
Niphargus carolinensis sp. nov. (Amphipoda: Niphargidae), an Endemic Species in the Northern Distribution Area of the Genus



Abstract
1. Introduction
2. Materials and Methods
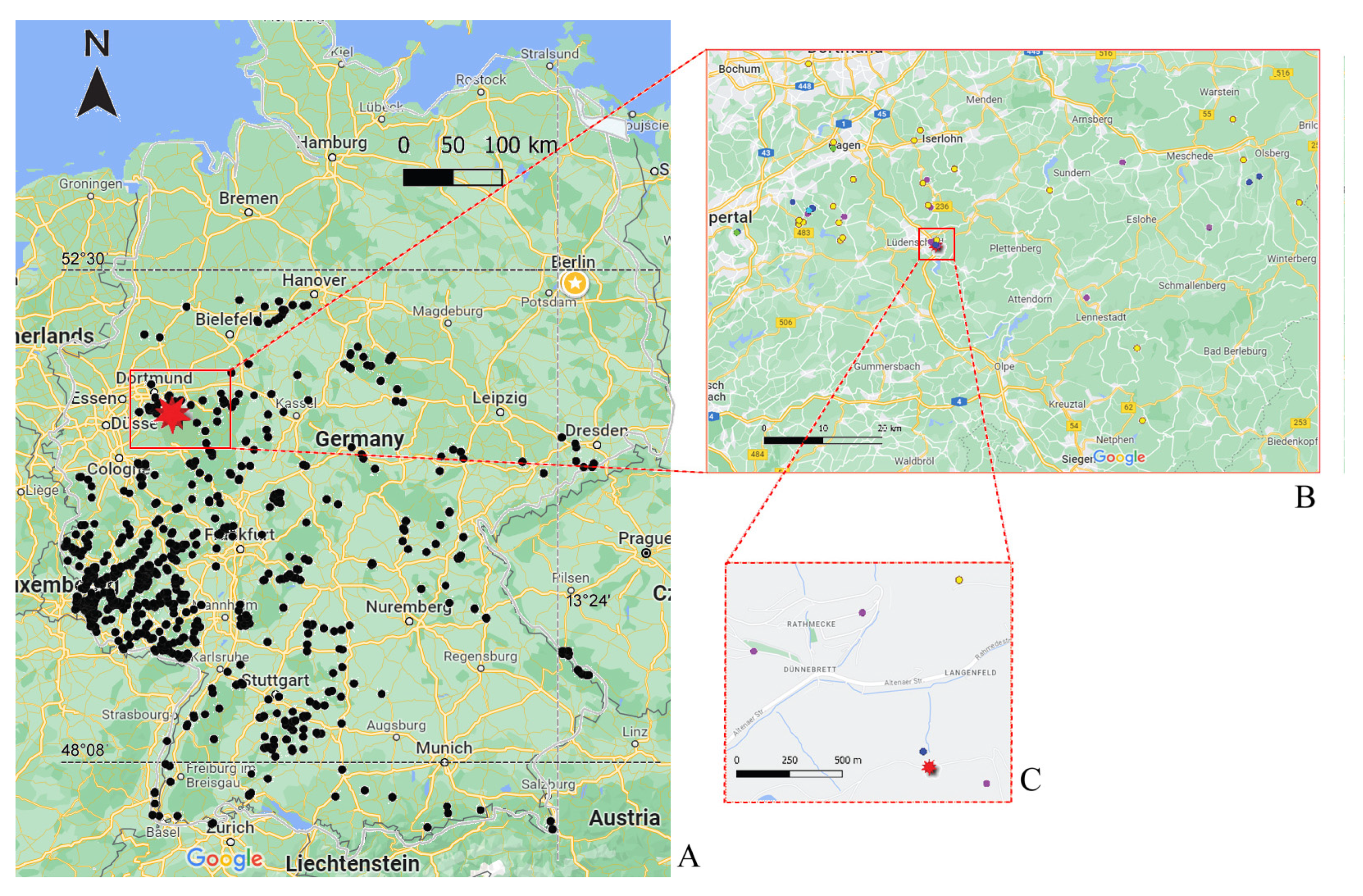
2.1. Sampling
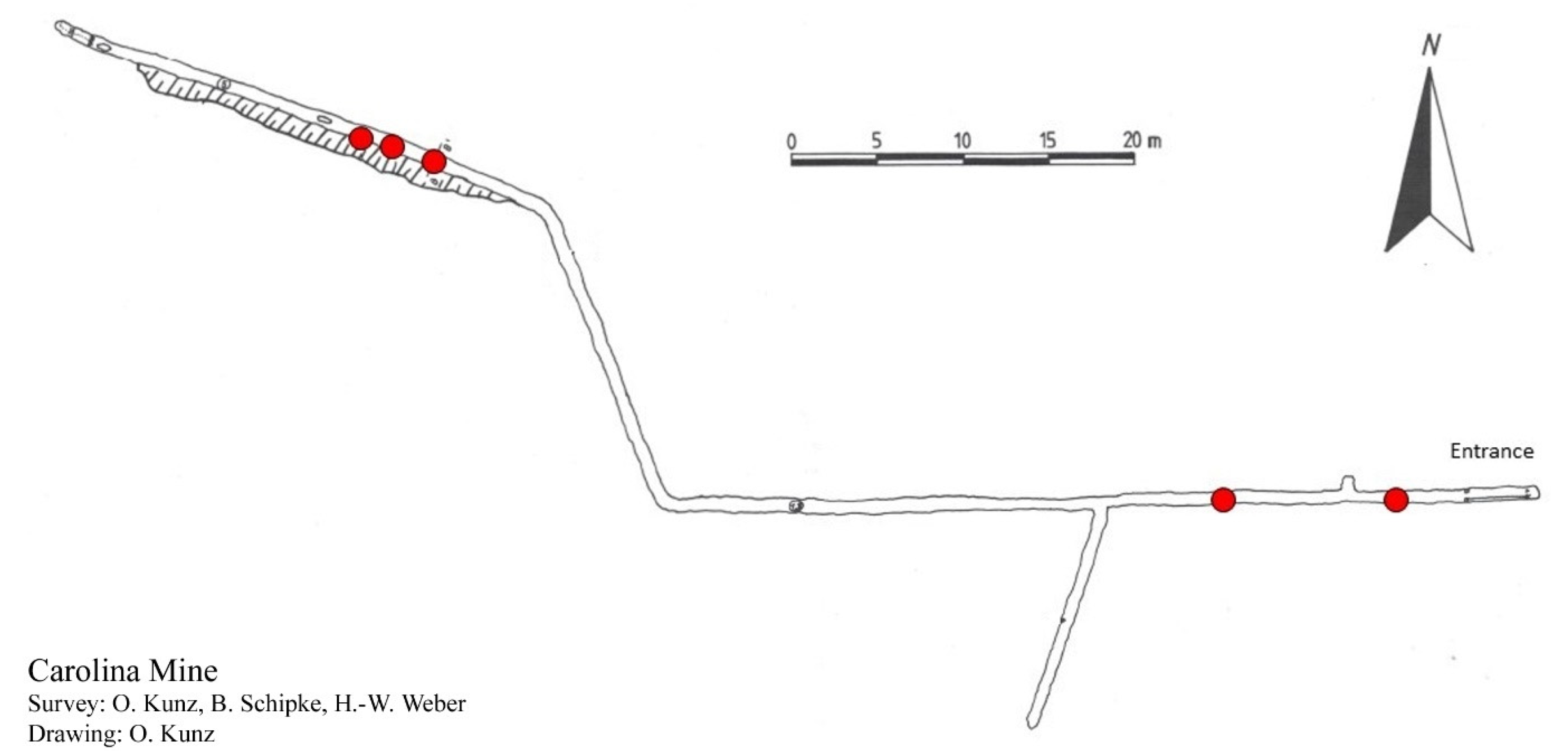
2.2. Sampling Site
