
Self-Similarity and Spatial Periodicity in Cerebral Cortical Patterning: Structural Design Notes for Neural Tissue Architects

Abstract
1. Introduction
2. The Cortical Column Conundrum
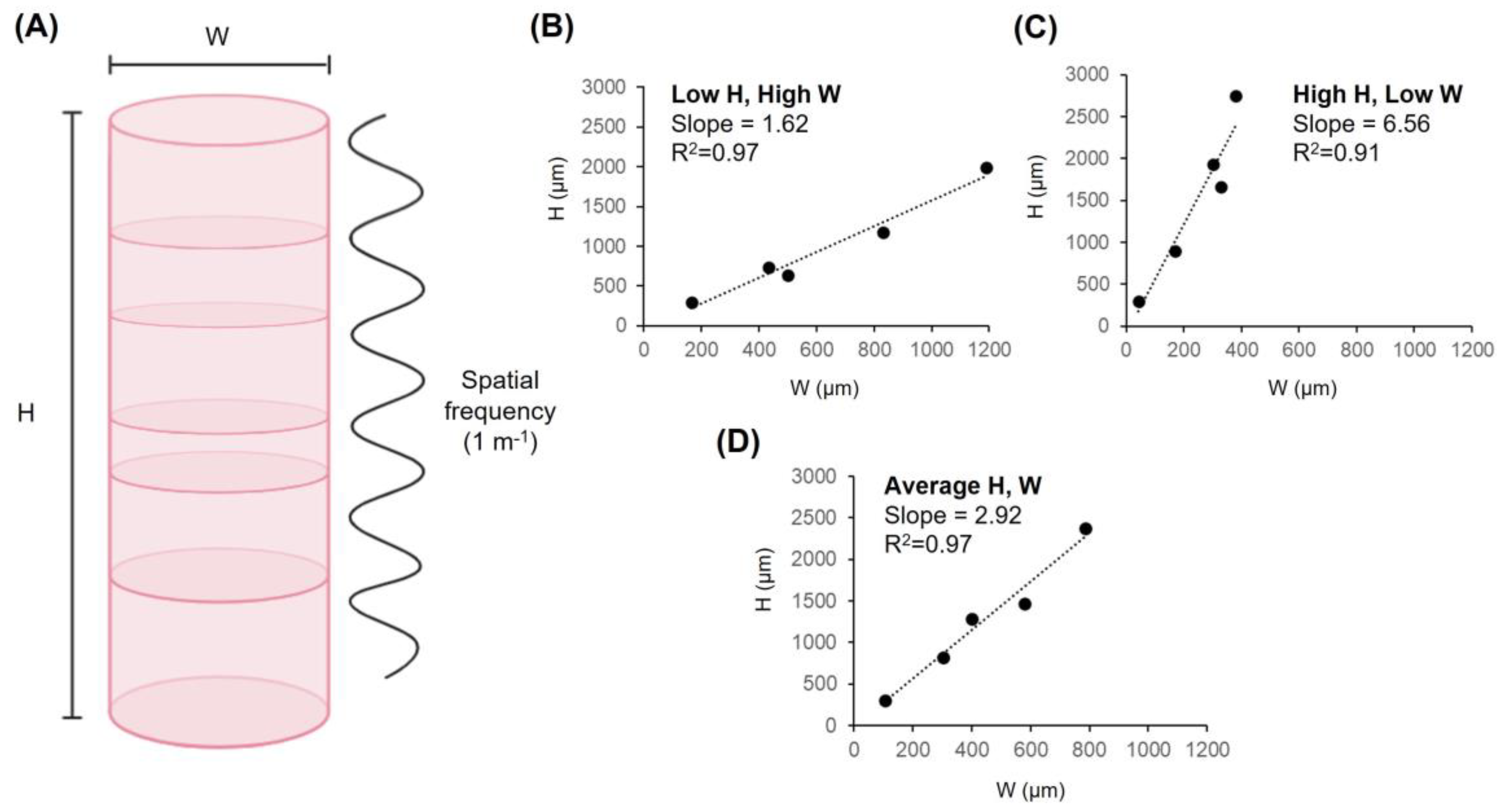
3. The Dimensions of Columns
4. Spatial Periodicities within Columns Predict Observed Laminar Organization
5. A Hypothesis of Cytoarchitectonic Self-Similarity and Periodicity
6. Implications for Neural Tissue Engineering
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gould, S.J.; Lewontin, R.C. The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1979, 205, 581–598. [Google Scholar]
- Buss, D.M.; Haselton, M.G.; Shackelford, T.K.; Bleske, A.L.; Wakefield, J.C. Adaptations, exaptations, and spandrels. Am. Psychol. 1998, 53, 533. [Google Scholar] [CrossRef] [PubMed]
- Raja, W.K.; Mungenast, A.E.; Lin, Y.T.; Ko, T.; Abdurrob, F.; Seo, J.; Tsai, L.H. Self-organizing 3D human neural tissue derived from induced pluripotent stem cells recapitulate Alzheimer’s disease phenotypes. PLoS ONE 2016, 11, e0161969. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, N.; Cantley, W.L.; Liaudanskaya, V.; Berk, A.; Du, C.; Rusk, W.; Peirent, E.; Koester, C.; Nieland, T.J.F.; Kaplan, D.L. A Long-Living Bioengineered Neural Tissue Platform to Study Neurodegeneration. Macromol. Biosci. 2020, 20, 2000004. [Google Scholar] [CrossRef]
- Tian, W.; Hou, S.; Ma, J.; Zhang, C.; Xu, Q.; Lee, I.; Li, H.; Spector, M.; Cui, F.; Chung, C.; et al. Hyaluronic acid–poly-D-lysine-based three-dimensional hydrogel for traumatic brain injury. Tissue Eng. 2005, 11, 513–525. [Google Scholar] [CrossRef]
- Zhang, T.; Yan, Y.; Wang, X.; Xiong, Z.; Lin, F.; Wu, R.; Zhang, R. Three-dimensional gelatin and gelatin/hyaluronan hydrogel structures for traumatic brain injury. J. Bioact. Compat. Polym. 2007, 22, 19–29. [Google Scholar] [CrossRef]
- Chung, B.G.; Kang, L.; Khademhosseini, A. Micro-and nanoscale technologies for tissue engineering and drug discovery applications. Expert Opin. Drug Discov. 2007, 2, 1653–1668. [Google Scholar] [CrossRef]
- Rouleau, N.; Cairns, D.M.; Rusk, W.; Levin, M.; Kaplan, D.L. Learning and synaptic plasticity in 3D bioengineered neural tissues. Neurosci. Lett. 2021, 750, 135799. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Tononi, G.; Edelman, G.M. Theoretical neuroanatomy and the connectivity of the cerebral cortex. Behav. Brain Res. 2002, 135, 69–74. [Google Scholar] [CrossRef]
- Bohm, D. A new theory of the relationship of mind and matter. Philos. Psychol. 1990, 3, 271–286. [Google Scholar] [CrossRef]
