
Comparing Brain Responses to Different Styles of Music through Their Real and Imagined Interpretation: An Analysis Based on EEG Connectivity Networks †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
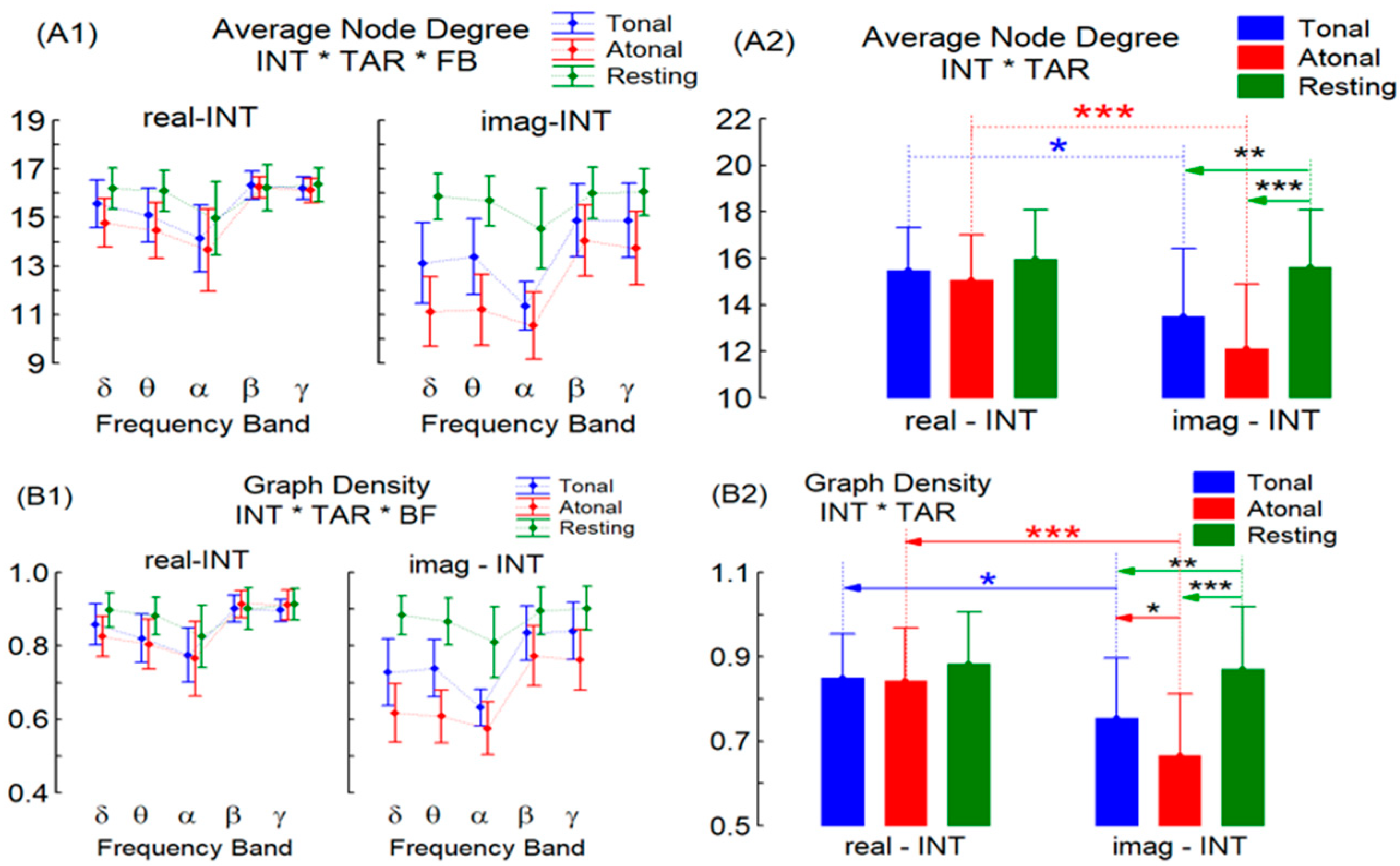
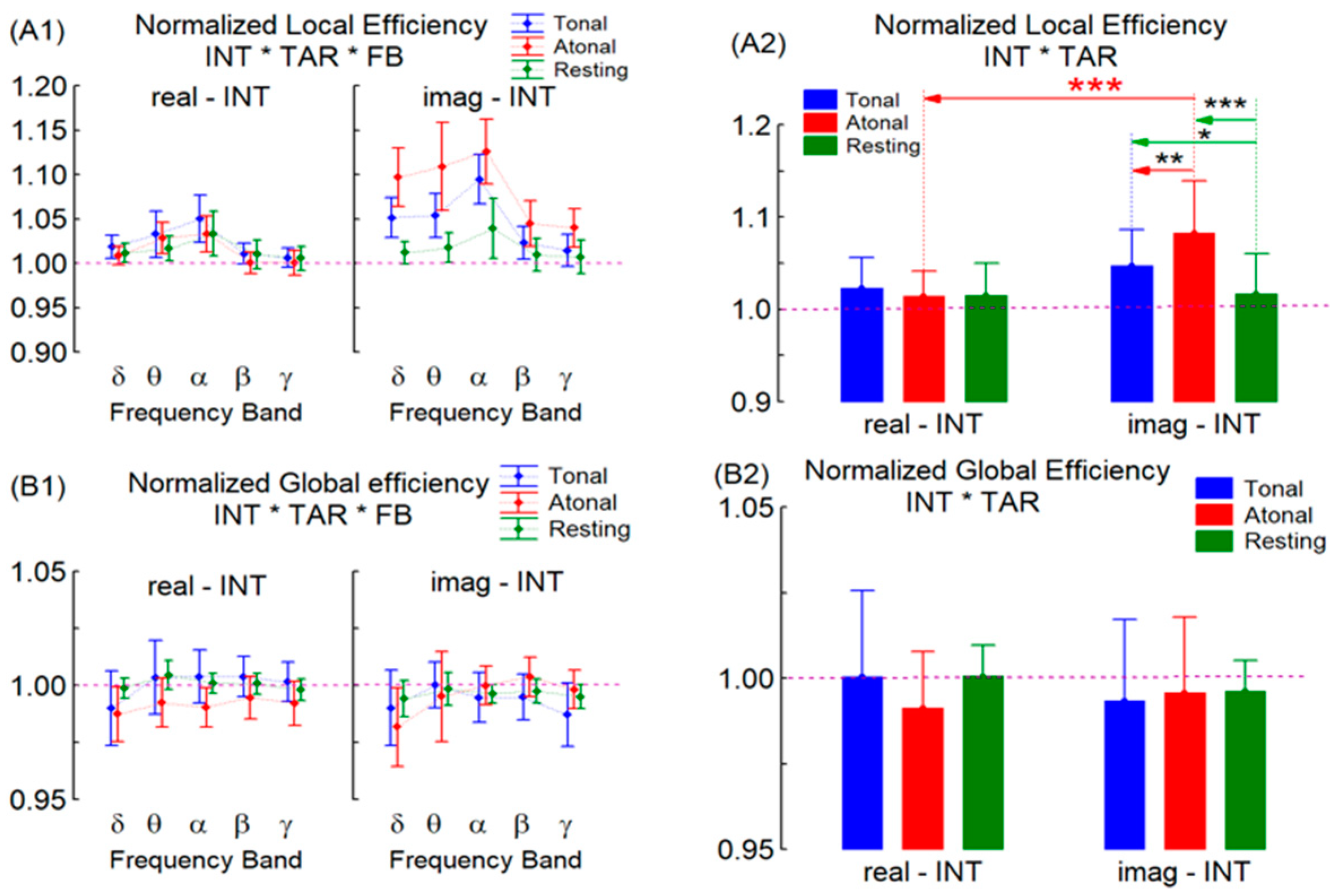
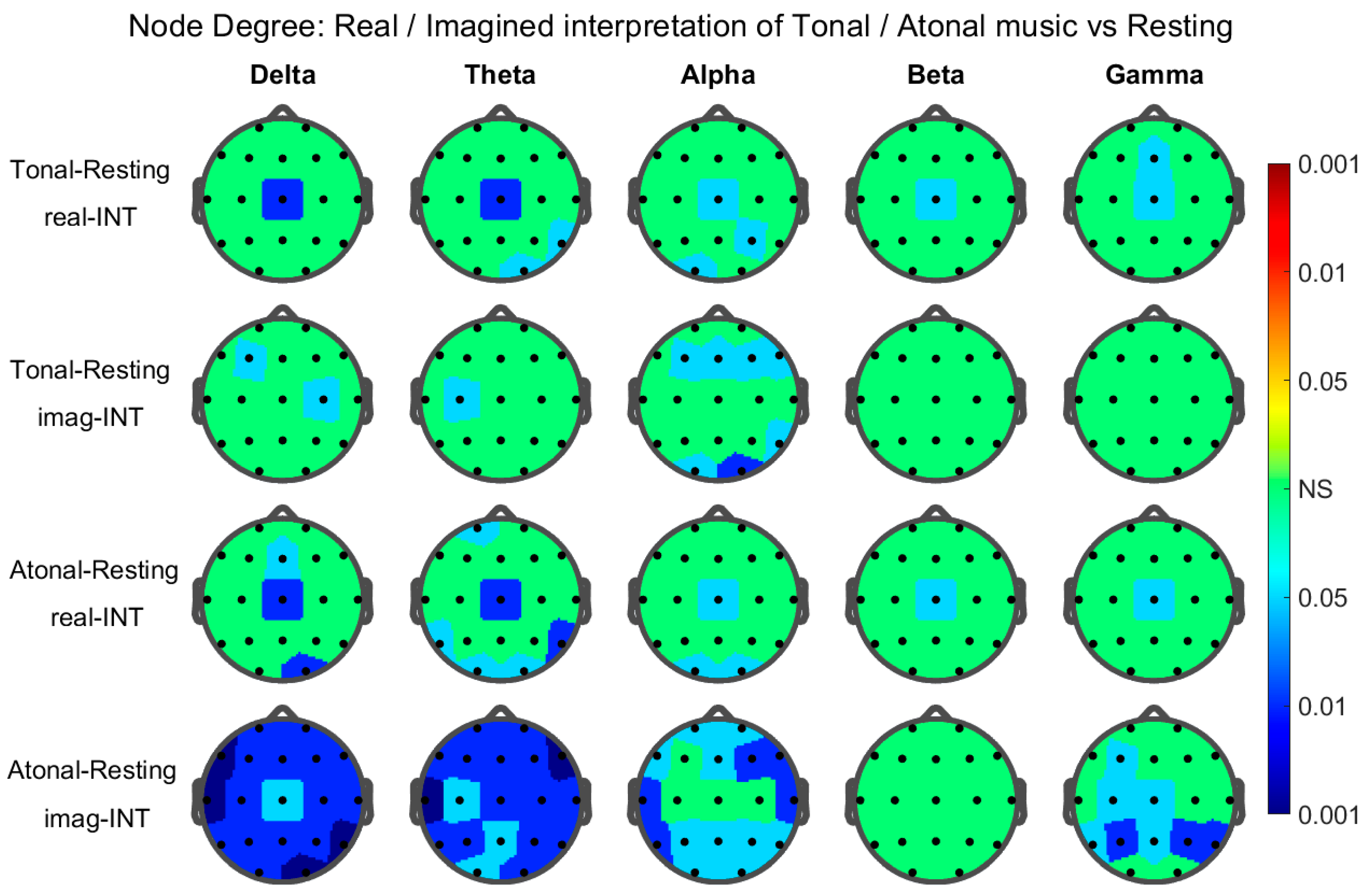
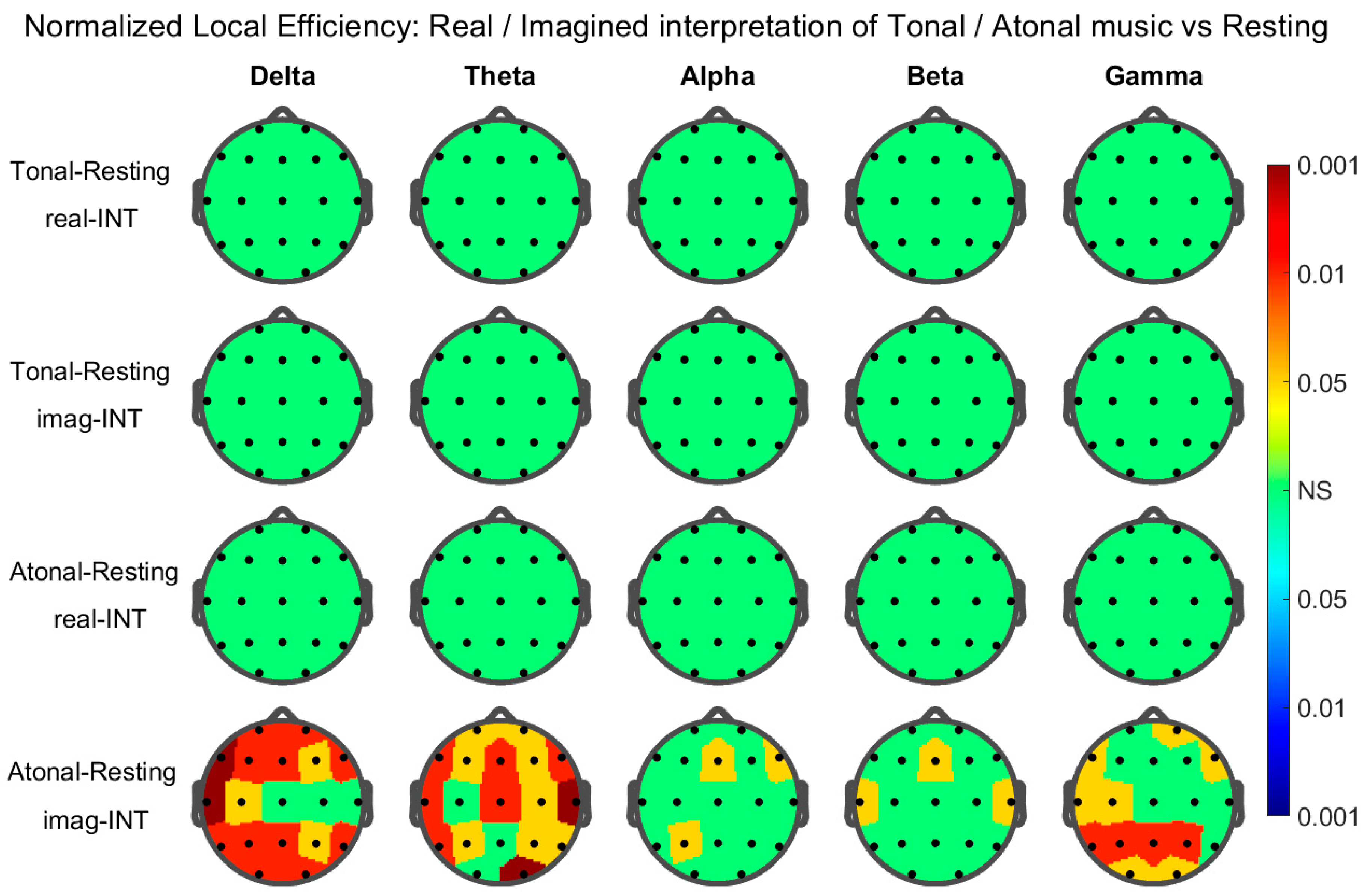
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaefer, R.S.; Morcom, A.M.; Roberts, N.; Overy, K. Moving to music: Effects of heard and imagined musical cues on movement-related brain activity. Front. Hum. Neurosci. 2014, 8, 774. [Google Scholar] [CrossRef] [PubMed]
- Zatorre, R.J.; Halpern, A.R. Mental Concerts: Musical Imagery and Auditory Cortex. Neuron 2005, 47, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Herholz, S.C.; Lappe, C.; Knief, A.; Pantev, C. Neural basis of music imagery and the effect of musical expertise. Eur. J. Neurosci. 2008, 28, 2352–2360. [Google Scholar] [CrossRef] [PubMed]
