
Infectivity of Viable but Non-Cultivable Cells of Pigmented and Nonpigmented Xanthomonas citri pv. anacardii Strains Demonstrate the Need to Establish Indexing Protocols for Cashew Propagules

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
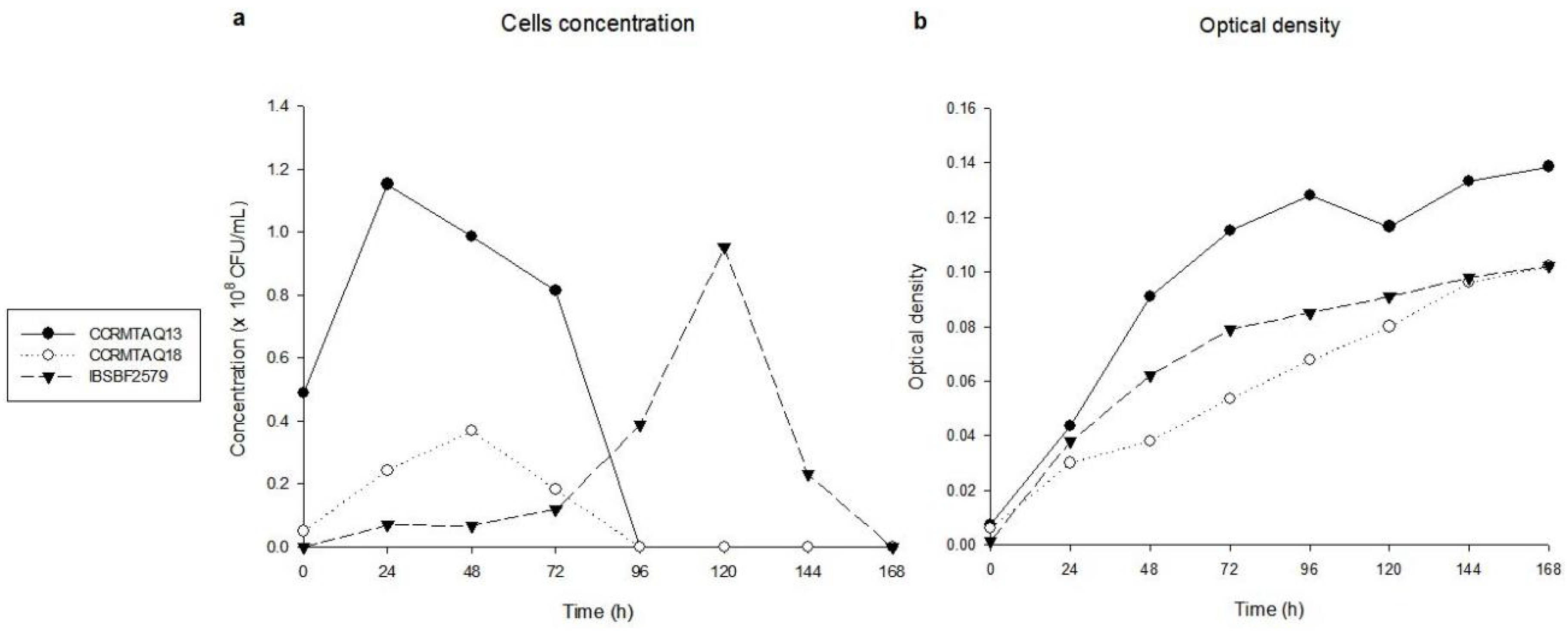
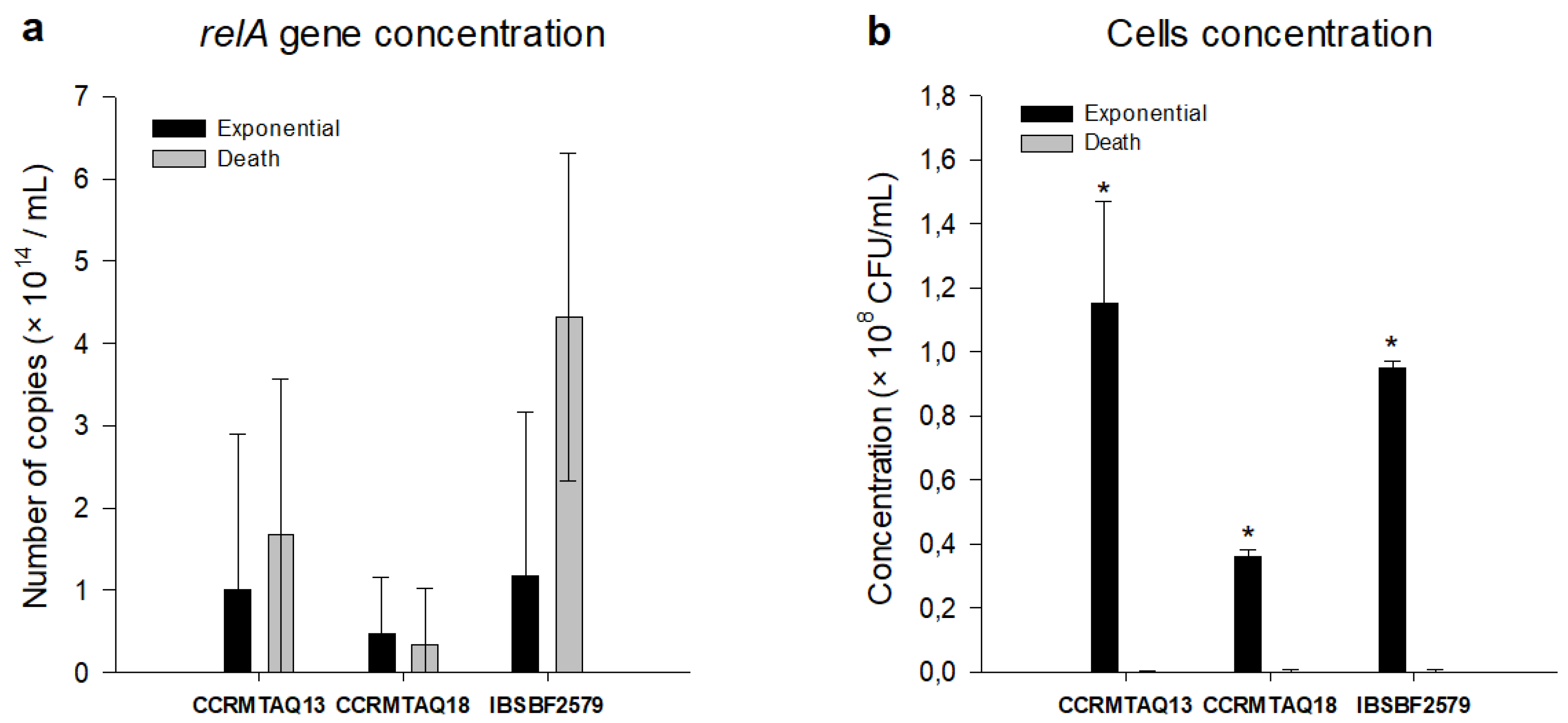
2.1. In Vitro Growth Phases
2.2. Genomic Annotation, Comparative Analysis, and Primer Construction for VBNC Status Assessment
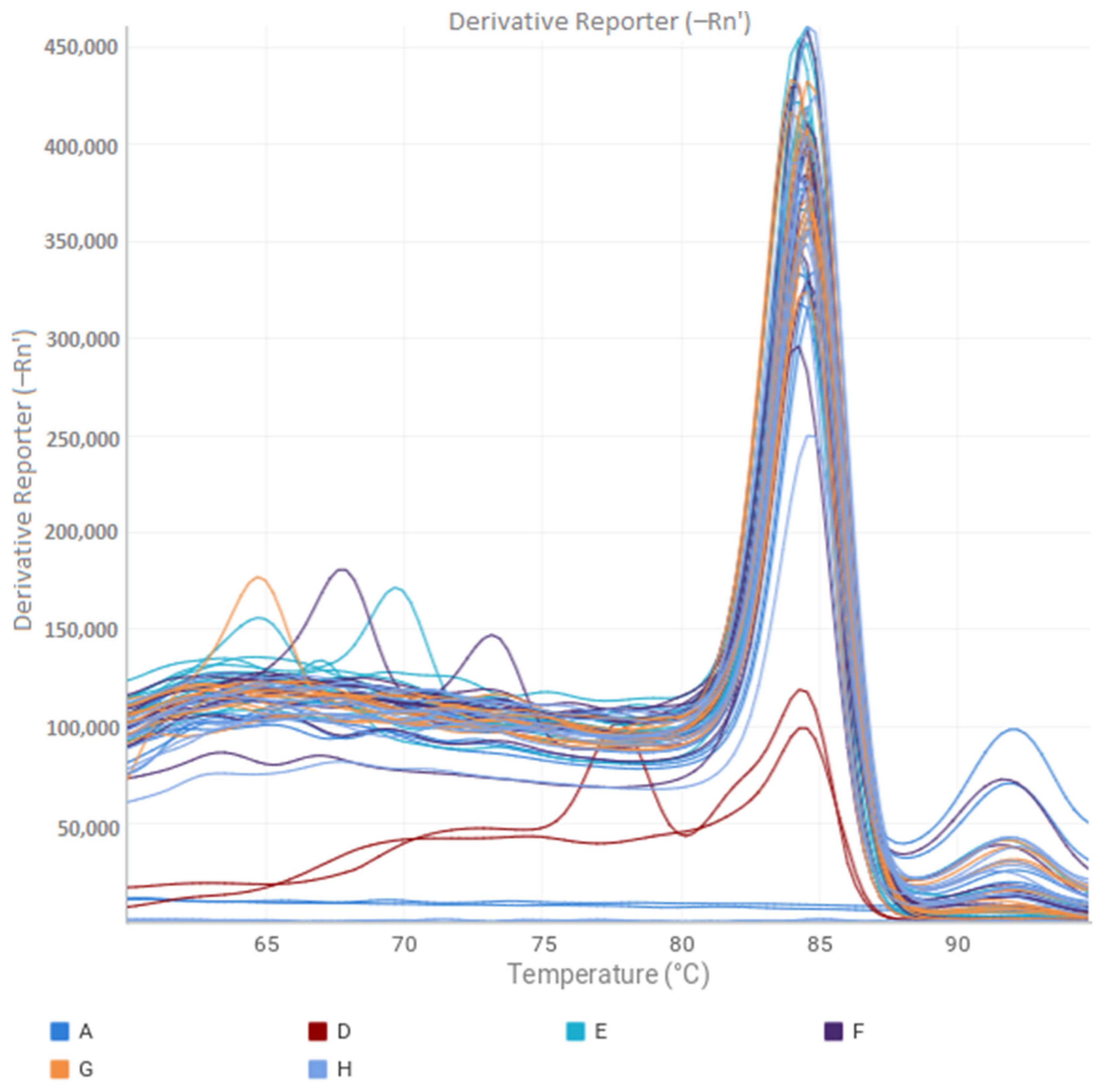
2.3. VBNC State Evaluation
2.4. VBNC State Cell Infectivity
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
