
Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Germination Tests
2.3. Data Analyses
3. Results
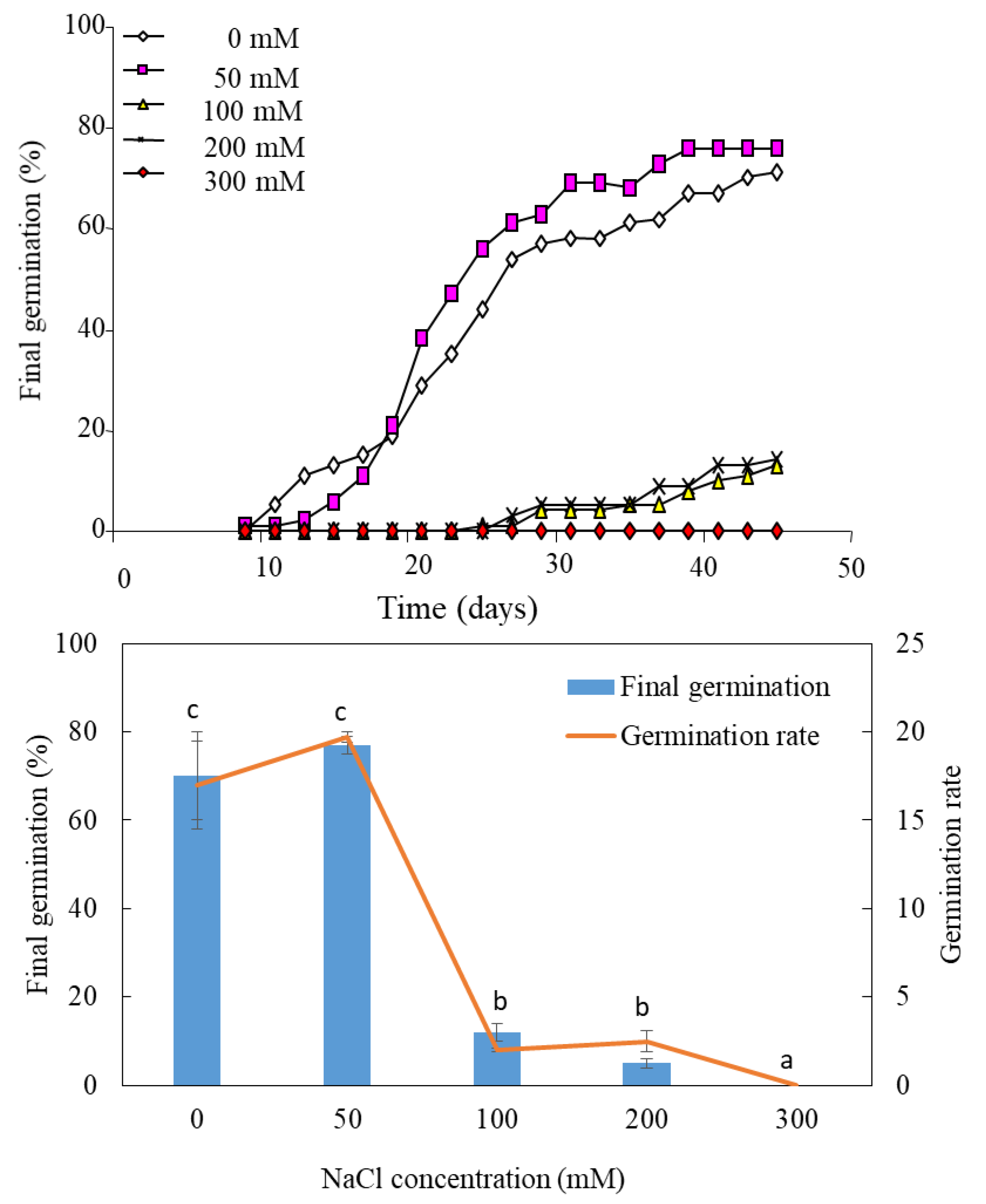
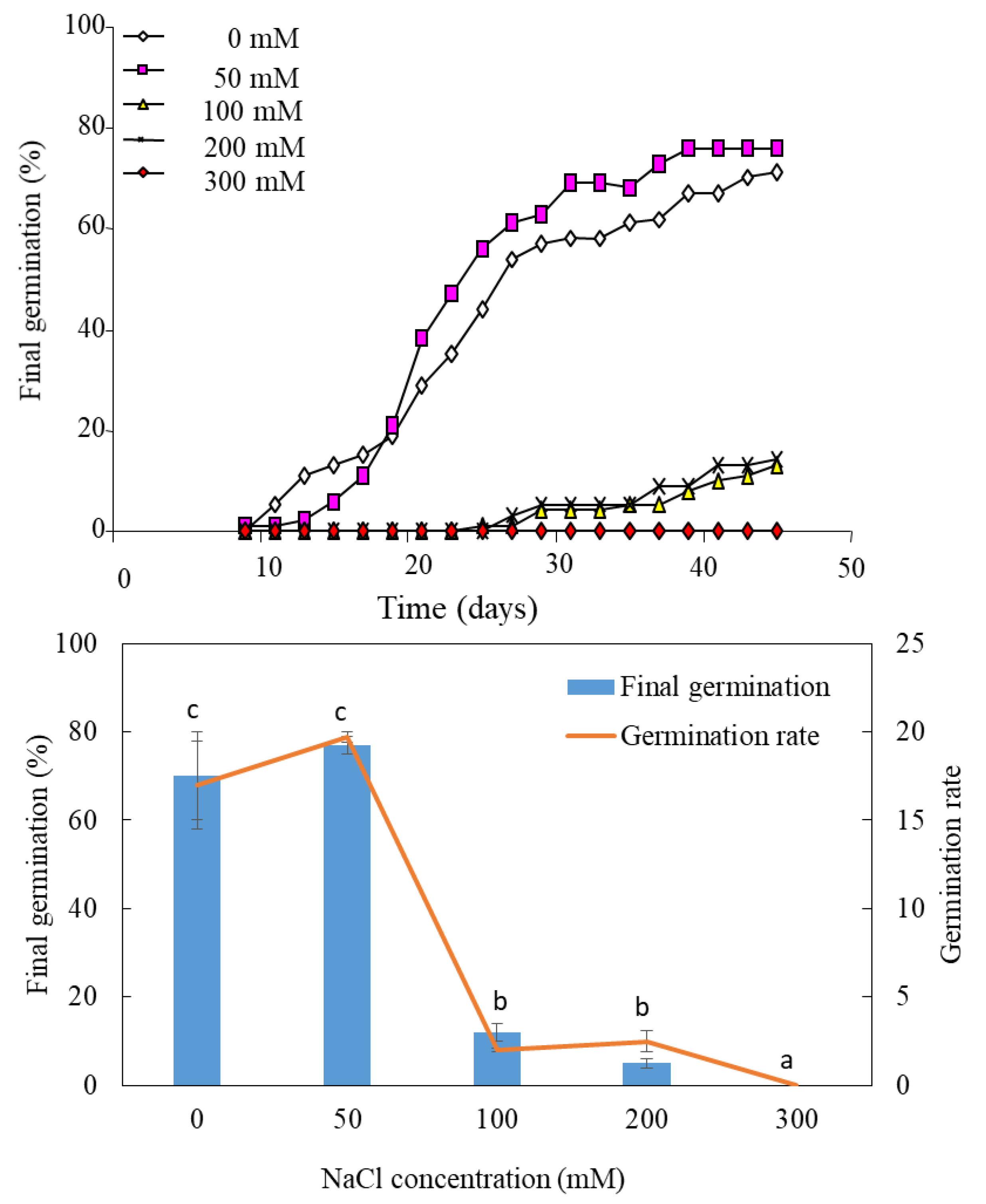
3.1. Effects of Salt Treatment on Final Germination
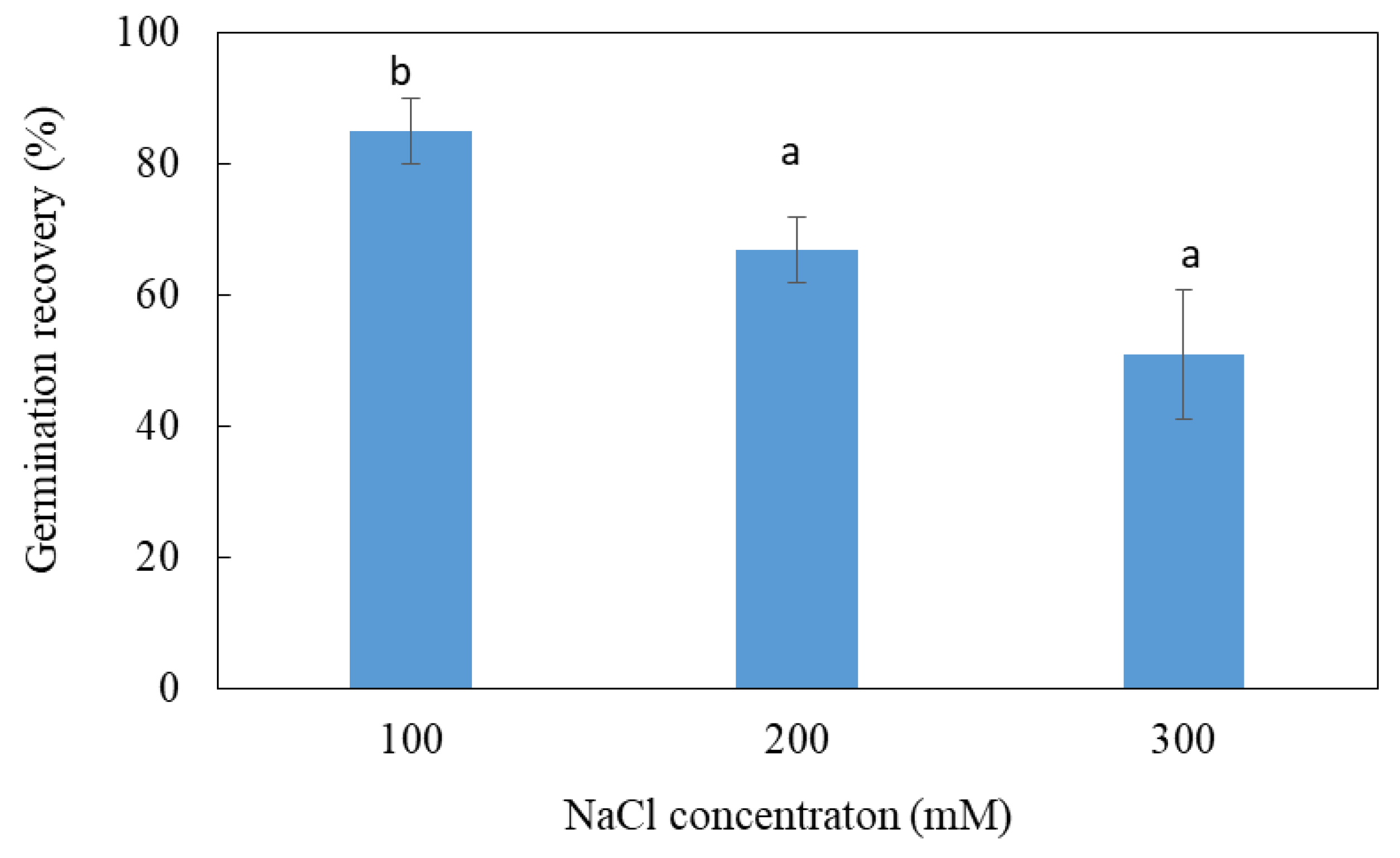
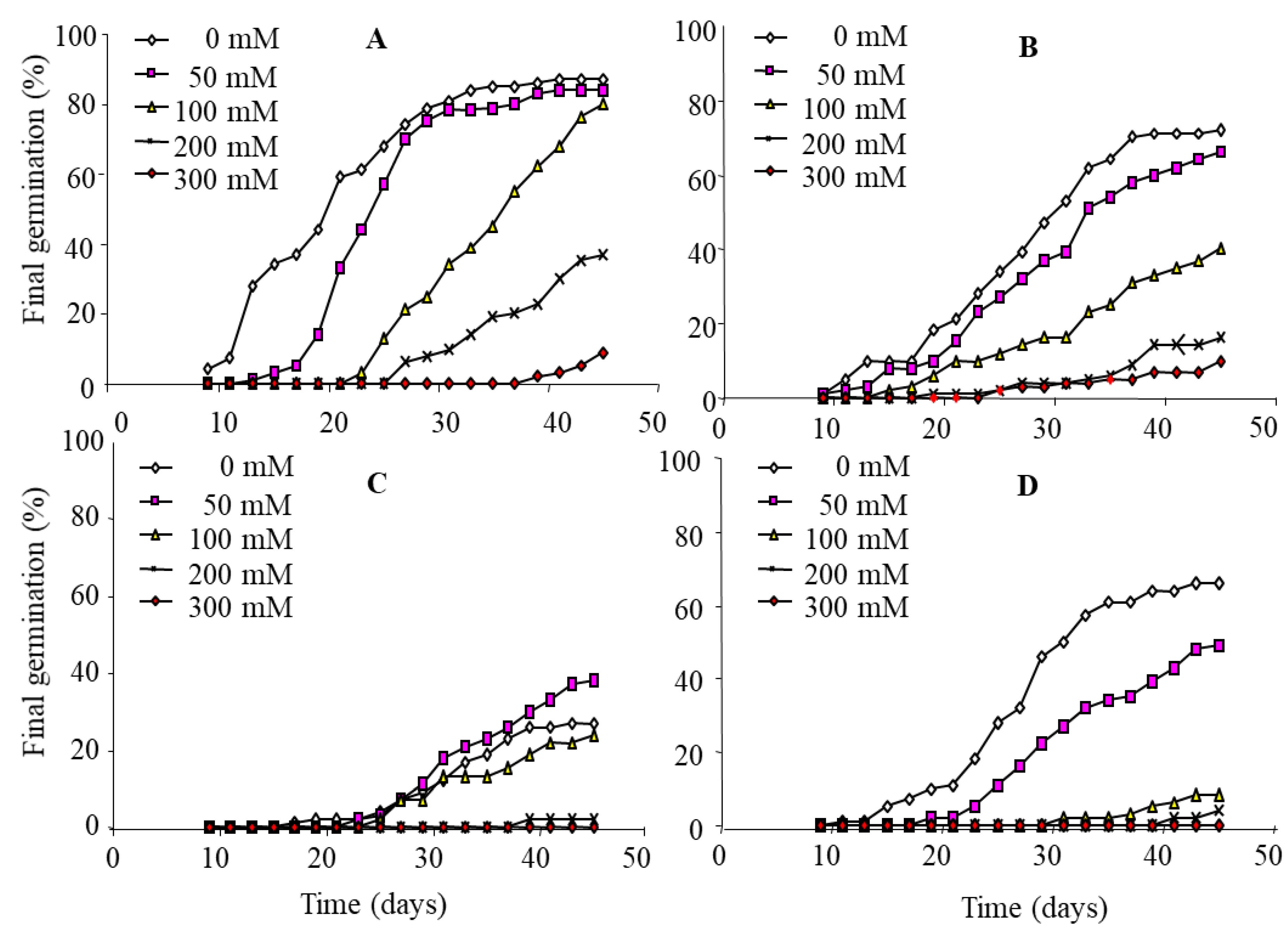
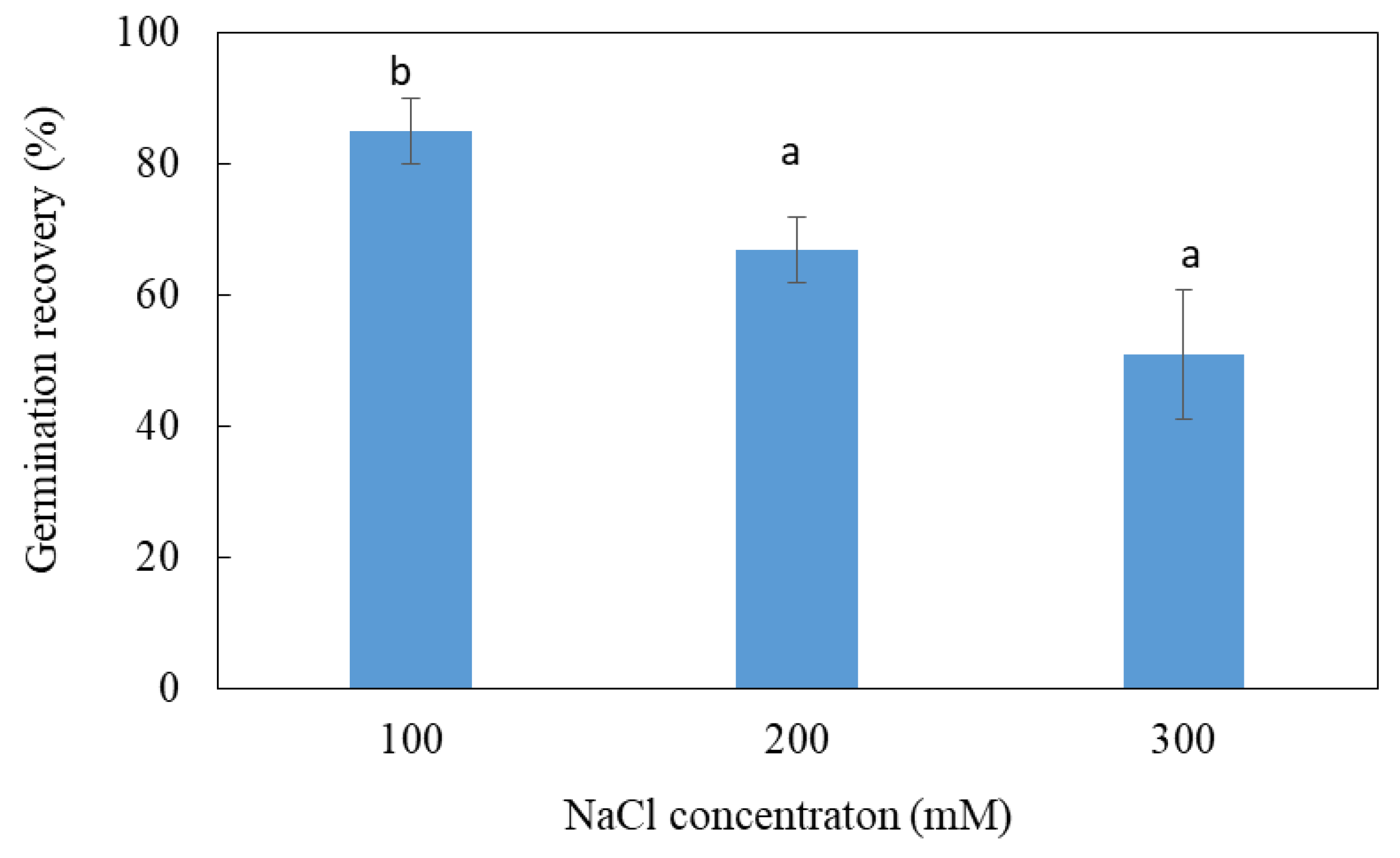
3.2. Effects of Salt Treatment on Germination Recovery
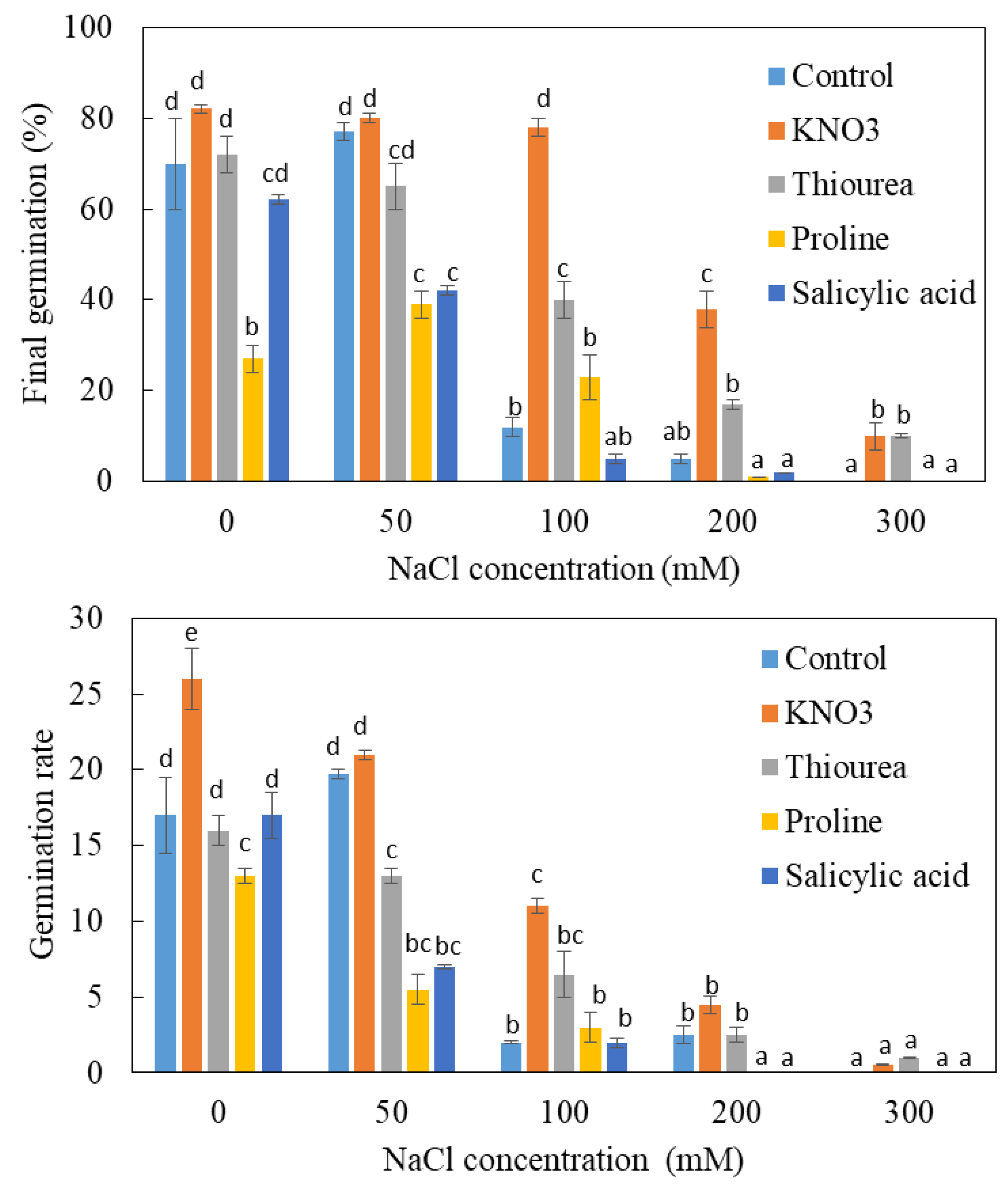
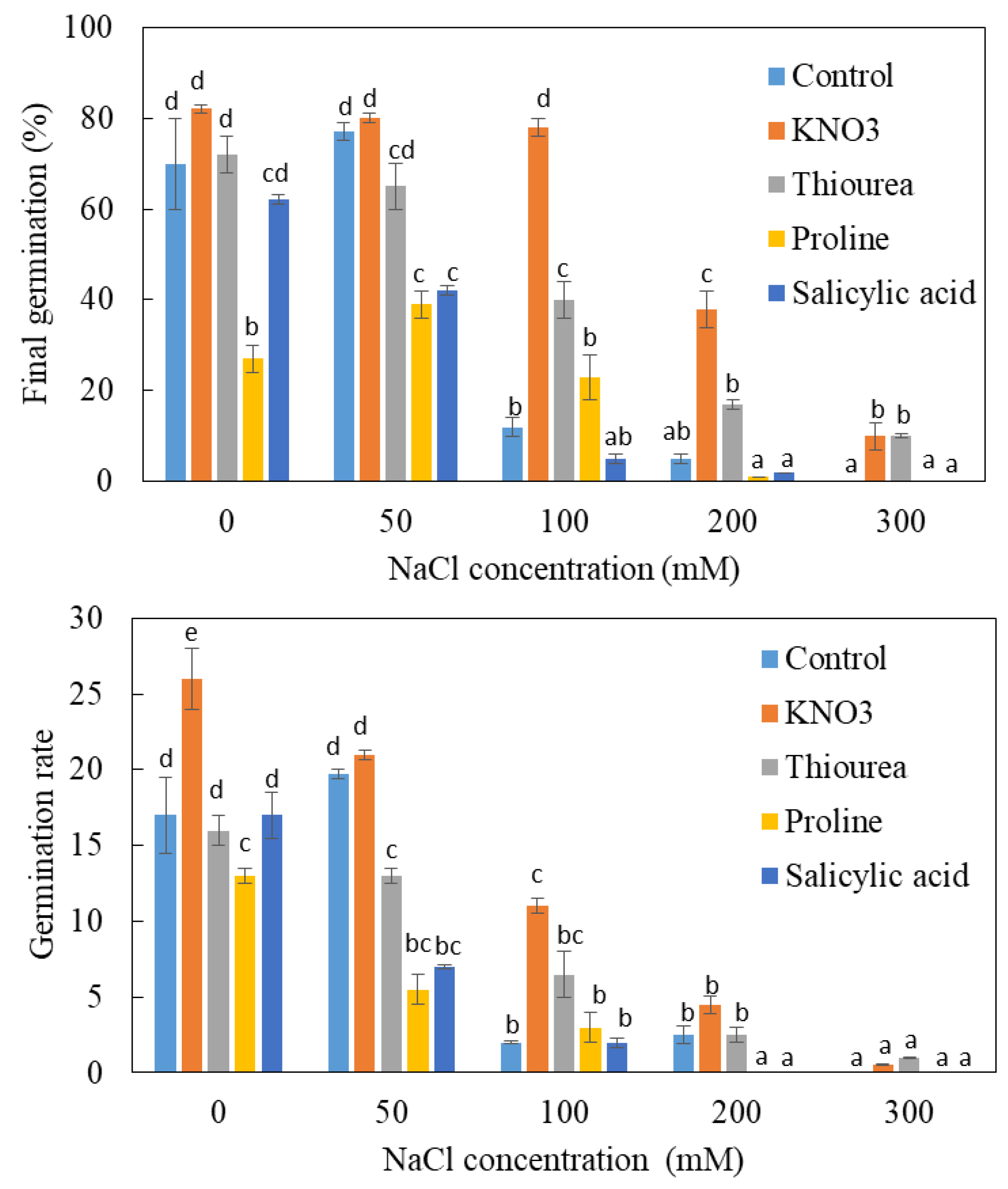
3.3. Effects of Chemical Priming on Germination under NaCl Salinity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, K.; Wu, Y.-H.; Tian, X.-Q.; Bai, Z.-Y.; Liang, Q.-Y.; Liu, Q.-L.; Pan, Y.-Z.; Zhang, L. Overexpression of DgWRKY4 enhances salt tolerance in chrysanthemum seedlings. Front. Plant Sci. 2017, 8, 1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Xie, L.; Sun, T.; Ding, B.; Li, Y.; Zhang, Y. Chrysanthemum morifolium aquaporin genes CmPIP1 and CmPIP2 are involved in tolerance to salt stress. Sci. Hortic. 2019, 256, 108627. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Llanderal, A.; Lao, M.T. Halophytes as an Option for the Restoration of Degraded Areas and Landscaping. In Handbook of Halophytes; Grigore, M.