
From Cone to Seed and Seedling—Characterization of Three Portuguese Pinus pinaster Aiton Populations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Characterization
2.2. Climate Data
2.3. Cone Sampling and Processing
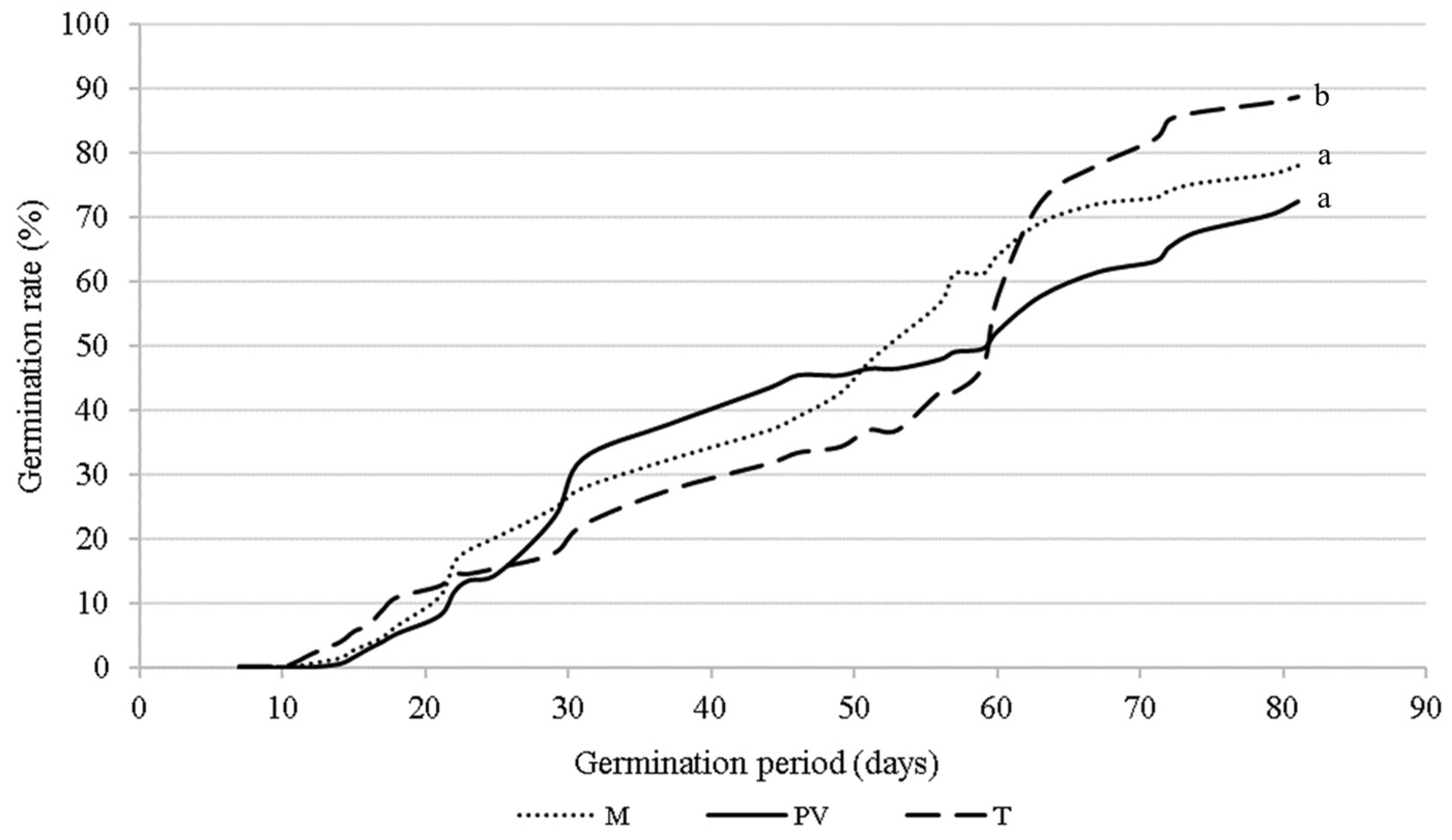
2.4. Seed Germination
2.5. Seedling Nursing and Biometric Traits
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Managing Forests for Climate Change. 2010. Available online: https://www.fao.org/3/i1960e/i1960e00.pdf (accessed on 28 February 2022).