- Rouleau, N.; Murugan, N.J.; Kaplan, D.L. Toward Studying Cognition in a Dish. Trends Cogn. Sci. 2021, 25, 294–304. [Google Scholar] [CrossRef]
- Guy, J.; Wagener, R.J.; Möck, M.; Staiger, J.F. Persistence of functional sensory maps in the absence of cortical layers in the somsatosensory cortex of reeler mice. Cereb. Cortex 2015, 25, 2517–2528. [Google Scholar] [CrossRef]
- Guy, J.; Staiger, J.F. The functioning of a cortex without layers. Front. Neuroanat. 2017, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.H.; Lai, T.K.; Su, P.; Liu, F. Altered cortical Cytoarchitecture in the Fmr1 knockout mouse. Mol. Brain 2019, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.M.; Rector, D.M.; Roy, S.; Van Dongen, H.P.; Belenky, G.; Panksepp, J. Sleep as a fundamental property of neuronal assemblies. Nat. Rev. Neurosci. 2008, 9, 910–919. [Google Scholar] [CrossRef]
- Horton, J.C.; Adams, D.L. The cortical column: A structure without a function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 837–862. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. Confusing cortical columns. Proc. Natl. Acad. Sci. USA 2008, 105, 12099–12100. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Boone, K.; Tuszyński, J.; Barclay, P.; Simon, C. Possible existence of optical communication channels in the brain. Sci. Rep. 2016, 6, 36508. [Google Scholar] [CrossRef]
- Anastassiou, C.A.; Koch, C. Ephaptic coupling to endogenous electric field activity: Why bother? Curr. Opin. Neurobiol. 2015, 31, 95–103. [Google Scholar] [CrossRef]
- Tang, R.; Dai, J. Biophoton signal transmission and processing in the brain. J. Photochem. Photobiol. B Biol. 2014, 139, 71–75. [Google Scholar] [CrossRef]
- Adams, D.L.; Horton, J.C. Capricious expression of cortical columns in the primate brain. Nat. Neurosci. 2003, 6, 113–114. [Google Scholar] [CrossRef]
- Rakic, P. Evolution of the neocortex: A perspective from developmental biology. Nat. Rev. Neurosci. 2009, 10, 724–735. [Google Scholar] [CrossRef]
- Molnár, Z.; Rockland, K.S. Cortical columns. In Neural Circuit and Cognitive Development; Academic Press: Cambridge, MA, USA, 2020; pp. 103–126. [Google Scholar]
- Buxhoeveden, D.P.; Casanova, M.F. The minicolumn hypothesis in neuroscience. Brain 2002, 125, 935–951. [Google Scholar] [CrossRef]
- Buxhoeveden, D.P.; Casanova, M.F. The minicolumn and evolution of the brain. Brain Behav. Evol. 2002, 60, 125–151. [Google Scholar] [CrossRef]
- Casanova, M.F.; Buxhoeveden, D.P.; Switala, A.E.; Roy, E. Minicolumnar pathology in autism. Neurology 2002, 58, 428–432. [Google Scholar] [CrossRef]