-N., Ed.; Springer Nature: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOPH a database of salt tolerant plants: Helping put halophytes to work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassaniti, C.; Romano, D. The use of halophytes for Mediterranean landscaping. Eur. J. Plant Sci. Biotechnol. 2011, 5, 58–63. [Google Scholar]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Gul, B.; Hameed, A.; Weber, D.J.; Khan, M.A. Assessing Seed Germination Responses of Great Basin Halophytes to Various Exogenous Chemical Treatments Under Saline Conditions. In Sabkha Ecosystems; Khan, M., Boër, B., Ȫzturk, M., Clüsener-Godt, M., Gul, B., Breckle, S.W., Eds.; Springer: Cham, Switzerland, 2016; Volume 48. [Google Scholar] [CrossRef]
- Pujol, J.A.; José, F.C.; Luis, R. Recovery germination from different osmotic conditions by four halophytes from South-eastern Spain. Ann. Bot. 2000, 85, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ungar, I.A. Action of compatible osmotica and growth regulators in alleviating the effect of salinity on the germination of dimorphic seeds of Arthrocneumum indicum. Int. J. Plant Sci. 1998, 159, 313–317. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Magné, C. Effect of salinity and chemical factors on seed germination in the halophyte Crithmum maritimum L. Plant Soil. 2008, 313, 83–87. [Google Scholar] [CrossRef]
- Atia, A.; Debez, A.; Barhoumi, Z.; Smaoui, A.; Abdelly, C. ABA, GA 3, and nitrate may control seed germination of Crithmum maritimum (Apiaceae) under saline conditions. Comptes Rendus Biol. 2009, 332, 704–710. [Google Scholar] [CrossRef]
- Atia, A.; Barhoum, Z.; Rabhi, M.; Abdely, C.; Smaoui, A. Environmental eco-physiology and economical potential of the halophyte Crithmum maritimum L. (Apiaceae). J. Med. Plants Res. 2011, 5, 3564–3571. [Google Scholar]
- Bouzidi, A.; Krouma, A.; Chaieb, M. Chemical seed priming alleviates salinity stress and improves Sulla carnosa germination in the saline depression of Tunisia. Plant Direct 2021, 5, e357. [Google Scholar] [CrossRef] [PubMed]
- Picó, F.X.; Retana, J. The flowering pattern of the perennial herb Lobularia maritima: An unusual case in the Mediterranean basin. Acta Oecologica 2001, 22, 209–217. [Google Scholar] [CrossRef]