2.3. Laboratory Work
2.4. Sequence Analysis
2.5. Morphological Analysis
3. Results
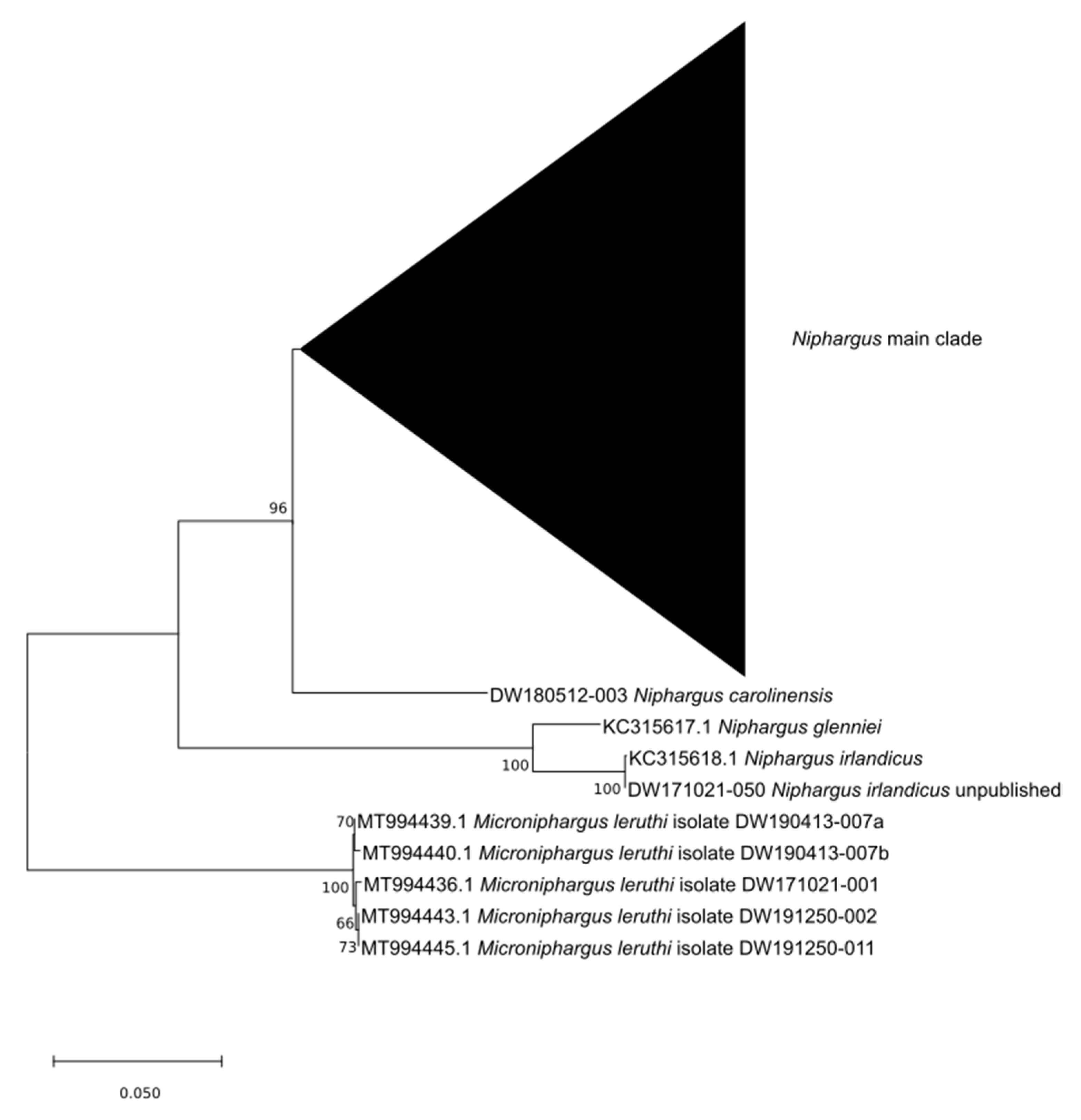
3.1. Molecular Identification
3.2. Morphology-Based Description
3.2.1. Diagnosis
3.2.2. Etymology
3.2.3. Type Locality
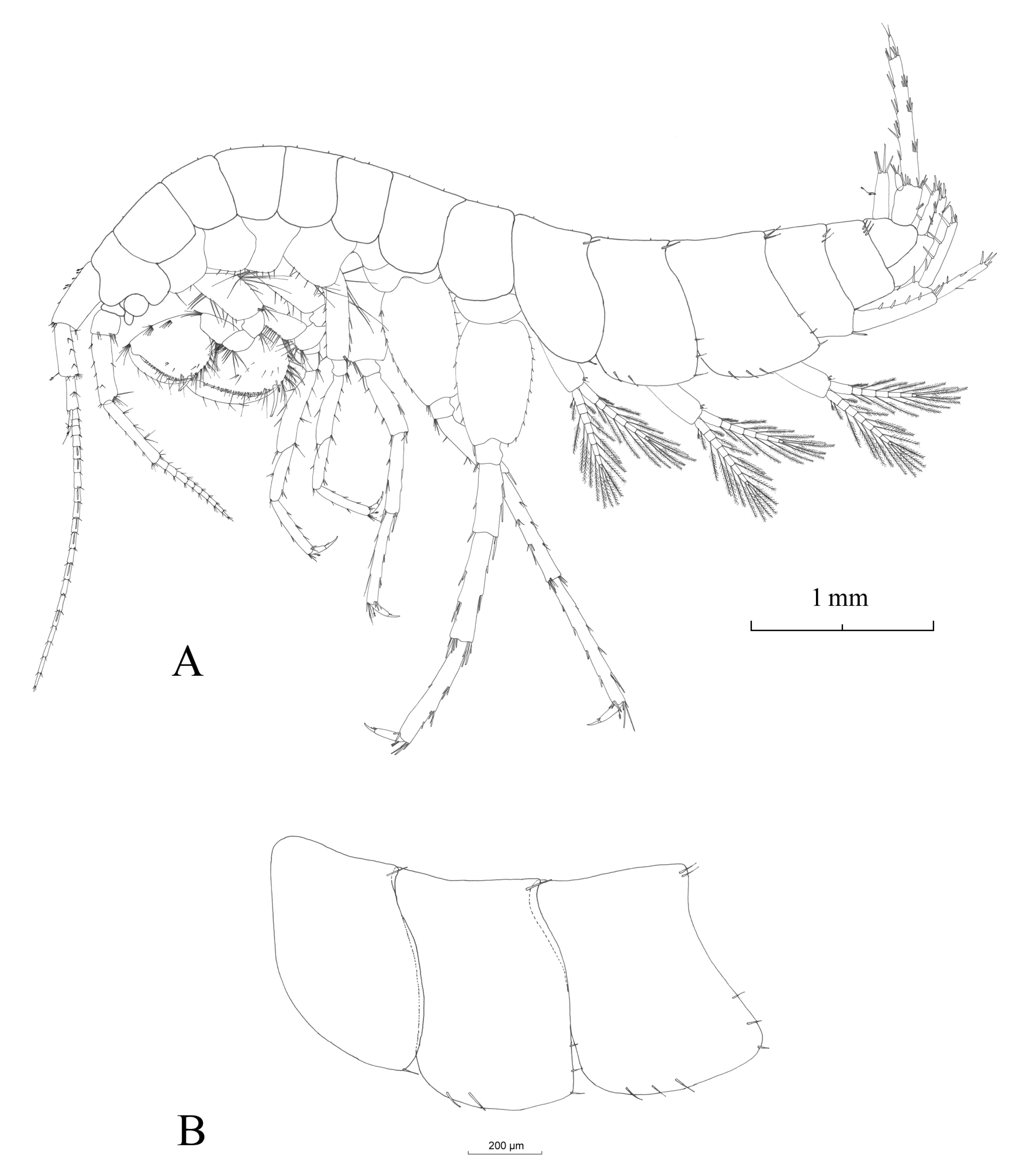