3.2. Determination of Exponential and Death Phases In Vitro
3.3. Genomic Annotation and Design of Primers for Evaluation of VBNC State
3.4. Assessment of VBNC Status via Quantitative PCR (qPCR)
3.5. VBNC Cell Infectivity in Cashew Seedlings
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barros, L.M. Botânica, origem e distribuição geográfica. In Cajucultura: Modernas Técnicas de Produção; Araújo, J.P.P., Silva, V.V., Eds.; EMBRAPA-CNPAT: Fortaleza, Brazil, 1995; pp. 55–71. [Google Scholar]
- Melo, Q.M.S.; Bleicher, E. Identification and management of the main cashew pests. In Cashew Plant Health; Melo, Q.M.S., Ed.; Embrapa Agroindústria Tropical: Brasília, Brazil, 2002; pp. 9–34. [Google Scholar]
- IBGE. Table 4—Area Destined for Harvest, Harvested Area, Quantity Produced, Average Yield and Value of Production in Brazil, the Major Regions and Federation Units, according to the Products of Permanent Crops; Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brazil, 2019. Available online: https://www.ibge.gov.br/estatisticas/economicas/agricultura-e-pecuaria/9117-producao-agricola-municipal-culturas-temporarias-e-permanentes.html?edicao=29008&t=resultados (accessed on 20 May 2022).