- Hutsler, J.J.; Casanova, M.F. Cortical construction in autism spectrum disorder: Columns, connectivity and the subplate. Neuropathol. Appl. Neurobiol. 2016, 42, 115–134. [Google Scholar] [CrossRef]
- Beker, S.; Kellner, V.; Kerti, L.; Stern, E.A. Interaction between amyloid-β pathology and cortical functional columnar organization. J. Neurosci. 2012, 32, 11241–11249. [Google Scholar] [CrossRef]
- Casanova, M.F.; Buxhoeveden, D.P.; Cohen, M.; Switala, A.E.; Roy, E.L. Minicolumnar pathology in dyslexia. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 2002, 52, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Aceves, G.A.G.; López, M.A.C.; Vanegas, M.A.; Meléndez, O.R.M.; Jiménez, S.M.; Cruz, J.C.P.; Peregrino, R.D.; Aguilar, A.G.; González, J.A.H. Fractal anatomy of the hippocampal formation. Surg. Radiol. Anat. 2018, 40, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, H.F.; Elston, G.N. Dendritic branching of pyramidal cells in the visual cortex of the nocturnal owl monkey: A fractal analysis. Fractals 2003, 11, 391–396. [Google Scholar] [CrossRef]
- Kiselev, V.G.; Hahn, K.R.; Auer, D.P. Is the brain cortex a fractal? Neuroimage 2003, 20, 1765–1774. [Google Scholar] [CrossRef]
- Milošević, N.T.; Ristanović, D.; Stanković, J.B. Fractal analysis of the laminar organization of spinal cord neurons. J. Neurosci. Methods 2005, 146, 198–204. [Google Scholar] [CrossRef]
- Mountcastle, V.B. Modality and topographic properties of single neurons of cat’s somatic sensory cortex. J. Neurophysiol. 1957, 20, 408–434. [Google Scholar] [CrossRef]
- Bugbee, N.M.; Goldman-Rakic, P.S. Columnar organization of corticocortical projections in squirrel and rhesus monkeys: Similarity of column width in species differing in cortical volume. J. Comp. Neurol. 1983, 220, 355–364. [Google Scholar] [CrossRef]
- Koo, B.-B.; Schettler, S.P.; Murray, D.E.; Lee, J.-M.; Killiany, R.J.; Rosene, D.L.; Kim, D.-S.; Ronen, I. Age-related effects on cortical thickness patterns of the Rhesus monkey brain. Neurobiol. Aging 2012, 33, 200.e23–200.e31. [Google Scholar] [CrossRef]
- Cowey, A. Projection of the retina on to striate and prestriate cortex in the squirrel monkey, Saimiri sciureus. J. Neurophysiol. 1964, 27, 366–393. [Google Scholar] [CrossRef]
- Petersen, C.C.; Sakmann, B. Functionally independent columns of rat somatosensory barrel cortex revealed with voltage-sensitive dye imaging. J. Neurosci. 2001, 21, 8435–8446. [Google Scholar] [CrossRef]
- Bureau, I.; Shepherd, G.M.; Svoboda, K. Precise development of functional and anatomical columns in the neocortex. Neuron 2004, 42, 789–801. [Google Scholar] [CrossRef]
