Lysates and Western Blot Analysis
4.7. In Vitro Studies with Mary-X Spheroids and Other Breast Cancer Cell Lines
4.8. EMT and Growth Factor Comparisons
4.9. Vascular and Lymphatic Endothelial Haptotaxis/Chemotaxis Comparisons
4.10. In Vivo/In Vitro Transgenic Reporter Imaging Comparisons
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brat, D.J.; Hruban, R.H. A metastasis caught in the act. N. Engl. J. Med. 1996, 335, 1733. [Google Scholar] [CrossRef] [PubMed]
- Alpaugh, M.L.; Tomlinson, J.S.; Shao, Z.M.; Barsky, S.H. A novel human xenograft model of inflammatory breast cancer. Cancer Res. 1999, 59, 5079–5084. [Google Scholar]
- Tomlinson, J.S.; Alpaugh, M.L.; Barsky, S.H. An intact overexpressed E-cadherin/α, β catenin axis characterizes the lymphovascular emboli of inflammatory breast carcinoma. Cancer Res. 2001, 61, 5231–5241. [Google Scholar]
- Modi, A.P.; Nguyen, J.P.T.; Wang, J.; Ahn, J.S.; Libling, W.A.; Klein, J.M.; Mazumder, P.; Barsky, S.H. Geometric tumor embolic budding characterizes inflammatory breast cancer. Breast Cancer Res. Treat. 2023, 197, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Houvenaeghel, G.; Cohen, M.; Classe, J.M.; Reyal, F.; Mazouni, C.; Chopin, N.; Martinez, A.; Daraï, E.; Coutant, C.; Colombo, P.E.; et al. Lymphovascular invasion has a significant prognostic impact in patients with early breast cancer, results from a large, national, multicenter, retrospective cohort study. ESMO Open 2021, 6, 100316. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, B.L.; Gagliardi, M.; Iles, L.K.; Evans, K.; Ivan, C.; Liu, X.; Liu, C.G.; Souza, G.; Rao, A.; Meric-Bernstam, F.; et al. Clinically relevant inflammatory breast cancer patient-derived xenograft–derived ex vivo model for evaluation of tumor-specific therapies. PLoS ONE 2018, 13, 1–23. [Google Scholar] [CrossRef]
- Cristofanilli, M.; Valero, V.; Buzdar, A.U.; Kau, S.W.; Broglio, K.R.; Gonzalez-Angulo, A.M.; Sneige, N.; Islam, R.; Ueno, N.T.; Buchholz, T.A.; et al. Inflammatory breast cancer (IBC) and patterns of recurrence: Understanding the biology of a unique disease. Cancer 2007, 110, 1436–1444. [Google Scholar] [CrossRef]
- Ye, Y.; Tian, H.; Lange, A.R.; Yearsley, K.; Robertson, F.M.; Barsky, S.H. The genesis and unique properties of the lymphovascular tumor embolus are because of calpain-regulated proteolysis of E-cadherin. Oncogene 2013, 32, 1702–1713. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, W.; Izban, M.G.; Ballard, B.R.; Barsky, S.H. Initiation of tumor dormancy by the lymphovascular embolus. Oncotarget, 2024; in press. [Google Scholar]
- Bronsert, P.; Enderle-Ammour, K.; Bader, M.; Timme, S.; Kuehs, M.; Csanadi, A.; Kayser, G.; Kohler, I.; Bausch, D.; Hoeppner, J.; et al. Cancer cell invasion and EMT marker expression: A three-dimensional study of the human cancer-host interface. J. Pathol. 2014, 234, 410–422. [Google Scholar] [CrossRef]
- Grigore, A.; Jolly, M.; Jia, D.; Farach-Carson, M.; Levine, H. Tumor Budding: The Name is EMT. Partial EMT. J. Clin. Med. 2016, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Sternlicht, M.D.; Safarians, S.; Calcaterra, T.C.; Barsky, S.H. Establishment and characterization of a novel human myoepithelial cell line and matrix-producing xenograft from a parotid basal cell adenocarcinoma. Vitr. Cell Dev. Biol. Anim. 1996, 32, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Sternlicht, M.D.; Safarians, S.; Rivera, S.P.; Barsky, S.H. Characterizations of the extracellular matrix and proteinase inhibitor content of human myoepithelial tumors. Lab. Investig. 1996, 74, 781–796. [Google Scholar]
- Sun, H.; Cao, S.; Mashl, R.J.; Mo, C.K.; Zaccaria, S.; Wendl, M.C.; Davies, S.R.; Bailey, M.H.; Primeau, T.M.; Hoog, J.; et al. Comprehensive characterization of 536 patient-derived xenograft models prioritizes candidates for targeted treatment. Nat. Commun. 2021, 12, 5086. [Google Scholar] [CrossRef]
