
Anatomy and Germination of Erythrina velutina Seeds under a Different Imbibition Period in Gibberellin

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
- % of germination (G) or germinability, calculated through [22] formula:in which: N = the number of germinated seeds by the end of the test; A = the total number of seeds placed to germinate.G = (N/A) × 100
- Germination speed index (GSI), calculated by the formula of [23]:in which: ESI = the germination speed index; G1, G2, Gn = the number of seeds germinated in the first, second and last counting; N1, N2, Nn = the number of sowing days at first, second and last counting.GSI = G1/N1 + G2/N2 + ⋯ + Gn/Nn
- Mean germination time (t) in days, calculated by the formula [24]:in which: ni = the number of seeds germinated by day; ti = the time of incubation.t = (∑niti)/∑ni
- Average germination speed (AGS) in days, calculated by the formula [24]:in which: t = the medium germination time.AGS = 1/t
- Relative germination frequency, in percentage, according to the formula [22]:in which: Fr = the germination relative frequency; ni = the number of seeds germinated by day; ∑ni = the total amount of germinated seeds.
3. Results
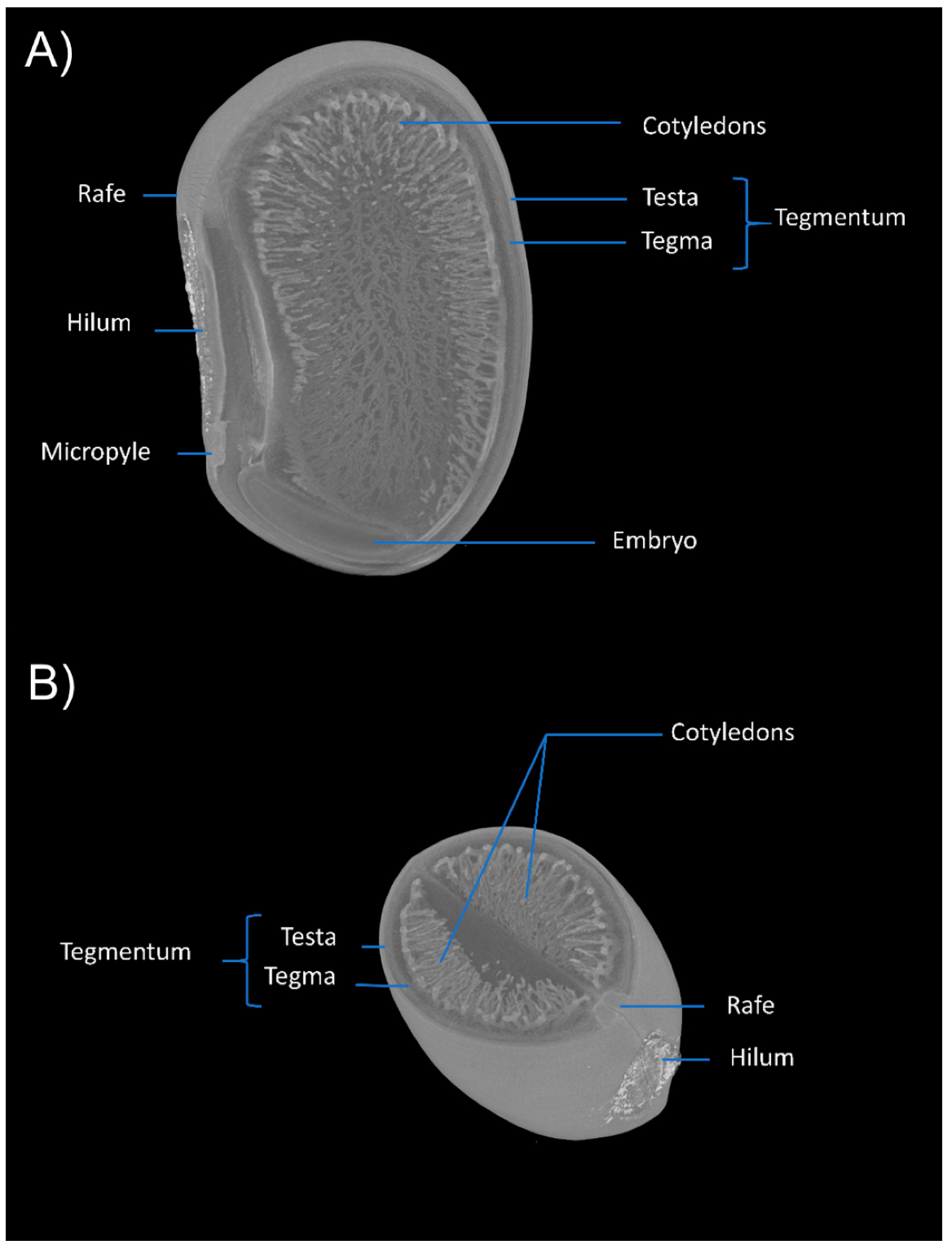
3.1. Anatomical Aspects
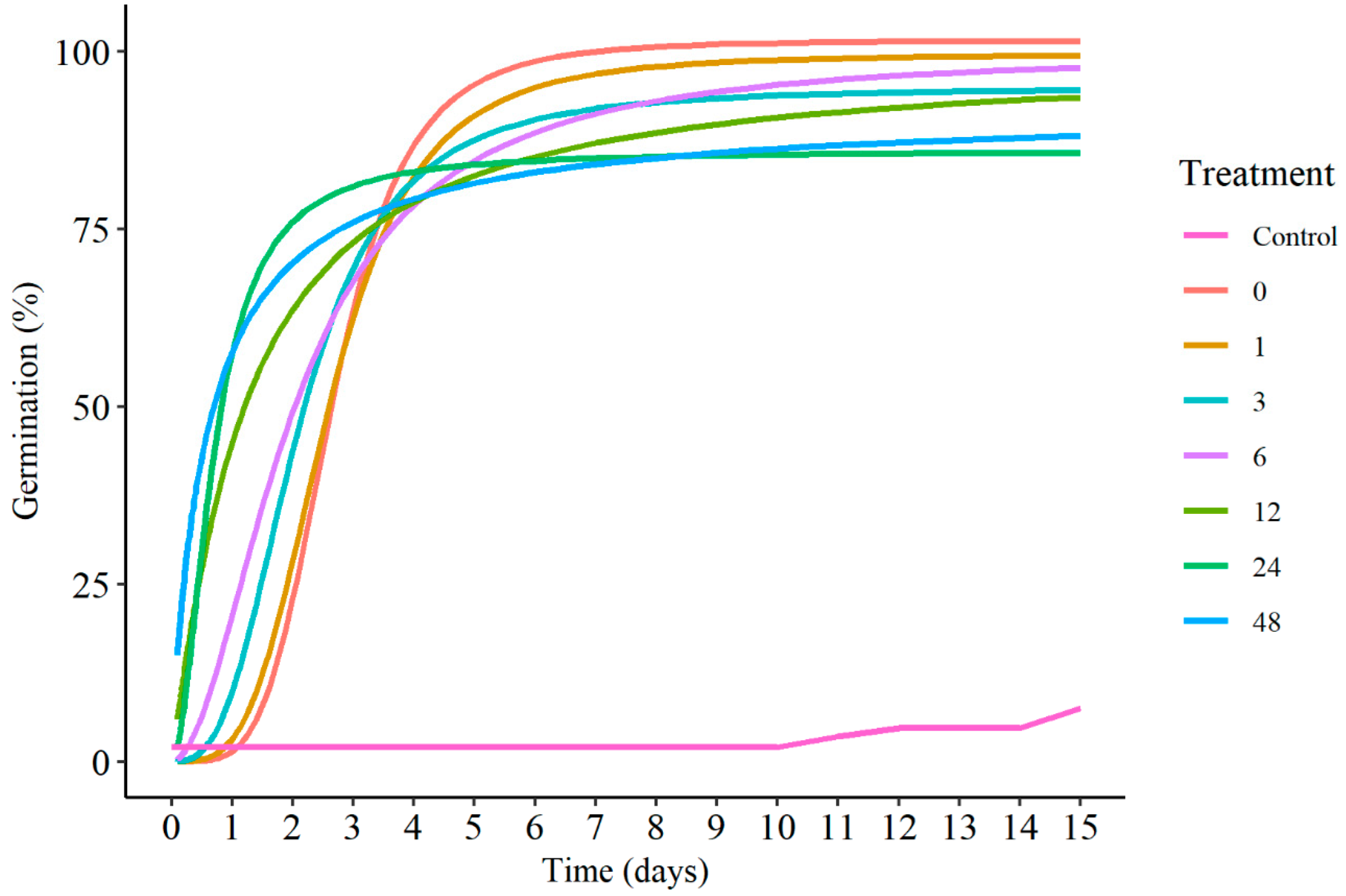
3.2. Germination Aspects
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Queiroz, L.P.; Cardoso, D.; Fernandes, M.F.; Moro, M.F. Diversity and Evolution of Flowering Plants of the Caatinga Domain. In Caatinga; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Junior, C.R.P.; Salvador, C.A.; Tavares, T.R.; Abreu, M.C.; Fagundes, H.S.; Almeida, W.S.; Neto, E.C.S.; Anjos, L.H.C.; Pereira, M.G. Lithic soils in the semi-arid region of Brazil: Edaphic characterization and susceptibility to erosion. J. Arid. Land 2022, 14, 56–69. [Google Scholar] [CrossRef]
- Salazar, A.A.; Arellano, E.C.; Muñoz-Sáez, A.; Miranda, M.D.; da Silva, F.O.; Zielonka, N.B.; Crowther, L.P.; Silva-Ferreira, V.; Oliveira-Reboucas, P.; Dicks, L.V. Restoration and conservation of priority areas of Caatinga’s semi-arid forest remnants can support connectivity within an agricultural landscape. Land 2021, 10, 550. [Google Scholar] [CrossRef]