- Trinh, A.T.; Harvey-Girard, E.; Teixeira, F.; Maler, L. Cryptic laminar and columnar organization in the dorsolateral pallium of a weakly electric fish. J. Comp. Neurol. 2016, 524, 408–428. [Google Scholar] [CrossRef]
- Wang, Y.; Brzozowska-Prechtl, A.; Karten, H.J. Laminar and columnar auditory cortex in avian brain. Proc. Natl. Acad. Sci. USA 2010, 107, 12676–12681. [Google Scholar] [CrossRef]
- LaBerge, D. Attention, consciousness, and electrical wave activity within the cortical column. Int. J. Psychophysiol. 2001, 43, 5–24. [Google Scholar] [CrossRef]
- Pakkenberg, B.; Gundersen, H.J.G. Neocortical neuron number in humans: Effect of sex and age. J. Comp. Neurol. 1997, 384, 312–320. [Google Scholar] [CrossRef]
- Goldman-Rakic, P.S.; Selemon, L.D.; Schwartz, M.L. Dual pathways connecting the dorsolateral prefrontal cortex with the hippocampal formation and parahippocampal cortex in the rhesus monkey. Neuroscience 1984, 12, 719–743. [Google Scholar] [CrossRef]
- Burggren, A.C.; Siddarth, P.; Mahmood, Z.; London, E.D.; Harrison, T.M.; Merrill, D.A.; Small, G.W.; Bookheimer, S.Y. Subregional hippocampal thickness abnormalities in older adults with a history of heavy cannabis use. Cannabis Cannabinoid Res. 2018, 3, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Buxhoeveden, D.P. Minicolumn size and human cortex. Prog. Brain Res. 2012, 195, 219–235. [Google Scholar]
- Casanova, M.F.; Buxhoeveden, D.; Gomez, J. Disruption in the inhibitory architecture of the cell minicolumn: Implications for autisim. Neuroscientist 2003, 9, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Milošević, N.T.; Ristanović, D. Fractality of dendritic arborization of spinal cord neurons. Neurosci. Lett. 2006, 396, 172–176. [Google Scholar] [CrossRef]
- Tallinen, T.; Chung, J.Y.; Rousseau, F.; Girard, N.; Lefèvre, J.; Mahadevan, L. On the growth and form of cortical convolutions. Nat. Phys. 2016, 12, 588–593. [Google Scholar] [CrossRef]
- Schwartz, E.L. Cortical anatomy, size invariance, and spatial frequency analysis. Perception 1981, 10, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Song, J.; Park, G.; Kim, J. Modeling of autism using organoid technology. Mol. Neurobiol. 2017, 54, 7789–7795. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, M.; Svenningsen, Å.F.; Thorsen, M.; Michel, T.M. Psychiatry in a dish: Stem cells and brain organoids modeling autism spectrum disorders. Biol. Psychiatry 2018, 83, 558–568. [Google Scholar] [CrossRef]