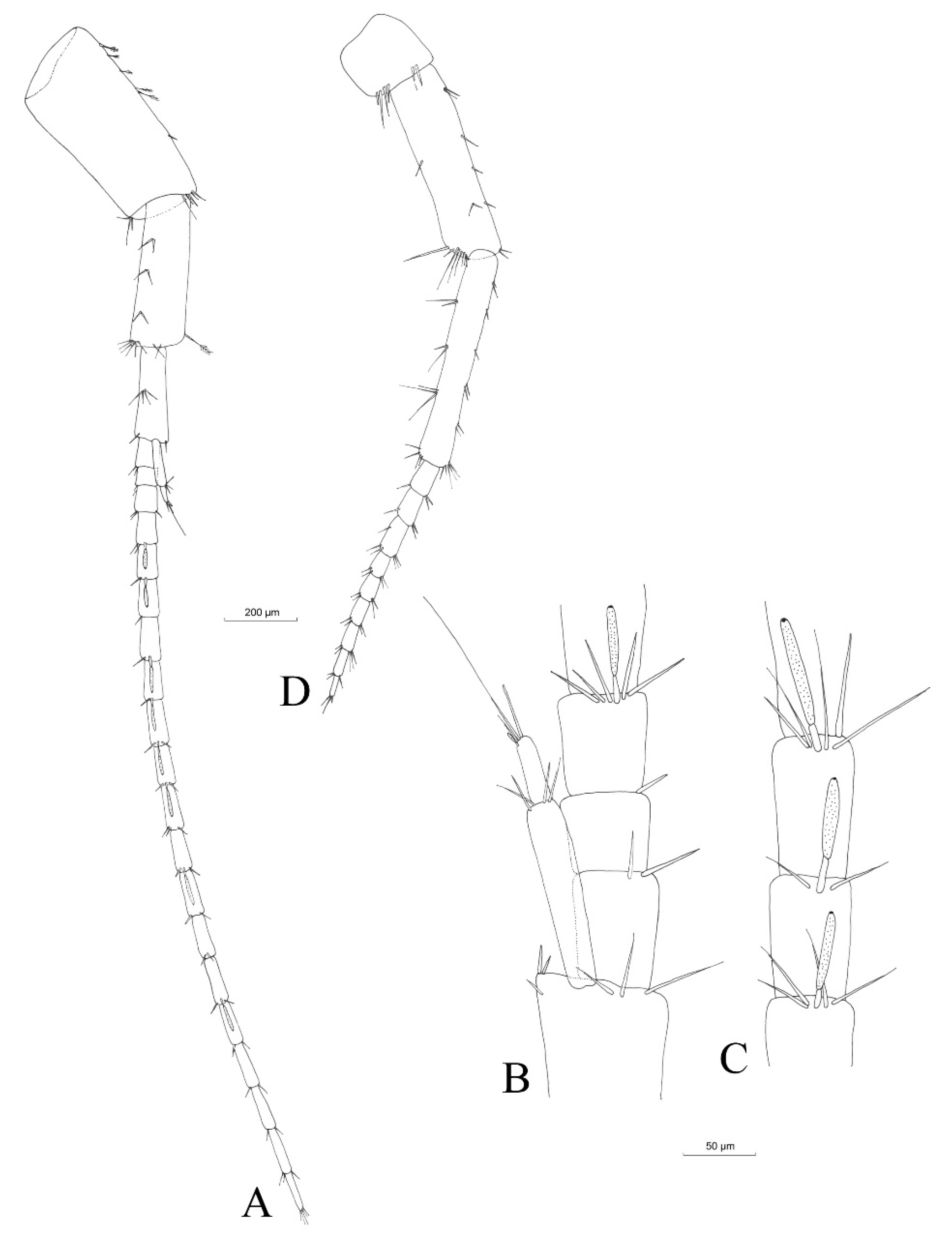
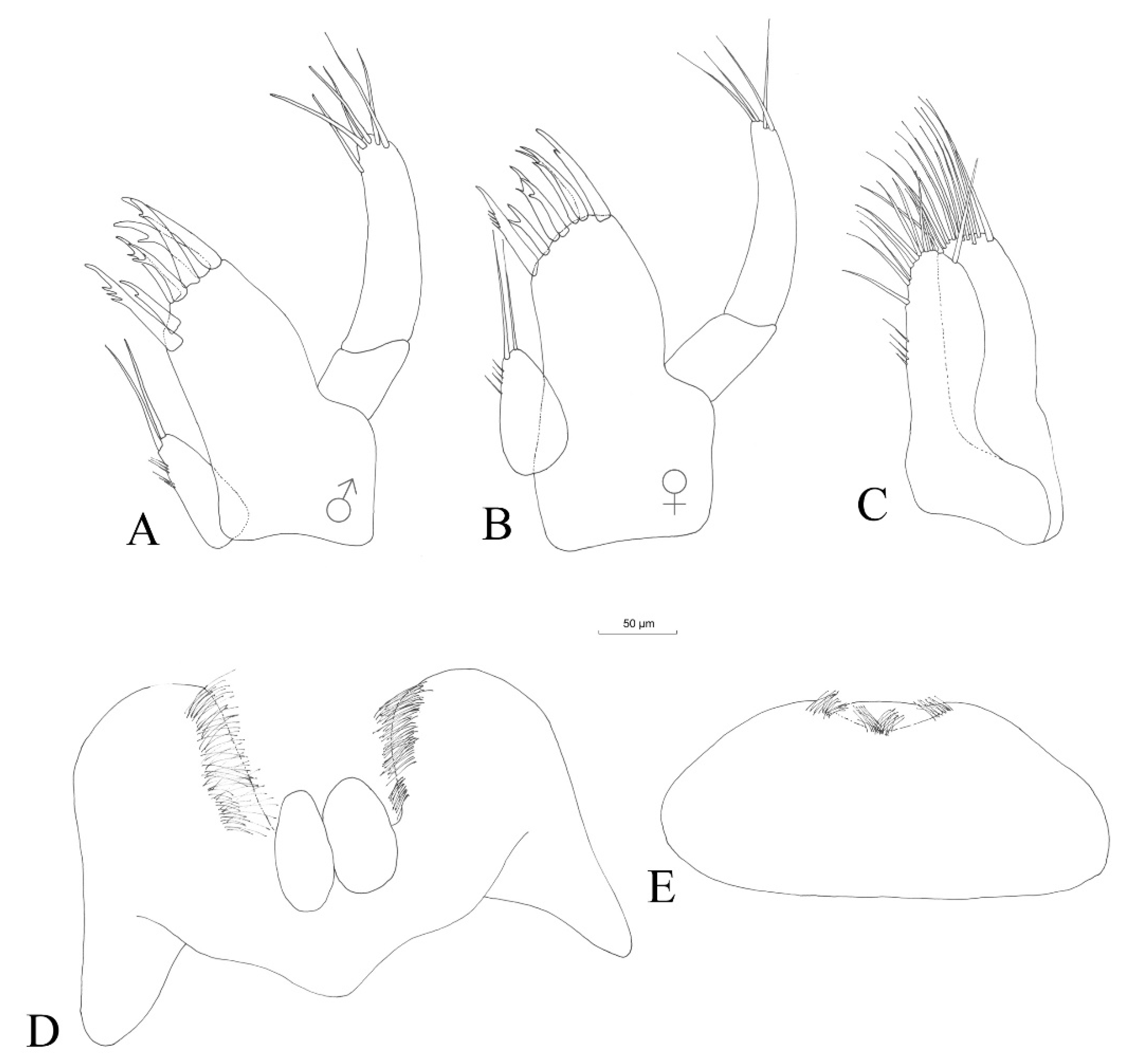
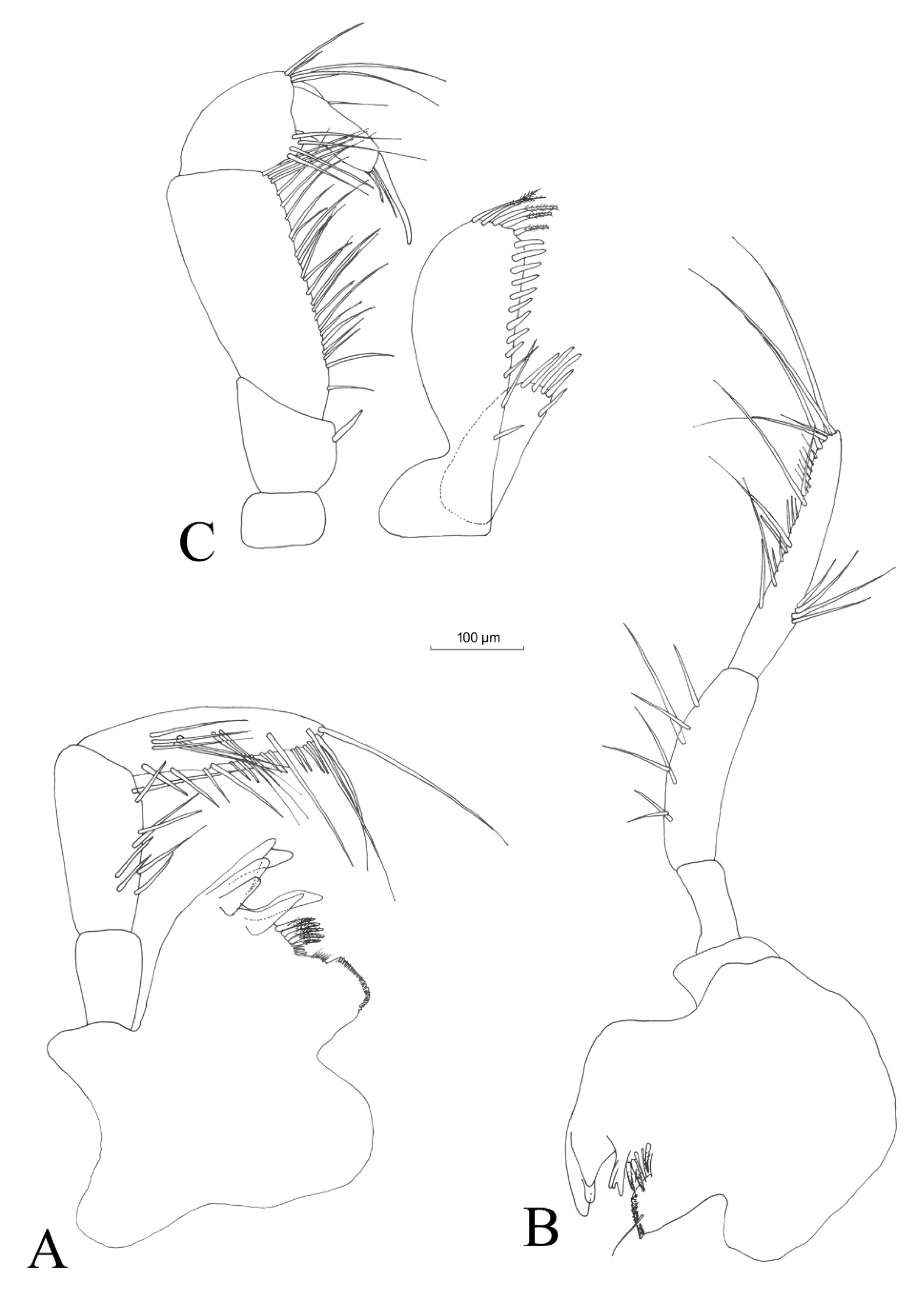
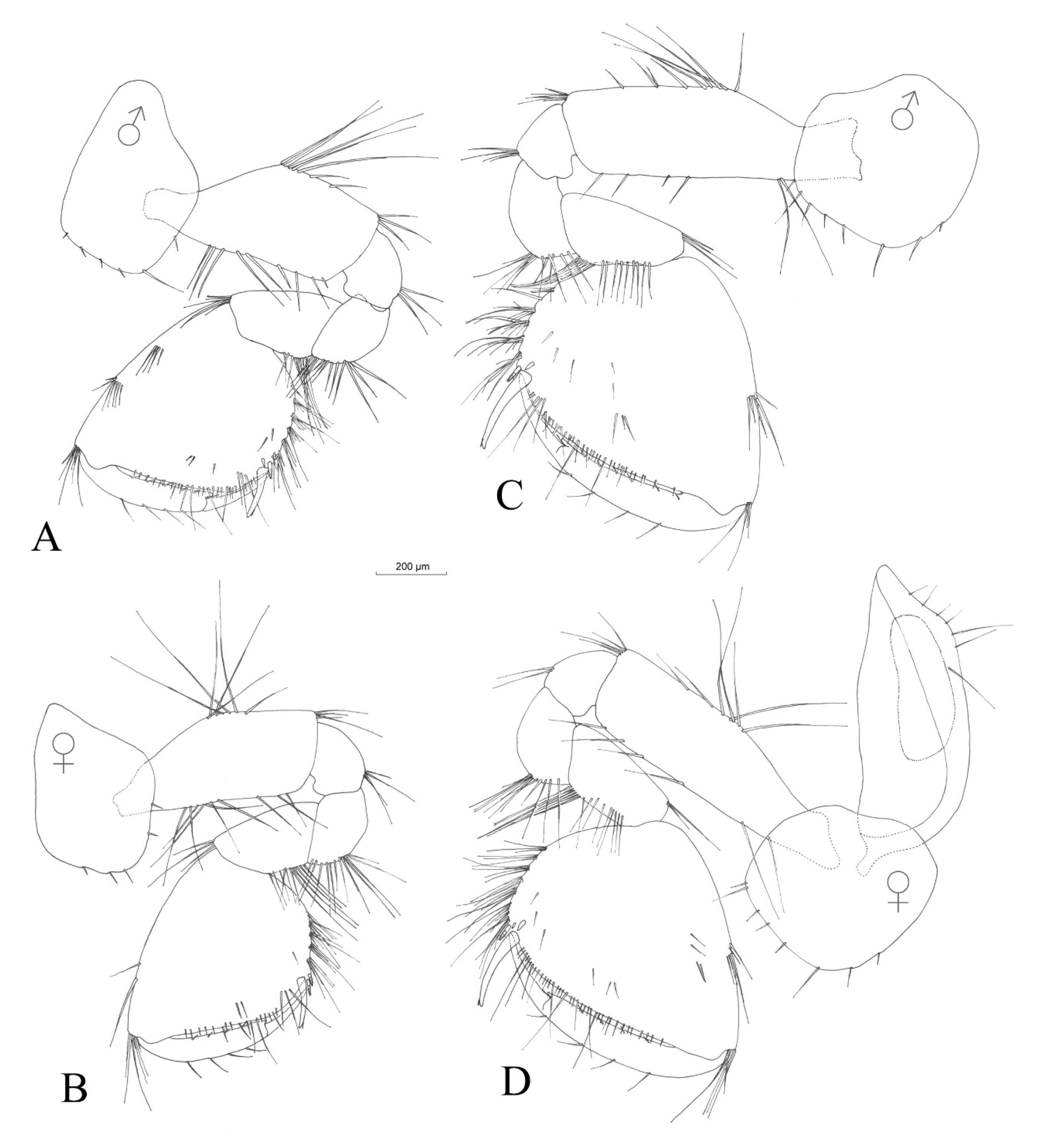
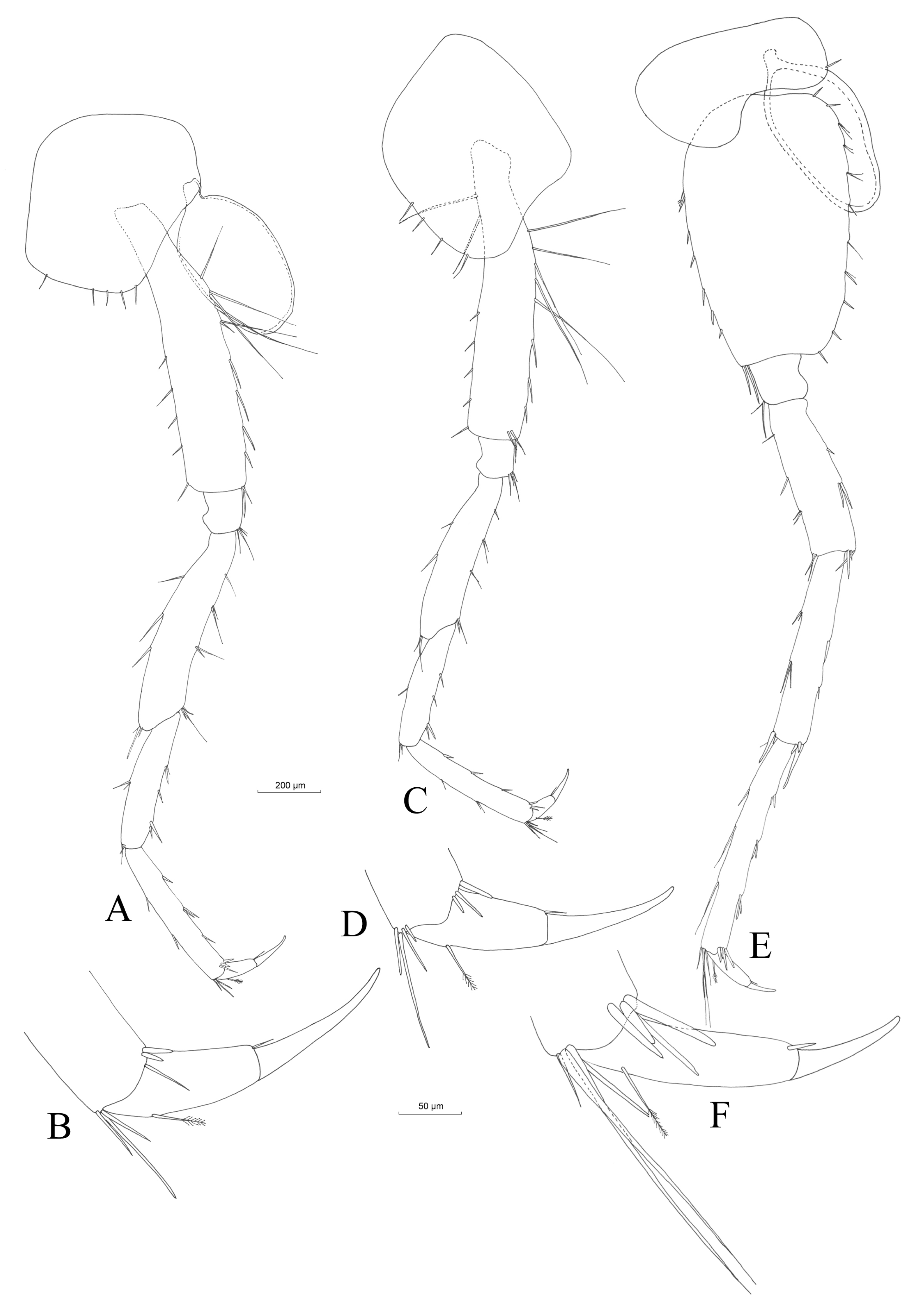
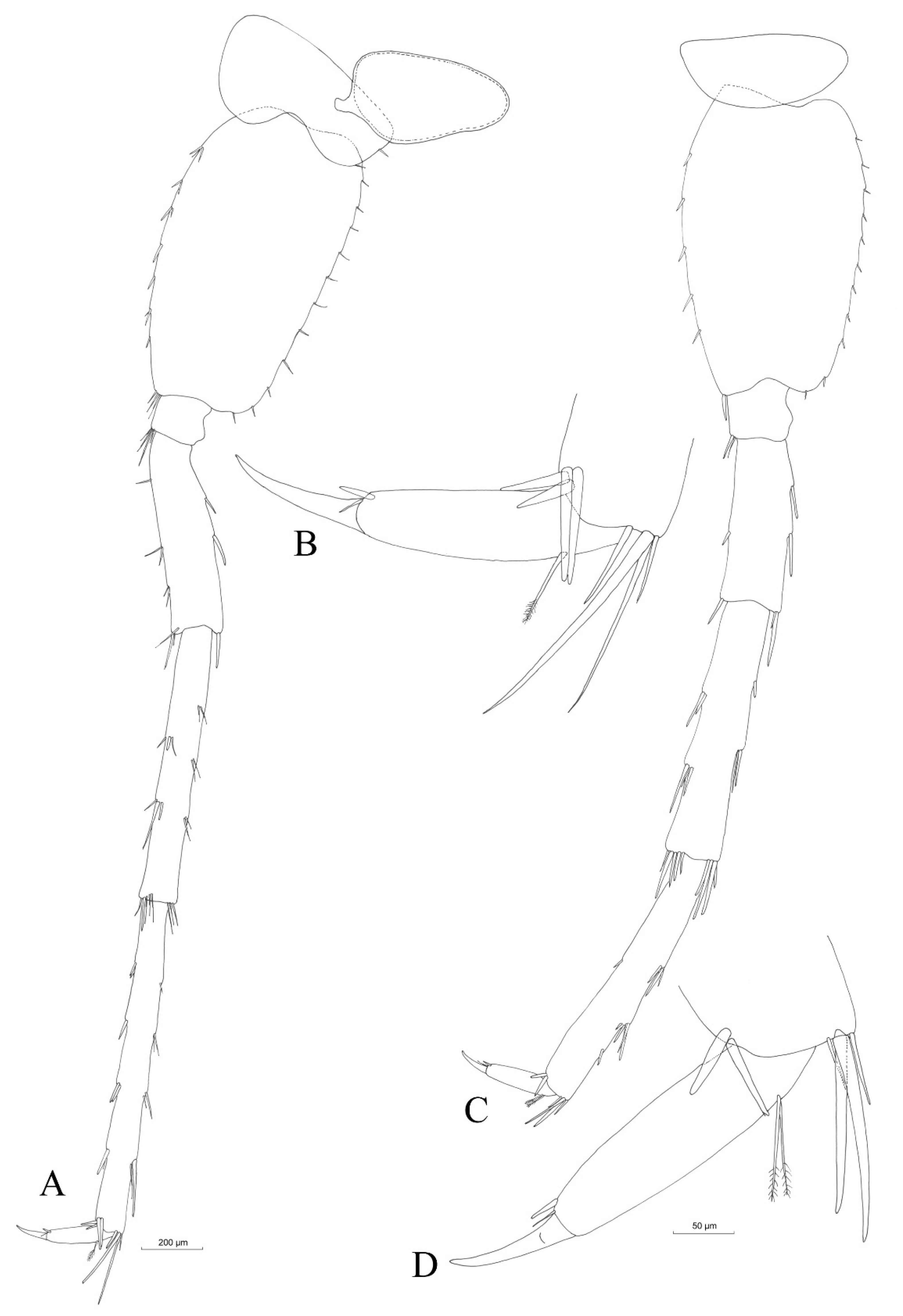
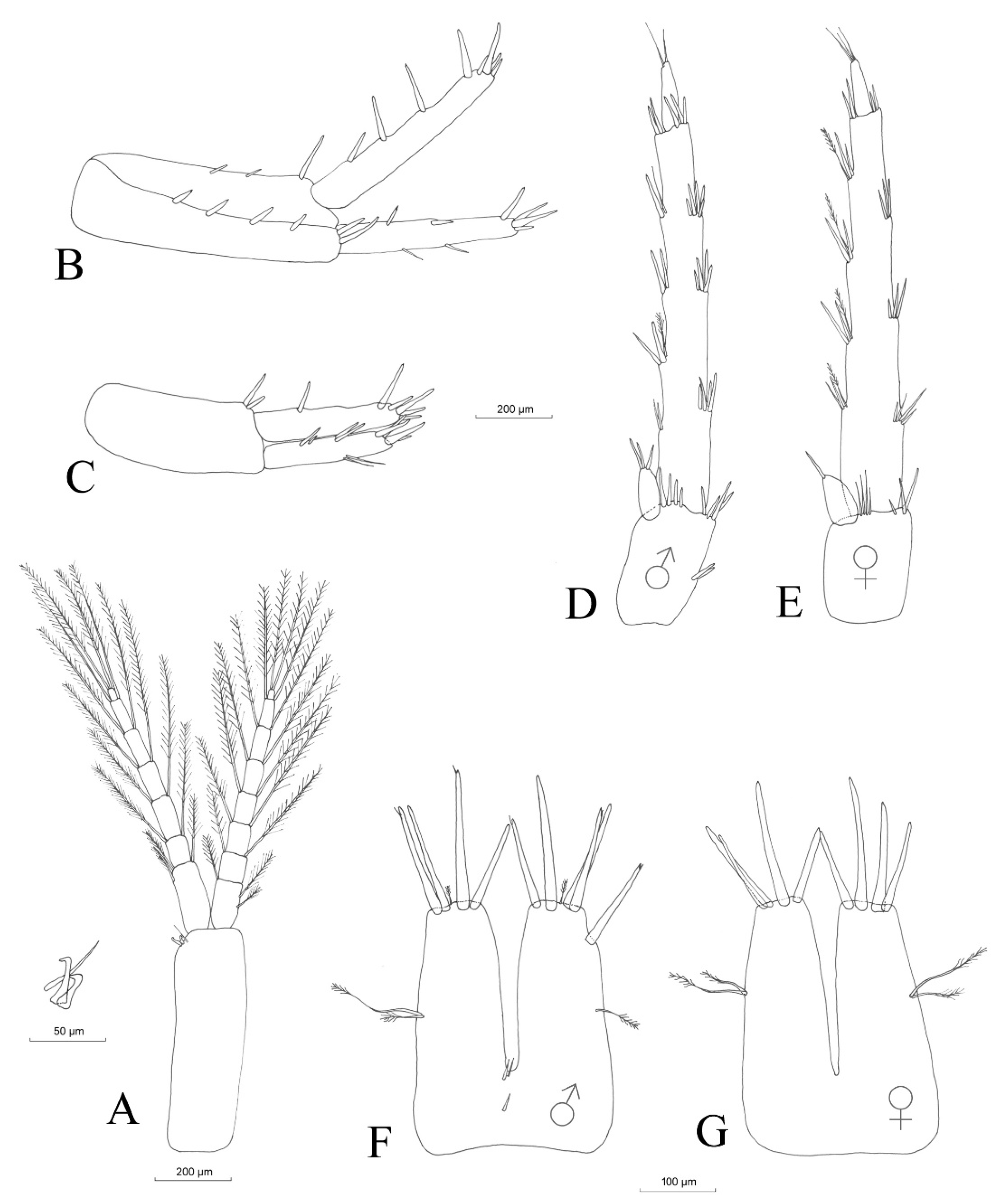