- Gama Marco, A.S.; Mariano, R.L.R.; Viana, F.M.P.; Ferreira, M.A.S.V.; Souza, E.B. Polyphasic Characterization of Pigmented Strains of Xanthomonas Pathogenic to Cashew Trees. Plant Dis. 2011, 95, 793–802. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Viana, F.M.P.; Cardoso, J.E.; Saraiva, H.A.O.; Ferreira, M.A.S.V.; Mariano, R.L.R.; Trindade, L.C. First Report of a Bacterial Leaf and Fruit Spot of Cashew Nut (Anacardium occidentale) Caused by Xanthomonas campestris pv. mangiferaeindicae in Brazil. Plant Dis. 2007, 91, 1361. [Google Scholar] [CrossRef] [PubMed]
- Da Gama, M.A.S.; Mariano, R.D.L.R.; Da Silva Júnior, W.J.; De Farias, A.R.G.; Barbosa, M.A.G.; Ferreira, M.Á.D.S.V.; Costa Júnior, C.R.L.; Santos, L.A.; De Souza, E.B. Taxonomic Repositioning of Xanthomonas campestris pv. viticola (Nayudu 1972) Dye 1978 as Xanthomonas citri pv. viticola (Nayudu 1972) Dye 1978 comb. nov. and Emendation of the Description of Xanthomonas citri pv. anacardii to Include Pigmented Isolates Pathogenic to Cashew Plant. Phytopathology 2018, 108, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information (NCBI). NCBI Genome. 2021. Available online: https://www.ncbi.nlm.nih.gov/genome/genomes/527? (accessed on 30 June 2021).
- Ghezzi, J.I.; Steck, T.R. Induction of the viable but non-culturable condition in Xanthomonas campestris pv. campestris in liquid microcosms and sterile soil. FEMS Microbiol. Ecol. 1999, 30, 203–208. [Google Scholar] [CrossRef]
- Campo, R.D.; Russi, P.; Mara, P.; Mara, H.; Peyrou, M.; León, I.P.D.; Gaggero, C. Xanthomonas axonopodis pv. citri enters the VBNC state after copper treatment and retains its virulence. FEMS Microbiol. Lett. 2009, 298, 143–148. [Google Scholar] [CrossRef]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef]
- Su, X.; Guo, L.; Ding, L.; Qu, K.; Shen, C. Induction of Viable but Nonculturable State in Rhodococcus and Transcriptome Analysis Using RNA-seq. PLoS ONE 2016, 11, e0147593. [Google Scholar] [CrossRef]
- Golmohammadi, M.; Cubero, J.; López, M.M.; Llop, P. The viable but non-culturable state in Xanthomonas citri subsp. citri is a reversible state induced by low nutrient availability and copper stress conditions. J. Life Sci. 2013, 7, 1019–1029. [Google Scholar]
- Lee, J.H.; Zhao, Y. Integration Host Factor Is Required for RpoN-Dependent hrpL Gene Expression and Controls Motility by Positively Regulating rsmB sRNA in Erwinia Amylovora. Phytopathology 2015, 106, 29–36. [Google Scholar] [CrossRef]
- Santander, R.D.; Figàs-Segura, À.; Biosca, E.G. Erwinia amylovora catalases KatA and KatG are virulence factors and delay the starvation-induced viable but non-culturable (VBNC) response. Mol. Plant Pathol. 2018, 19, 922–934. [Google Scholar] [CrossRef]
- Jiang, N.; Lv, Q.Y.; Xu, X.; Cao, Y.S.; Walcott, R.R.; Li, J.Q.; Luo, L.X. Induction of the viable but nonculturable state in Clavibacter michiganensis subsp. michiganensis and in planta resuscitation of the cells on tomato seedlings. Plant Pathol. 2016, 65, 826–836. [Google Scholar] [CrossRef]
- Postnikova, O.A.; Shao, J.; Mock, N.M.; Baker, C.J.; Nemchinov, L.G. Gene Expression Profiling in Viable but Nonculturable (VBNC) Cells of Pseudomonas syringae pv. syringae. Front. Microbiol. 2015, 6, 1419. [Google Scholar] [CrossRef]
- Kan, Y.; Jiang, N.; Xu, X.; Lyu, Q.; Gopalakrishnan, V.; Walcott, R.; Burdman, S.; Li, J.; Luo, L. Induction and Resuscitation of the Viable but Non-culturable (VBNC) State in Acidovorax citrulli, the Causal Agent of Bacterial Fruit Blotch of Cucurbitaceous Crops. Front. Microbiol. 2019, 10, 1081. [Google Scholar] [CrossRef]
- Kong, H.G.; Bae, J.Y.; Lee, H.J.; Joo, H.J.; Jung, E.J.; Chung, E.; Lee, S.-W. Induction of the Viable but Nonculturable State of Ralstonia solanacearum by Low Temperature in the Soil Microcosm and Its Resuscitation by Catalase. PLoS ONE 2014, 9, e109792. [Google Scholar] [CrossRef]