- Brennan, E.B. Agronomic aspects of strip intercropping lettuce with alyssum for biological control of aphids. Biol. Control. 2013, 65, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Saville, D.J.; Wratten, S.D. Preferences of the wheat bug (Nysius huttoni) for particular growth stages of the potential trap crop, alyssum (Lobularia maritima). N. Z. Plant Prot. 2019, 72, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Gulzar, S.; Khan, M.A. Effects of salinity and temperature on the germination of Urochondra stulosa (trin.) C.E. Hubbard. Seed Sci. Technol. 2001, 29, 21–29. [Google Scholar]
- Debez, A.; Ben Hamed, K.; Grignon, C.; Abdelly, C. Salinity effects on germination, growth, and seed production of the halophyte Cakile maritima. Plant Soil 2004, 262, 179–189. [Google Scholar] [CrossRef]
- Keiffer, C.H.; Ungar, I.A. Germination responses of halophytes seeds exposed to prolonged hypersaline conditions. In Biology of Salt Tolerant Plants; Khan, M., Ungar, I.A., Eds.; Pakistan University of Karachi: Karachi, Pakistan, 1996; pp. 43–50. [Google Scholar]
- Plumer, J.A.; Rogers, A.D.; Turner, D.W.; Bell, D.T. Light, nitrogen compounds, smoke and GA 3 break dormancy and enhance germination in the Australian everlasting daisy, Shoenia filifolia Sbsp. Subulifolia. Seed Sci. Technol. 2001, 29, 321–330. [Google Scholar]