3.2.4. Description
Material Examined
Description (Male)
Mouthparts
3.2.5. Sexual Dimorphism
4. Discussion
4.1. New Species
4.1.1. Position of N. carolinensis sp. nov. in the Niphargid Tree of Life
4.1.2. Morphological Difference between Niphargus carolinensis sp. nov. and Species in Nearby Vicinity
4.2. Ecological Data
4.3. Endemic Species in the North

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Distribution | Literature |
|---|---|---|
| Niphargus carolinensis | 0.06 | This study |
| Niphargus enslini | 82 | [19] |
| Niphargus fontanus | 1150 | Weber and Schmitt in prep. |
| Niphargus puteanus | 756 | [20] |
| Niphargus schellenbergi | 551 | [15] |
| Niphargus tonywhitteni | 400 | [56] |
| Niphargus virei A | 420, 640 | [15,57] |
| Niphargus virei C | 93 | [15,57] |
4.4. Low Intraspecific Variability
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horton, T.; Lowry, J.; De Broyer, C.; Bellan-Santini, D.; Copilaș-Ciocianu, D.; Corbari, L.; Costello, M.J.; Daneliya, M.; Dauvin, J.-C.; Fišer, C.; et al. World Amphipoda Database. 2023. Available online: https://www.marinespecies.org/amphipoda (accessed on 7 September 2023).