- Palmieri, A.C.; Amaral, A.M.; Homem, R.A.; Machado, M.A. Differential expression of pathogenicity- and virulence-related genes of Xanthomonas axonopodis pv. citri under copper stress. Genet. Mol. Biol. 2010, 33, 348–353. [Google Scholar] [CrossRef]
- Asakura, H.; Ishiwa, A.; Arakawa, E.; Makino, S.; Okada, Y.; Yamamoto, S.; Igimi, S. Gene expression profile of Vibrio cholerae in the cold stress-induced viable but non-culturable state. Environ. Microbiol. 2007, 9, 869–879. [Google Scholar] [CrossRef]
- Um, H.Y.; Kong, H.G.; Lee, H.J.; Choi, H.K.; Park, E.J.; Kim, S.T.; Murugiyan, S.; Chung, E.; Kang, K.Y.; Lee, S. Altered Gene Expression and Intracellular Changes of the Viable But Nonculturable State in Ralstonia solanacearum by Copper Treatment. Plant Pathol. J. 2013, 29, 374–385. [Google Scholar] [CrossRef]
- Meng, L.; Alter, T.; Aho, T.; Huehn, S. Gene expression profiles of Vibrio parahaemolyticus in viable but non-culturable state. FEMS Microbiol. Ecol. 2015, 91, fiv035. [Google Scholar] [CrossRef]
- Ordax, M.; Biosca, E.G.; Wimalajeewa, S.C.; López, M.M.; Marco-Noales, E. Survival of Erwinia amylovora in mature apple fruit calyces through the viable but nonculturable (VBNC) state. J. Appl. Microbiol. 2009, 107, 106–116. [Google Scholar] [CrossRef]
- Yaron, S.; Matthews, K.R. A reverse transcriptase-polymerase chain reaction assay for detection of viable Escherichia coli O157:H7: Investigation of specific target genes. J. Appl. Microbiol. 2002, 92, 633–640. [Google Scholar] [CrossRef]
- Chen, S.; Li, X.; Wang, Y.; Zeng, J.; Ye, C.; Li, X.; Guo, L.; Zhang, S.; Yu, X. Induction of Escherichia coli into a VBNC state through chlorination/chloramination and differences in characteristics of the bacterium between states. Water Res. 2018, 142, 279–288. [Google Scholar] [CrossRef]
- Jameelah, M.; Dewanti-Hariyadi, R.; Nurjanah, S. Expression of rpoS, ompA and hfq genes of Cronobacter sakazakii strain Yrt2a during stress and viable but nonculturable state. Food Sci. Biotechnol. 2018, 27, 915–920. [Google Scholar] [CrossRef]
- Silva Junior, W.J.; Farias, A.R.; Lima, N.B.; Benko-Iseppon, A.M.; Aburjaile, F.; Balbino, V.Q.; Falcão, R.M.; Leitão Paiva Júnior, S.D.; Sousa-Paula, L.C.; Mariano, R.L.; et al. Complete Genome Sequence of Xanthomonas citri pv. anacardii Strain IBSBF2579 from Brazil. Genome Announc. 2018, 6, e01574-17. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bairoch, A.; Apweiler, R.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gaisteger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. The Universal Protein Resource (UniProt). Nucleic Acids Res. 2005, 33 (Suppl. S1), D154–D159. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Aburjaile, F.F.; Madec, M.-N.; Parayre, S.; Miyoshi, A.; Azevedo, V.; Loir, Y.L.; Falentin, H. The long-term survival of Propionibacterium freudenreichii in a context of nutrient shortage. J. Appl. Microbiol. 2016, 120, 432–440. [Google Scholar] [CrossRef]
- Maximiano, M.R.; Oliveira-Neto, O.B.; Franco, O.L.; Mehta, A. Validation of an in vitro system for studies of pathogenicity mechanisms in Xanthomonas campestris. FEMS Microbiol. Lett. 2017, 364, fnx217. [Google Scholar] [CrossRef]
- González-Escalona, N.; Fey, A.; Höfle, M.G.; Espejo, R.T.; Guzmán, C.A. Quantitative reverse transcription polymerase chain reaction analysis of Vibrio cholerae cells entering the viable but non-culturable state and starvation in response to cold shock. Environ. Microbiol. 2006, 8, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Avarbock, D.; Salem, J.; Li, L.; Wang, Z.; Rubin, H. Cloning and characterization of a bifunctional RelA/SpoT homologue from Mycobacterium Tuberculosis. Gene 1999, 233, 261–269. [Google Scholar] [CrossRef]