- Ribeiro, E.M.S.; Arroyo-Rodríguez, V.; Santos, B.; Tabarelli, M.; Leal, I.R. Chronic anthropogenic disturbance drives the biological impoverishment of the Brazilian Caatinga vegetation. J. Appl. Ecol. 2015, 52, 611–620. [Google Scholar] [CrossRef]
- Schulz, K.; Voigt, K.; Beusch, C.; Almeida-Cortez, J.S.; Kowarik, I.; Walz, A.; Cierjacks, A. Grazing deteriorates the soil carbon stocks of Caatinga forest ecosystems in Brazil. For. Ecol. Manag. 2016, 367, 62–70. [Google Scholar] [CrossRef]
- Antongiovanni, M.; Venticinque, E.M.; Fonseca, C.R. Fragmentation patterns of the Caatinga drylands. Landsc. Ecol. 2018, 33, 1353–1367. [Google Scholar] [CrossRef]
- Carvalho, P.E.M. Mulungu (Erythrina velutina); Circular Técnica 160; Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA, Embrapa Florestas: Colombo, PR, Brazil, 2008; pp. 1–8. [Google Scholar]
- Brasileiro, A.C.; Lima-Araujo, F.; Alcântara, J.A.; Pontes, A.S.M.; Neto, J.A.; de Oliveira Tavares, R.; Silvino, A.S.; Vizentin-Bugoni, J. Birds of the Parque Ecológico Lagoa da Fazenda, Sobral, Ceará state, northeastern Brazil. Check List 2017, 13, 2037. [Google Scholar] [CrossRef][Green Version]
- Rodrigues, D.R.; da Silva, A.F.; Cavalcanti, M.I.P.; Escobar, I.E.C.; Fraiz, A.C.R.; Ribeiro, P.R.D.A.; Neto, R.A.F.; de Freitas, A.D.S.; Fernandes-Júnior, P.I. Phenotypic, genetic and symbiotic characterization of Erythrina velutina rhizobia from Caatinga dry forest. Braz. J. Microbiol. 2018, 49, 503–512. [Google Scholar] [CrossRef]
- Silva, P.A.; Silva, L.L.; Brito, L. Using bird-flower interactions to select native tree resources for urban afforestation: The case of Erythrina velutina. Urban For. Urban Green. 2020, 51, 126677. [Google Scholar] [CrossRef]
- da Silva, M.M.; Santana, A.S.; Pimentel, R.M.; Silva, F.C.; Randau, K.P.; Soares, L.A. Anatomy of leaf and stem of Erythrina velutina. Rev. Bras. Farm. 2013, 23, 200–206. [Google Scholar] [CrossRef]
- Oliveira, A.K.; Coelho, M.F.B.; Maia, S.S.S.; Diógenes, F.E.P.; Medeiros Filho, S. Allelopathy of extracts of different organs of coral tree on the germination of lettuce. Hortic. Bras. 2012, 30, 480–483. [Google Scholar] [CrossRef][Green Version]
- Santos, L.W.; Coelho, M.F.B.; Azevedo, R.A.B.; Lima, A.K.B.; Souza, J.W.N. Erythrina velutina Willd: Fabaceae: Árvore de múltiplos usos no nordeste brasileiro. Rev. Verde Agroecol. E Desenvolv. Sustentável 2013, 8, 72–80. [Google Scholar]