- Gordo, F.J.; Montero, G.; Gil, L. La Problemática de la Regeneración Natural de los Pinares en los Arenales de la Meseta Castellana; Instituto Universitario de Investigación en Gestión Forestal Sostenible: Palencia, Spain, 2012. [Google Scholar]
- Calama, R.; Manso, R.; Lucas-Borja, M.E.; Espelta, J.M.; Piqué, M.; Bravo, F.; Del Peso, C.; Pardos, M. Natural regeneration in Iberian pines: A review of dynamic processes and proposals for management. For. Syst. 2017, 26, eR02S. [Google Scholar] [CrossRef]
- Calvo, L.; Hernández, V.; Valbuena, L.; Taboada, A. Provenance and seed mass determine seed tolerance to high temperatures associated to forest fires in Pinus pinaster. Ann. For. Sci. 2015, 73, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Alía, R.; Chambel, R.; Notivol, E.; Climent, J.; González-Martínez, S.C. Environment-dependent microevolution in a Mediterranean pine (Pinus pinaster Aiton). BMC Evol. Biol. 2014, 14, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausas, J.G.; Bladé, C.; Valdecantos, A.; Seva, J.P.; Fuentes, D.; Alloza, J.A.; Vilagrosa, A.; Bautista, S.; Cortina, J.; Vallejo, R. Pines and oaks in the restoration of Mediterranean landscapes of Spain: New perspectives for an old practice—A review. Plant Ecol. 2004, 171, 209–220. [Google Scholar] [CrossRef]
- Proença, V.; Pereira, H.M.; Vicente, L. Resistance to wildfire and early regeneration in natural broadleaved forest and pine plantation. Acta Oecologica 2010, 36, 626–633. [Google Scholar] [CrossRef]
- Maia, P.; Pausas, J.; Vasques, A.; Keizer, J.J. Fire severity as a key factor in post-fire regeneration of Pinus pinaster (Ait.) in Central Portugal. Ann. For. Sci. 2012, 69, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Charco, J.; Venturas, M.; Gil, L.; Nanos, N. Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources. Forests 2017, 8, 312. [Google Scholar] [CrossRef] [Green Version]
- Tapias, R.; Climent, J.; Pardos, J.A.; Gil, L. Life histories of Mediterranean pines. Plant Ecol. 2004, 171, 53–68. [Google Scholar] [CrossRef]
- Juez, L.; González-Martínez, S.C.; Nanos, N.; De-Lucas, A.I.; Ordóñez, C.; del Peso, C.; Bravo, F. Can seed production and restricted dispersal limit recruitment in Pinus pinaster Aiton from the Spanish Northern Plateau? For. Ecol. Manag. 2014, 313, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, V.R. Are wildfires a disaster in the Mediterranean basin? A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Budde, K.B.; Heuertz, M.; Hernández-Serrano, A.; Pausas, J.G.; Vendramin, G.G.; Verdú, M.; González-Martínez, S.C. In situ genetic association for serotiny, a fire-related trait, in Mediterranean maritime pine (Pinus pinaster). New Phytol. 2013, 201, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, I.; Alía, R.; Yan, W.; David, T.; Aguiar, A.; Almeida, M.H. Genotype × Environment interactions in Pinus pinaster at age 10 in a multienvironment trial in Portugal: A maximum likelihood approach. Ann. For. Sci. 2010, 67, 612. [Google Scholar] [CrossRef]
- Correia, I.; Santos, L.; Faria, C.; Nóbrega, C.; Almeida, H.; David, T. Cone to Seedling—Variation between Pinus pinaster Provenances from Contrasting Altitudes. For. Sci. 2014, 60, 724–732. [Google Scholar] [CrossRef]
- Wahid, N.; Bounoua, L. The relationship between seed weight, germination and biochemical reserves of maritime pine (Pinus pinaster Ait.) in Morocco. New For. 2012, 44, 385–397. [Google Scholar] [CrossRef]
- Fernández-García, V.; Fulé, P.Z.; Marcos, E.; Calvo, L. The role of fire frequency and severity on the regeneration of Mediterranean serotinous pines under different environmental conditions. For. Ecol. Manag. 2019, 444, 59–68. [Google Scholar] [CrossRef]