- Villodre, E.S.; Hu, X.; Larson, R.; Finetti, P.; Gomez, K.; Balema, W.; Stecklein, S.R.; Santiago-Sanchez, G.; Krishnamurthy, S.; Song, J.; et al. Lipocalin 2 promotes inflammatory breast cancer tumorigenesis and skin invasion. Mol. Oncol. 2021, 15, 2752–2765. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.; Lebleu, V.S.; Kalluri, R. Biology C: EMT Program is Dispensable for Metastasis but Induces Chemoresistance in Pancreatic Cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef]
- Amoh, Y.; Bouvet, M.; Li, L.; Tsuji, K.; Moossa, A.R.; Katsuoka, K.; Hoffman, R.M. Visualization of nascent tumor angiogenesis in lung and liver metastasis by differential dual-color fluorescence imaging in nestin-linked-GFP mice. Clin. Exp. Metastasis 2006, 23, 315–322. [Google Scholar] [CrossRef]
- Xu, C.; Li, X.; Liu, P.; Li, M.; Luo, F. Patient-derived xenograft mouse models: A high fidelity tool for individualized medicine. Oncol. Lett. 2019, 17, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Ye, Y.; Yearsley, K.; Jones, S.; Barsky, S.H. The lymphovascular embolus of inflammatory breast cancer expresses a stem cell-like phenotype. Am. J. Pathol. 2008, 173, 561–574. [Google Scholar] [CrossRef]
- Ye, X.; Gaucher, J.F.; Vidal, M.; Broussy, S. A Structural Overview of Vascular Endothelial Growth Factors Pharmacological Ligands: From Macromolecules to Designed Peptidomimetics. Molecules 2021, 26, 6759. [Google Scholar] [CrossRef]
- SenGupta, S.; Parent, C.A.; Bear, J.E. The principles of directed cell migration. Nat. Rev. Mol. Cell. Biol. 2021, 22, 529–547. [Google Scholar] [CrossRef]
- Popławski, P.; Zarychta-Wiśniewska, W.; Burdzińska, A.; Bogusławska, J.; Adamiok-Ostrowska, A.; Hanusek, K.; Rybicka, B.; Białas, A.; Kossowska, H.; Iwanicka-Nowicka, R.; et al. Renal cancer secretome induces migration of mesenchymal stromal cells. Stem Cell Res. Ther. 2023, 14, 200. [Google Scholar] [CrossRef]
- Suetsugu, A.; Katz, M.; Fleming, J.; Truty, M.; Thomas, R.; Moriwaki, H.; Bouvet, M.; Saji, S.; Hoffman, R.M. Multi-color palette of fluorescent proteins for imaging the tumor microenvironment of orthotopic tumorgraft mouse models of clinical pancreatic cancer specimens. J. Cell Biochem. 2012, 113, 2290–2295. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, A.; Katz, M.; Fleming, J.; Truty, M.; Thomas, R.; Saji, S.; Moriwaki, H.; Bouvet, M.; Hoffman, R.M. Imageable fluorescent metastasis resulting in transgenic GFP mice orthotopically implanted with human-patient primary pancreatic cancer specimens. Anticancer. Res. 2012, 32, 1175–1180. [Google Scholar] [PubMed]
- Hiroshima, Y.; Zhao, M.; Maawy, A.; Zhang, Y.; Katz, M.H.; Fleming, J.B.; Uehara, F.; Miwa, S.; Yano, S.; Momiyama, M.; et al. Hoffman RM Efficacy of Salmonella typhimurium A1-R versus chemotherapy on a pancreatic cancer patient-derived orthotopic xenograft (PDOX). J. Cell Biochem. 2014, 115, 1254–1261. [Google Scholar] [CrossRef]
- Suetsugu, A.; Katz, M.; Fleming, J.; Truty, M.; Thomas, R.; Saji, S.; Moriwaki, H.; Bouvet, M.; Hoffman, R.M. Non-invasive fluorescent-protein imaging of orthotopic pancreatic-cancer-patient tumorgraft progression in nude mice. Anticancer Res. 2012, 32, 3063–3067. [Google Scholar]
- Suetsugu, A.; Hassanein, M.K.; Reynoso, J.; Osawa, Y.; Nagaki, M.; Moriwaki, H.; Saji, S.; Bouvet, M.; Hoffman, R.M. The cyan fluorescent protein nude mouse as a host for multicolor-coded imaging models of primary and metastatic tumor microenvironments. Anticancer Res. 2012, 32, 31–38. [Google Scholar]
- Mii, S.; Duong, J.; Tome, Y.; Uchugonova, A.; Liu, F.; Amoh, Y.; Saito, N.; Katsuoka, K.; Hoffman, R.M. The role of hair follicle nestin-expressing stem cells during whisker sensory-nerve growth in long-term 3D culture. J. Cell. Biochem. 2013, 114, 1674–1684. [Google Scholar] [CrossRef] [PubMed]