- Agra, M.F.; Queiroz, R.T. Erythrina velutina Mulungu. In Espécies Nativas da Flora Brasileira de Valor Econômico Atual ou Potencial: Plantas Para o Futuro: Região Nordeste; Coradin, L., Camillo, J., Pareyn, F.G.C., Eds.; Ministério do Meio Ambiente, Secretaria de Biodiversidade: Brasília, DF, Brazil, 2018; pp. 789–900. [Google Scholar]
- Geisler, G.E.; Pinto, T.T.; Santos, M.; Paulilo, M.T.S. Seed structures in water uptake, dormancy release and germination of two tropical forest Fabaceae species with physically dormant seeds. Braz. J. Bot. 2016, 40, 67–77. [Google Scholar] [CrossRef]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar] [PubMed]
- Gloria, B.A.; Guerreiro, S.M.C. Anatomia Vegetal; Editora UFV: Viçosa, MG, Brazil, 2012. [Google Scholar]
- Almeida, G.M.; Rodrigues, J.G.L. Development of plants by interference auxins, cytokinins, gibberellins and ethylene. Braz. J. Appl. Technol. Agric. Sci. 2016, 9, 111–117. [Google Scholar]
- Urbanova, T.; Leubner-Metzger, G. Gibberellins and seed germination. Annu. Plant Rev. 2016, 49, 253–284. [Google Scholar]
- Filho, J.M. Seed vigor testing: An overview of the past, present and future perspective. Sci. Agric. 2015, 72, 363–374. [Google Scholar] [CrossRef]
- Gomes Junior, F.G.; Cicero, S.M. X-ray analysis to assess mechanical damage in sweet corn seeds. J. Seed Sci. 2012, 34, 78–85. [Google Scholar] [CrossRef]
- Labouriau, L.G.; Valadares, M.E.B. On the germination of seeds Calotropis procera (Ait.) Ait.f. An. Da Acad. Bras. De Ciências 1976, 48, 263–284. [Google Scholar]
- Maguire, J.D. Speeds of germination-aid selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- Ferreira, A.G.; Borghetti, F. Germinação—do Básico ao Aplicado; Artmed: Porto Alegre, RS, Brazil, 2013. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 21 October 2021).
- Matheus, M.T.; Guimarães, R.M.; Bacelar, M.; Oliveira, S.A.D.S. Dormancy overcome of two Erythrina species seeds. Rev. Caatinga 2010, 23, 48–53. [Google Scholar]
- Silva, K.B.; Alves, E.U.; Bruno, R.L.A.; Matos, V.P.; Gonçalves, E.P. Morfologia de frutos, sementes, plântulas e plantas de Erythrina velutina willd, Leguminoseae-Papilionideae. J. Seed Sci. 2008, 30, 104–114. [Google Scholar] [CrossRef]