- Marques, S.; Garcia-Gonzalo, J.; Botequim, B.; Ricardo, A.; Borges, J.G.; Tomé, M.; Oliveira, M. Assessing wildfire occurrence probability in Pinus pinaster Ait. stands in Portugal. For. Syst. 2012, 21, 111. [Google Scholar] [CrossRef] [Green Version]
- Bravo, F.; Maguire, D.A.; González-Martínez, S.C. Factors affecting cone production in Pinus pinaster Ait.: Lack of growth-reproduction trade-offs but significant effects of climate and tree and stand characteristics. For. Syst. 2017, 26, e07S. [Google Scholar] [CrossRef] [Green Version]
- Zas, R.; Cendán, C.; Sampedro, L. Mediation of seed provisioning in the transmission of environmental maternal effects in Maritime pine (Pinus pinaster Aiton). Heredity 2013, 111, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Zas, R.; Sampedro, L. Heritability of seed weight in Maritime pine, a relevant trait in the transmission of environmental maternal effects. Heredity 2014, 114, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Escudero, A.; Núñez, Y.; Pérez-García, F. Is fire a selective force of seed size in pine species? Acta Oecologica 2000, 21, 245–256. [Google Scholar] [CrossRef]
- Reyes, O.; Casal, M. The influence of seed age on germinative response to the effects of fire in Pinus pinaster, Pinus radiata and Eucalyptus globulus. Ann. For. Sci. 2001, 58, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Serrano, A.; Verdu, M.; González-Martínez, S.C.; Pausas, J.G. Fire structures pine serotiny at different scales. Am. J. Bot. 2013, 100, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Vidal, E.; Sampedro, L.; Zas, R. Is the benefit of larger seed provisioning on seedling performance greater under abiotic stress? Environ. Exp. Bot. 2017, 134, 45–53. [Google Scholar] [CrossRef] [Green Version]
- ICNF. 6.o Inventário Floresta Nacional. 2015. Available online: http://www2.icnf.pt/portal/florestas/ifn/resource/doc/ifn/ifn6/IFN6_Relatorio_completo-2019-11-28.pdf (accessed on 1 February 2022).
- Almeida, A.F.; Capelo, J.; Mesquita, S. Mata Nacional de Leiria: Indicadores Fitoecológicos. Silva Lusit 2002, 10, 195–200. [Google Scholar]
- Freire, J.; Tavares, M.; Campos, J. Ritmos de Crescimento das Espécies Pinus pinaster, Acacia melanoxylon e Acacia dealbata nas Dunas do Litoral Norte e Centro. Silva Lusit 2003, 11, 67–76. [Google Scholar]
- Fabião, A.; Fabião, A.; Carneiro, M.; Pereira, F.; Cancela, J.H.; Pimentel, F. Rehabilitation of woody riparian vegetation in the margins of the Linhos lagoon, at Urso National Forest, Rev. Ciências Agrárias 2006, 29, 90–101. [Google Scholar]
- Guerra, S. Flora e habitats da zona costeira da Mata Nacional de Leiria; MsC Dissertation, Universidade de Aveiro: Aveiro, Portugal, 2011; Available online: https://ria.ua.pt/handle/10773/8585 (accessed on 1 February 2022).
- Oliveira, M. Determinantes do Desenvolvimento do Pinhal Bravo em Áreas Dunares (Dunas de Mira), 2014. Ph.D. Thesis, Universidade de Coimbra, Coimbra, Portugal, 2014. [Google Scholar]
- IPMA. Clima Normais. 2020. Available online: https://www.ipma.pt/pt/oclima/normais.clima/1971-2000/normalclimate7100.jsp (accessed on 1 February 2022).
- Marques, P. Os Solos da Mata Nacional de Leiria: Características e Classificação; Instituto Superior de Agronomia: Lisboa, Portuguese, 2010. [Google Scholar]
- IPMA. Boletim Climatológico Anual Portugal Continental. 2017. Available online: https://www.ipma.pt/resources.www/docs/im.publicacoes/edicoes.online/20180315/GLoWcBXPqGauRNYDpUAC/cli_20171201_20171231_pcl_aa_co_pt.pdf (accessed on 1 February 2022).
- IPMA. Boletim Climatológico Anual Portugal Continental. 2018. Available online: https://www.ipma.pt/resources.www/docs/im.publicacoes/edicoes.online/20190131/ddliTNijyEWyfrhUMDTy/cli_20180901_20181231_pcl_aa_co_pt.pdf (accessed on 1 February 2022).