- Pandey, H.; Nandi, S.K.; Palni, L.M.S. Chemical stimulation of seed germination in Aconitum heterophyllum Wall. & A. Balfourii Stapf: Important Himalayan species of medicinal value. Seed Sci. Technol. 2000, 28, 39–48. [Google Scholar]
- Gul, B.; Weber, D.J. Effect of dormancy relieving compounds on the seed germination of non dormant Allenrolfea occidentalis under salinity stress. Ann. Bot. 1998, 82, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Ungar, I.A. Alleviation of innate and salinity–induced dormancy in Atriplex griffithii Moq. Var. Stocksii Boiss. Seeds Sci. Technol. 2000, 28, 29–37. [Google Scholar]
- Shim, S.I.; Moon, J.-C.; Jang, C.S.; Raymer, P.; Kim, W. Effect of potassium nitrate priming on seed germination of seashore paspalum. HortScience 2008, 43, 2259–2262. [Google Scholar] [CrossRef] [Green Version]
- Murru, V.; Santo, A.; Piaza, C.; Hugit, L.; Bacchetta, G. Seed germination, salt stress tolerance, and the effect of nitrate on three Tyrrnian coastal species of the Silene mollissima aggregate (Caryophyllaceae). Botany 2015, 93, 881–892. [Google Scholar] [CrossRef]