- Ribeiro, R.C.; Gomes, S.E.V.; Dantas, B.F. Physiological quality of Erythrina velutina Willd. seeds (Fabaceae) under different storage conditions. Sci. For. 2018, 46, 562–570. [Google Scholar] [CrossRef]
- Takata, W.; Silva, E.G.; Corsato, J.M.; Ferreira, G. Germination of pomegranate (Punica granatum L.) seeds under gibberellin concentrations. Rev. Bras. Frutic. 2014, 36, 254–260. [Google Scholar] [CrossRef]
- Cárdenas, J.; Carranza, C.; Miranda, D.; Magnitskiy, S. Effect of GA3, KNO3, and removing of basal point of seeds on germination of sweet granadilla (Passiflora ligularis JUSS) and yellow passion fruit (Passiflora edulis F. flavicarpa). Rev. Bras. Frutic. 2013, 35, 853–859. [Google Scholar] [CrossRef]
- Nobrega, M.A.S.; Pontes, M.S.; Santiago, E.F. Aplicação exógena de GA3 e tiametoxam sobre a dinâmica da germinação de sementes de Psidium guineense Swartz (Myrtaceae). Acta Biomed. Bras. 2018, 9, 58–66. [Google Scholar] [CrossRef]
- De Sousa, H.U.; Ramos, J.D.; Pasqual, M.; Ferreira, E.A. Efeito do ácido giberélico sobre a germinação de sementes de porta-enxertos cítricos. Rev. Bras. Frutic. 2002, 24, 496–499. [Google Scholar] [CrossRef]
- Afonso, M.V.; Paranhos, J.T.; Tabaldi, L.A.; Soriani, H.H. Germinação in vitro de sementes e parâmetros morfofisiológicos de microestacas de Tabernaemontana catharinensis A. DC. Iheringia. Série Botânica 2018, 73, 39–45. [Google Scholar] [CrossRef]
- Junior, C.A.; de Oliveira Vitoriano, J.; Da Silva, D.L.S.; de Lima Farias, M.; de Lima Dantas, N.B. Water uptake mechanism and germination of Erythrina velutina seeds treated with atmospheric plasma. Sci. Rep. 2016, 6, srep33722. [Google Scholar] [CrossRef]
- Siqueira, J.V.G.; Barros, J.P.A.; Araújo, Y.P.; Silva, T.G.F.; Souza, L.S.B. Tratamentos pré-germinativos em sementes de espécies da Caatinga. J. Environ. Anal. Prog. 2017, 2, 499–508. [Google Scholar] [CrossRef]
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil; Editora Plantarum: Nova Odessa, SP, Brazil, 2016. [Google Scholar]
- Silva, K.B.; Alves, E.U.; Bruno, R.L.A.; Pereira, E.; Gonçalves, M.D.S.S.B.; Viana, J.S. Quebra de dormência em sementes de Erythina velutina Willd. Rev. Bras. De Biociências 2007, 5, 180–182. [Google Scholar]
- Rissi, R.N.; Júnior, R.F.G. Escarificação de sementes e quebra de dormência de mulungu (Erythrina velutina WILLD.—Leguminosae). Rev. Biol. FAFIBE 2011, 1, 1–11. [Google Scholar]