- IPMA. Boletim Climatológico Anual Portugal Continental. 2019. Available online: https://www.ipma.pt/resources.www/docs/im.publicacoes/edicoes.online/20200318/piMisHmKRkiueuXQgHpM/cli_20191201_20191231_pcl_aa_co_pt.pdf (accessed on 1 February 2022).
- Cendán, C.; Sampedro, L.; Zas, R. The maternal environment determines the timing of germination in Pinus pinaster. Environ. Exp. Bot. 2013, 94, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Wahid, N.; El Hadrami, I.; Boulli, A. Variabilité du potentiel germinatif des semences de certaines populations marocaines de pin maritime (Pinus pinaster Ait.) sous conditions contrôlées. Acta Bot. Gallica 2007, 154, 7–19. [Google Scholar] [CrossRef]
- Alvarez, R.; Valbuena, L.; Calvo, L. Effect of high temperatures on seed germination and seedling survival in three pine species (Pinus pinaster, P. sylvestris and P. nigra). Int. J. Wildland Fire 2007, 16, 63–70. [Google Scholar] [CrossRef]
- Valbuena, L.; Taboada, A.; Tárrega, R.; De la Rosa, A.; Calvo, L. Germination response of woody species to laboratory-simulated fire severity and airborne nitrogen deposition: A post-fire recovery strategy perspective. Plant Ecol. 2019, 220, 1057–1069. [Google Scholar] [CrossRef]
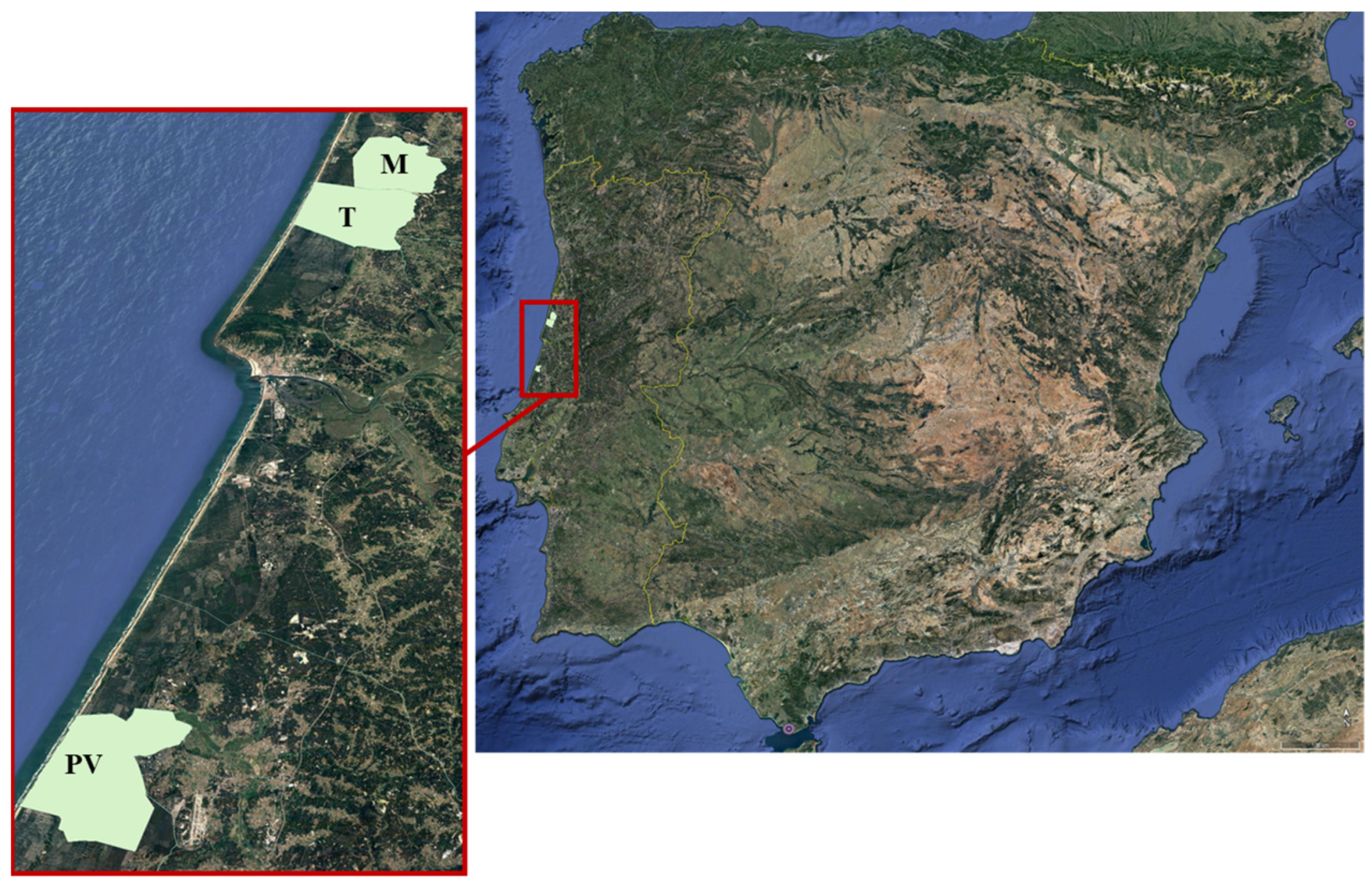

{kind=link}
{kind=link}
| Site | Year | Precip (mm/year) | Mean Temp (°C) | Temp Max (°C) | Temp Min (°C) | Max Days without Rain |
|---|---|---|---|---|---|---|
| M | 1 | 1115 | 15.3 ± 4.6 | 40.8 | −0.9 | 98 |
| 2 | 960 | 15.5 ± 3.9 | 35.8 | 0.1 | 30 | |
| T | 1 | 1082 | 15.2 ± 4.5 | 40.5 | 0.2 | 98 |
| 2 | 838 | 15.6 ± 3.9 | 35.4 | 1.0 | 30 | |
| PV | 1 | 952 | 15.4 ± 4.7 | 39.9 | 1.6 | 100 |
| 2 | 742 | 15.7 ± 4.1 | 34.9 | 2.2 | 35 |
| Population | Cone Mass (g) | Seeds Per Cone | Mean Seed Mass (g) | Germination Rate (%) |