- Hilton, J.R.; Thomas, J.A. Regulation of pregerminative rates of respiration in seeds of various seed species by potassium nitrate. J. Exp. Bot. 1996, 37, 1516–1524. [Google Scholar] [CrossRef]
- Hilhorst, H.W.M.; Karseen, C.M. Dual Effect of light on the gibberelin and nitrate stimulated seed germination of Sisymbryom officinale and Arabidopsis thaliana. Plant Physiol. 1988, 86, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Hilhorst, H.W.M.; Karssen, M. Effect of chemical environment in seed germination. In The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International Wallingford: Wallingford, UK, 2000; pp. 293–311. [Google Scholar]
- Adkins, S.; Graham, M.; Simpson, G.; Naylor, J.M. The physiological basis of seed dormancy in Avena fatua. Physiolgia Plant. 1984, 60, 234–238. [Google Scholar] [CrossRef]
- Roberts, E.H.; Smith, R.D. Dormancy and the pentose phosphate pathway. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; Elsevier: Amsterdam, The Netherlands; North-Holland Biomedical Press: New York, NY, USA, 1977; pp. 385–411. [Google Scholar]
- Mani, S.; Van de cotte, B.; Van Montagu, N.; Verbruggen, N. Altered Levels of Proline Dehydrogenase Cause Hypersensivity to Proline and Its analogue in Arabidopsis. Plant Physiol. 2002, 128, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Poljakoff-Mayber, A.; Somers, G.F.; Werker, E.; Gallagher, J.L. Seeds germination of Kosteletzka virginica germination and salt tolerance. Am. J. Bot. 1994, 81, 54–59. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharment, J. NaCl-induced senescence in leavesof rice (Oryza sativa) cultivar differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Garcia, A.B.; De Almeider, E.J.; Iyer, S.; Gerats, T.; Van Mantagu, M.; Caplan, A.B. Effects of osmoprotectants upon NaCl stress in rice. Plant Physiol. 1997, 115, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Borsani OValpuesta, A.; Miguel, A.B. Evidence for a Role of Salicylic Acid in the Oxidative Damage Generated by and Osmotic Stress in Arabidopsis Seedlings. Plant Physiol. 2001, 126, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Takaki, M.; Rosim, R.E. Aspirin increases tolerance to high temperature in seeds of Raphanus sativus L. cVAR Early Scarlet Globe. Seed Sci. Technol. 2000, 28, 179–183. [Google Scholar]