- Amoh, Y.; Li, L.; Yang, M.; Moossa, A.R.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Nascent blood vessels in the skin arise from nestin-expressing hair-follicle cells. Proc. Natl. Acad. Sci. USA 2004, 101, 13291–13295. [Google Scholar] [CrossRef]
- Hoffman, R.M. Nestin-driven green fluorescent protein as an imaging marker for nascent blood vessels in mouse models of cancer. Methods Mol. Biol. 2011, 689, 183–204. [Google Scholar] [CrossRef]
- Koning, J.J.; Konijn, T.; Lakeman, K.A.; O’Toole, T.; Kenswil, K.J.; Raaijmakers, M.H.; Michurina, T.V.; Enikolopov, G.; Mebius, R.E. Nestin-Expressing Precursors Give Rise to Both Endothelial as well as Nonendothelial Lymph Node Stromal Cells. J. Immunol. 2016, 197, 2686–2694. [Google Scholar] [CrossRef] [PubMed]
- Mahooti, S.; Porter, K.; Alpaugh, M.L.; Ye, Y.; Xiao, Y.; Jones, S.; Tellez, J.D.; Barsky, S.H. Breast carcinomatous tumoral emboli can result from encircling lymphovasculogenesis rather than lymphovascular invasion. Oncotarget 2010, 1, 131–147. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Assen, F.P.; Leithner, A.; Abe, J.; Schachner, H.; Asfour, G.; Bago-Horvath, Z.; Stein, J.V.; Uhrin, P.; Sixt, M.; et al. Lymph node blood vessels provide exit routes for metastatic tumor cell dissemination in mice. Science 2018, 359, 1408–1411. [Google Scholar] [CrossRef]
- Pereira, E.R.; Kedrin, D.; Seano, G.; Gautier, O.; Meijer, E.F.J.; Jones, D.; Chin, S.-M.; Kitahara, S.; Bouta, E.M.; Chang, J.; et al. Lymph node metastases can invade local blood vessels, exit the node, and colonize distant organs in mice. Science 2018, 359, 1403–1407. [Google Scholar] [CrossRef]
- Giuliano, A.E.; Ballman, K.V.; McCall, L.; Beitsch, P.D.; Brennan, M.B.; Kelemen, P.R.; Ollila, D.W.; Hansen, N.M.; Whitworth, P.W.; Blumencranz, P.W.; et al. Effect of Axillary Dissection vs No Axillary Dissection on 10-Year Overall Survival Among Women with Invasive Breast Cancer and Sentinel Node Metastasis: The ACOSOG Z0011 (Alliance) Randomized Clinical Trial. JAMA 2017, 318, 918–926. [Google Scholar] [CrossRef]
- Barsky, S.H.; Gentchev, L.; Basu, A.S.; Jimenez, R.E.; Boussaid, K.; Gholap, A.S. Use and validation of epithelial recognition and fields of view algorithms on virtual slides to guide TMA construction. Biotechniques 2009, 47, 927–938. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | VEGF | Vascular Haptotaxis | Lymphatic Haptotaxis |
|---|---|---|---|
| Mary-X | +++++ VEGF-C ++++ VEGF-D | +++++ | +++++ |
| 231 | +++ VEGF-C ++ VEGF-D | ++ | ++ |
| MCF-7 | ++ VEGF-C + VEGF-D | + | + |
| HMS-1 * | − VEGF-C − VEGF-D | −− | −− |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Hoffman, R.M.; Ye, Y.; Dillard, J.; Barsky, S.H. Lymphovascular Tumoral Emboli in Inflammatory Breast Cancer Result from Haptotaxis-Mediated Encircling Lymphangiogenesis. Lymphatics 2024, 2, 195-211. https://doi.org/10.3390/lymphatics2040016
Wang J, Hoffman RM, Ye Y, Dillard J, Barsky SH. Lymphovascular Tumoral Emboli in Inflammatory Breast Cancer Result from Haptotaxis-Mediated Encircling Lymphangiogenesis. Lymphatics. 2024; 2(4):195-211. https://doi.org/10.3390/lymphatics2040016
Chicago/Turabian StyleWang, Justin, Robert M. Hoffman, Yin Ye, Jordan Dillard, and Sanford H. Barsky. 2024. "Lymphovascular Tumoral Emboli in Inflammatory Breast Cancer Result from Haptotaxis-Mediated Encircling Lymphangiogenesis" Lymphatics 2, no. 4: 195-211. https://doi.org/10.3390/lymphatics2040016
APA StyleWang, J., Hoffman, R. M., Ye, Y., Dillard, J., & Barsky, S. H. (2024). Lymphovascular Tumoral Emboli in Inflammatory Breast Cancer Result from Haptotaxis-Mediated Encircling Lymphangiogenesis. Lymphatics, 2(4), 195-211. https://doi.org/10.3390/lymphatics2040016