- Cairns, D.M.; Rouleau, N.; Parker, R.N.; Walsh, K.G.; Gehrke, L.; Kaplan, D.L. A 3D human brain–like tissue model of herpes-induced Alzheimer’s disease. Sci. Adv. 2020, 6, eaay8828. [Google Scholar] [CrossRef]
- Lee, P.; Kim, H.R.; Jeong, Y. Detection of gray matter microstructural changes in Alzheimer’s disease continuum using fiber orientation. BMC Neurol. 2020, 20, 362. [Google Scholar] [CrossRef]
- de Lussanet, M.H.; Boström, K.J.; Wagner, H. A model of uniform cortical composition predicts the scaling laws of the mammalian cerebrum. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chwalek, K.; Tang-Schomer, M.D.; Omenetto, F.G.; Kaplan, D.L. In vitro bioengineered model of cortical brain tissue. Nat. Protoc. 2015, 10, 1362–1373. [Google Scholar] [CrossRef]
- Knowlton, S.; Anand, S.; Shah, T.; Tasoglu, S. Bioprinting for neural tissue engineering. Trends Neurosci. 2018, 41, 31–46. [Google Scholar] [CrossRef]
- Rouleau, N.; Murugan, N.J.; Kaplan, D.L. Functional bioengineered models of the central nervous system. Nat. Rev. Bioeng. 2023, 1, 252–270. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Column Dimensions (µm) | Predicted Wavenumbers | References | |||
|---|---|---|---|---|---|---|
| Width | Height | |||||
| Min | Max | Min | Max | |||
| Rhesus monkey (M. mulatta) | 380 | 1190 | 1990 | 2750 | 1.67–7.24 | [35,36] |
| Squirrel monkey (S. sciureus) | 330 | 830 | 1176 | 1664 | 1.42–5.04 | [35] |
| Wistar rat (M. musculus) | 170 | 435 | 735 | 900 | 1.69–5.29 | [38,39] |
| Electric fish (A. leptorhynchus) | 41.6 | 166.4 | 300 | 300 | 1.80–7.21 | [40] |
| Chick (G. gallus) | 300 | 500 | 640 | 1928 | 1.28–6.43 | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rouleau, N.; Murugan, N.J. Self-Similarity and Spatial Periodicity in Cerebral Cortical Patterning: Structural Design Notes for Neural Tissue Architects. Anatomia 2023, 2, 222-231. https://doi.org/10.3390/anatomia2030020
Rouleau N, Murugan NJ. Self-Similarity and Spatial Periodicity in Cerebral Cortical Patterning: Structural Design Notes for Neural Tissue Architects. Anatomia. 2023; 2(3):222-231. https://doi.org/10.3390/anatomia2030020
Chicago/Turabian StyleRouleau, Nicolas, and Nirosha J. Murugan. 2023. "Self-Similarity and Spatial Periodicity in Cerebral Cortical Patterning: Structural Design Notes for Neural Tissue Architects" Anatomia 2, no. 3: 222-231. https://doi.org/10.3390/anatomia2030020
APA StyleRouleau, N., & Murugan, N. J. (2023). Self-Similarity and Spatial Periodicity in Cerebral Cortical Patterning: Structural Design Notes for Neural Tissue Architects. Anatomia, 2(3), 222-231. https://doi.org/10.3390/anatomia2030020