- Souza, V.N.; Araújo, A.V.; Pinto, M.A.D.C.; Brito, A.S. Tratamentos físicos e químicos para acelerar e uniformizar a emergência de plântulas de Erythrina velutina Willd. Enciclopédia Biosf. 2016, 13, 1732–1741. [Google Scholar] [CrossRef]
- Phartyal, S.S.; Rosbakh, S.; Ritz, C.; Poschlod, P. Ready for change: Seed traits contribute to the high adaptability of mudflat species to their unpredictable habitat. J. Veg. Sci. 2019, 31, 331–342. [Google Scholar] [CrossRef]
- Rosbakh, S.; Phartyal, S.S.; Poschlod, P. Seed germination traits shape community assembly along a hydroperiod gradient. Ann. Bot. 2019, 125, 67–78. [Google Scholar] [CrossRef]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a Mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guariz, H.R.; Shimizu, G.D.; Paula, J.C.B.; Sperandio, H.V.; Ribeiror Junior, W.A.; Oliveira, H.C.; Jussiani, E.I.; Andrello, A.C.; Marubayashi, R.Y.P.; Picoli, M.H.S.; et al. Anatomy and Germination of Erythrina velutina Seeds under a Different Imbibition Period in Gibberellin. Seeds 2022, 1, 210-220. https://doi.org/10.3390/seeds1030018
Guariz HR, Shimizu GD, Paula JCB, Sperandio HV, Ribeiror Junior WA, Oliveira HC, Jussiani EI, Andrello AC, Marubayashi RYP, Picoli MHS, et al. Anatomy and Germination of Erythrina velutina Seeds under a Different Imbibition Period in Gibberellin. Seeds. 2022; 1(3):210-220. https://doi.org/10.3390/seeds1030018
Chicago/Turabian StyleGuariz, Hugo R., Gabriel D. Shimizu, Jean C. B. Paula, Huezer V. Sperandio, Walte A. Ribeiror Junior, Halley C. Oliveira, Eduardo I. Jussiani, Avacir C. Andrello, Rodrigo Y. P. Marubayashi, Marcelo H. S. Picoli, and et al. 2022. "Anatomy and Germination of Erythrina velutina Seeds under a Different Imbibition Period in Gibberellin" Seeds 1, no. 3: 210-220. https://doi.org/10.3390/seeds1030018
APA StyleGuariz, H. R., Shimizu, G. D., Paula, J. C. B., Sperandio, H. V., Ribeiror Junior, W. A., Oliveira, H. C., Jussiani, E. I., Andrello, A. C., Marubayashi, R. Y. P., Picoli, M. H. S., Ruediger, J., Couto, A. P. S., & Moraes, K. A. M. (2022). Anatomy and Germination of Erythrina velutina Seeds under a Different Imbibition Period in Gibberellin. Seeds, 1(3), 210-220. https://doi.org/10.3390/seeds1030018