|---|---|---|---|---|
| M | 100.3 ± 26.9 a | 135.7 ± 30.4 a | 0.056 ± 0.01 a | 76.7 b |
| T | 89.1 ± 28.6 b | 80.4 ± 43.5 b | 0.059 ± 0.01 b | 88.9 a |
| PV | 88.6 ± 26.8 b | 116.3 ± 27.5 c | 0.054 ± 0.01 c | 72.5 b |
| Estimate | Std. Error | z Value | p (>|z|) | |
|---|---|---|---|---|
| Intercept | −0.566251 | 0.367505 | −1.738 | 0.082162 |
| Mean seed mass | 16.277369 | 6.389929 | 2.547 | 0.010855 * |
| Mean cone mass | 0.010359 | 0.002757 | 3.757 | 0.000172 * |
| T population (T) | 0.906588 | 0.189284 | 4.790 | 1.67 × 10−6 * |
| Goodness of fit test: Χ2 = 8.47; df = 8 | 0.39 | |||
| Population | T Height (cm) | S Height (cm) | bsd (mm) | Ratio T Height/bsd |
|---|---|---|---|---|
| M | 21.34 ± 2.94 a | 18.18 ± 3.01a | 3.52 ± 0.37 a | 6.11 ± 0.95 a |
| T | 21.06 ± 3.01 a | 18.12 ± 3.11a | 3.51 ± 0.46 a | 5.69 ± 1.09 a |
| PV | 19.66 ± 2.90 b | 16.71 ± 2.76 b | 3.52 ± 0.37 a | 6.02 ± 0.88 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maia, P.; Corticeiro, S. From Cone to Seed and Seedling—Characterization of Three Portuguese Pinus pinaster Aiton Populations. Seeds 2022, 1, 126-135. https://doi.org/10.3390/seeds1020011
Maia P, Corticeiro S. From Cone to Seed and Seedling—Characterization of Three Portuguese Pinus pinaster Aiton Populations. Seeds. 2022; 1(2):126-135. https://doi.org/10.3390/seeds1020011
Chicago/Turabian StyleMaia, Paula, and Sofia Corticeiro. 2022. "From Cone to Seed and Seedling—Characterization of Three Portuguese Pinus pinaster Aiton Populations" Seeds 1, no. 2: 126-135. https://doi.org/10.3390/seeds1020011
APA StyleMaia, P., & Corticeiro, S. (2022). From Cone to Seed and Seedling—Characterization of Three Portuguese Pinus pinaster Aiton Populations. Seeds, 1(2), 126-135. https://doi.org/10.3390/seeds1020011