- Potrykus, K.; Cashel, M. Preferential Cellular Accumulation of ppGpp or pppGpp in Escherichia coli. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; De Brujin, F.J., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 479–488. [Google Scholar] [CrossRef]
- Primm, T.P.; Andersen, S.J.; Mizrahi, V.; Avarbock, D.; Rubin, H.; Barry, C.E. The Stringent Response of Mycobacterium tuberculosis Is Required for Long-Term Survival. J. Bacteriol. 2000, 182, 4889–4898. [Google Scholar] [CrossRef] [PubMed]
- Catara, V.; Cubero, J.; Pothier, J.F.; Bosis, E.; Bragard, C.; Đermić, E.; Holeva, M.C.; Jacques, M.; Petter, F.; Pruvost, O.; et al. Trends in Molecular Diagnosis and Diversity Studies for Phytosanitary Regulated Xanthomonas. Microorganisms 2021, 9, 862. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Product | Gene Size (bp) | Reference |
|---|---|---|---|
| fliG | FliG flagellar protein | 975 | [20] |
| relA | (P)ppGpp synthase | 2163 | |
| omp | Outer membrane protein | 1515 | [21] |
| rps | 30S Ribossomal protein | 789 | |
| dps | DNA-binding ferritin-like protein (Oxidative damage protectant) | 540 | |
| mobA | Molybdenum cofactor guanylyltransferase | 570 | [24] |
| soxR | Transcriptional regulator soxR family | 451 | [25] |
| katG | Catalase-peroxidase | 2249 | |
| gltB | Glutamate synthase alfa subunit | 4470 | [22] |
| gltD | Glutamate synthase beta subunit | 1410 | |
| pilM | Type IV pilus assembly protein | 1086 | |
| hfq | RNA Hfq binding protein | 282 | [26] |
| murG | UDP-N-acetylglucosamine–N-acetylmuramyl-(pentapeptide) pyrophosphoryl-undecaprenol N-acetylglucosamine transferase | 1290 | [16] |
| Incubation Period (Days) | Final Severity (mm2) | |||
|---|---|---|---|---|
| Strain | Exponential | VBNC | Exponential | VBNC |
| CCRMTAQ13 | 2.44 bA 1 | 2.66 bA | 6.91 aA | 5.43 aB |
| CCRMTAQ18 | 3.13 aA | 2.69 bA | 4.72 cA | 5.09 aA |
| IBSBF2579 | 2.27 bB | 3.41 aA | 5.98 bA | 5.12 aB |
| VC 2 (%) | 24.9 | 15.71 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucena, L.P.; Albuquerque, G.R.; Souza, E.B.; Benko-Iseppon, A.M.; Carvalho, R.D.O.; Aburjaile, F.F.; Gama, M.A.S. Infectivity of Viable but Non-Cultivable Cells of Pigmented and Nonpigmented Xanthomonas citri pv. anacardii Strains Demonstrate the Need to Establish Indexing Protocols for Cashew Propagules. Bacteria 2022, 1, 207-217. https://doi.org/10.3390/bacteria1040016
Lucena LP, Albuquerque GR, Souza EB, Benko-Iseppon AM, Carvalho RDO, Aburjaile FF, Gama MAS. Infectivity of Viable but Non-Cultivable Cells of Pigmented and Nonpigmented Xanthomonas citri pv. anacardii Strains Demonstrate the Need to Establish Indexing Protocols for Cashew Propagules. Bacteria. 2022; 1(4):207-217. https://doi.org/10.3390/bacteria1040016
Chicago/Turabian StyleLucena, Lucas Pontes, Géssyka Rodrigues Albuquerque, Elineide Barbosa Souza, Ana Maria Benko-Iseppon, Rodrigo Dias Oliveira Carvalho, Flávia Figueira Aburjaile, and Marco Aurélio Siqueira Gama. 2022. "Infectivity of Viable but Non-Cultivable Cells of Pigmented and Nonpigmented Xanthomonas citri pv. anacardii Strains Demonstrate the Need to Establish Indexing Protocols for Cashew Propagules" Bacteria 1, no. 4: 207-217. https://doi.org/10.3390/bacteria1040016
APA StyleLucena, L. P., Albuquerque, G. R., Souza, E. B., Benko-Iseppon, A. M., Carvalho, R. D. O., Aburjaile, F. F., & Gama, M. A. S. (2022). Infectivity of Viable but Non-Cultivable Cells of Pigmented and Nonpigmented Xanthomonas citri pv. anacardii Strains Demonstrate the Need to Establish Indexing Protocols for Cashew Propagules. Bacteria, 1(4), 207-217. https://doi.org/10.3390/bacteria1040016