- Herholz, S.C.; Zatorre, R.J. Musical training as a framework for brain plasticity: Behavior, function, and structure. Neuron 2012, 76, 486–502. [Google Scholar] [CrossRef] [PubMed]
- Kristeva, R.; Chakarov, V.; Schulte-Mönting, J.; Spreer, J. Activation of cortical areas in music execution and imagining: A high-resolution EEG study. Neuroimage 2003, 20, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, R.S.; Vlek, R.J.; Desain, P. Music perception and imagery in EEG: Alpha band effects of task and stimulus. Int. J. Psychophysiol. 2011, 82, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Herholz, S.C.; Halpern, A.R.; Zatorre, R.J. Neuronal correlates of perception, imagery, and memory for familiar tunes. J. Cogn. Neurosci. 2012, 24, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, D.J.; Macrae, C.N.; Green, A.E.; Kelley, W.M. Sound of silence activates auditory cortex. Nature 2005, 434, 158. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kirino, E. Dynamic reconfiguration of the supplementary motor area network during imagined music performance. Front. Hum. Neurosci. 2017, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Kirino, E. Increased functional connectivity of the angular gyrus during imagined music performance. Front. Hum. Neurosci. 2019, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Manuel Santapau, M.; Gamundí, A.; Pereda, E.; González, J.J. Modifications in the Topological Structure of EEG Functional Connectivity Networks during Listening Tonal and AtonalConcert Music in Musicians and Non-Musicians. Brain Sci. 2021, 11, 159. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, A.; Modroño, C.; Santapau, M.; González, J.J. Comparing Brain Responses to Different Styles of Music through Their Real and Imagined Interpretation: An Analysis Based on EEG Connectivity Networks. Med. Sci. Forum 2022, 8, 13. https://doi.org/10.3390/IECBS2021-10667
González A, Modroño C, Santapau M, González JJ. Comparing Brain Responses to Different Styles of Music through Their Real and Imagined Interpretation: An Analysis Based on EEG Connectivity Networks. Medical Sciences Forum. 2022; 8(1):13. https://doi.org/10.3390/IECBS2021-10667
Chicago/Turabian StyleGonzález, Almudena, Cristián Modroño, Manuel Santapau, and Julián J. González. 2022. "Comparing Brain Responses to Different Styles of Music through Their Real and Imagined Interpretation: An Analysis Based on EEG Connectivity Networks" Medical Sciences Forum 8, no. 1: 13. https://doi.org/10.3390/IECBS2021-10667
APA StyleGonzález, A., Modroño, C., Santapau, M., & González, J. J. (2022). Comparing Brain Responses to Different Styles of Music through Their Real and Imagined Interpretation: An Analysis Based on EEG Connectivity Networks. Medical Sciences Forum, 8(1), 13. https://doi.org/10.3390/IECBS2021-10667