- Janda, T.; Sazalai, G.; Tari, I.; Pàldi, E. Hydroponic treatement with salisalic acid decreases the effects of chiling injury in maize.(Zea mays L.) Plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Hongua, C.; Leynan, T.; Junmei, S.; Xiaou, H.; Xianguo, C. Exogenous salicylic acid reveals an osmotic regulatory role in priming the seed germination of Leymus chinensis under salt-alkali stress. Environ. Exp. Bot. 2021, 18, 104498. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zammali, I.; Dabbous, A.; Youssef, S.; Ben Hamed, K. Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity. Seeds 2022, 1, 99-109. https://doi.org/10.3390/seeds1020009
Zammali I, Dabbous A, Youssef S, Ben Hamed K. Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity. Seeds. 2022; 1(2):99-109. https://doi.org/10.3390/seeds1020009
Chicago/Turabian StyleZammali, Iman, Amira Dabbous, Seria Youssef, and Karim Ben Hamed. 2022. "Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity" Seeds 1, no. 2: 99-109. https://doi.org/10.3390/seeds1020009
APA StyleZammali, I., Dabbous, A., Youssef, S., & Ben Hamed, K. (2022). Effects of Chemical Priming on the Germination of the Ornamental Halophyte Lobularia maritima under NaCl Salinity. Seeds, 1(2), 99-109. https://doi.org/10.3390/seeds1020009