- Karaman, G.S. New Data on Two Subterranean Species of the Family Niphargidae from Spain, Niphargus gallicus Schell., 1935 and N. delamarei Ruffo, 1954 (Contribution to the Knowledge of the Amphipoda 282. Contrib. Sect. Nat. Math. Biotech. Sci. 2017, 36, 105–120. [Google Scholar] [CrossRef]
- Esmaeili-Rineh, S.; Sari, A.; Fišer, C.; Bargrizaneh, Z. Completion of Molecular Taxonomy: Description of Four Amphipod Species (Crustacea: Amphipoda: Niphargidae) from Iran and Release of Database for Morphological Taxonomy. Zool. Anz. 2017, 271, 57–79. [Google Scholar] [CrossRef]
- Stevens, G.C. The Latitudinal Gradient in Geographical Range: How so Many Species Coexist in the Tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Hartke, T.R.; Fišer, C.; Hohagen, J.; Kleber, S.; Hartmann, R.; Koenemann, S. Morphological and Molecular Analyses of Closely Related Species in the Stygobiontic Genus Niphargus (Amphipoda). J. Crustac. Biol. 2011, 31, 701–709. [Google Scholar] [CrossRef][Green Version]
- Karaman, S. Beitrag Zur Kenntnis der Süsswasser-Amphipoden. Prirodosl. Razpr. 1932, 2, 179–232. [Google Scholar]
- Schellenberg, A. Krebstiere Oder Crustacea. In IV: Flohkrebse oder Amphipoda; die Tierwelt Deutschlands und der angrenzenden Meeresteile nach ihren Merkmalen und nach ihrer Lebenweise; Gustav Fischer: Jena, Germany, 1942. [Google Scholar]
- Schellenberg, A. Bemerkungen zu meinem Niphargus-Schlüssel und zur Verbreitung und Variabilität der Arten, Nebst Beschreibung Neuer Niphargus-Formen. Mitteilungen Aus Dem Zool. Mus. Berl. 1937, 22, 1–30. [Google Scholar] [CrossRef]
- Schellenberg, A. Die Höhere Krebsfauna im Süsswasser Deutschlands, ihre Zusammensetzung und ihr Artenzuwachs. Arch. Für Hydrobiol. 1937, 31, 229–241. [Google Scholar]
- Schellenberg, A. Schlüssel der Amphipodengattung Niphargus mit Fundortangaben und Mehreren Neuen Formen. Zool. Anz. 1935, 111, 204–211. [Google Scholar]
- Schellenberg, A. Der Niphargus Des Thüringer Waldes und Die Glazialreliktenfrage. Arch. Für Hydrobiol. 1935, 29, 274–281. [Google Scholar]
- Schellenberg, A. Weitere Deutsche und Ausländische Niphargiden. Zool. Anz. 1933, 102, 22–33. [Google Scholar]
- Schellenberg, A. Vier Blinde Amphipodenarten in einem Brunnen Oberbayerns. Zool. Anz. 1932, 98, 131–139. [Google Scholar]
- Weber, D. Grundwasserkrebse Niphargus und Microniphargus Im Harz an ihrer Nördlichen Verbreitungsgrenze—Ergebnisse Phylogenetischer Untersuchungen. Abh. Zur Karst-Und Höhlenkunde 2022, 40, 213–224. [Google Scholar]
- Weber, D. Molecular Phylogeny and Systematics of Central and Western European Niphargids (Crustacea: Amphipoda); Université liebre de Bruxelles: Bruxelles, Belgium, 2022. [Google Scholar]
- Weber, D. Die Niphargen (Malacostraca: Amphipoda: Niphargidae) der Schwäbischen Alb—Ergebnisse Phylogenetischer Studien. Laichinger Höhlenfreund 2022, 57, 25–35. [Google Scholar]
- Weber, D. Die Höhlenfauna und -Flora des Höhlenkatastergebietes Rheinland-Pfalz/Saarland, 4. Teil. Abhandlungen zur Karst- und Höhlenkunde; Verband der deutschen Höhlen- und Karstforscher e.V.: München, Germany, 2001; Volume 33, p. 1088. [Google Scholar]
- Weber, D. Die Evertebartenfauna der Höhlen und Künstlichen Hohlräume des Katastergebietes Westfalen Einschließlich der Quellen- und Grundwasserfauna. Abhandlungen zur Karst- und Höhlenkunde; Verband der deutschen Höhlen- und Karstforscher e.V.: München, Germany, 1991; Volume 25, p. 701. [Google Scholar]
- Weber, D.; Brad, T.; Stoch, F.; Flot, J.-F. Rediscovery and Redescription of Niphargus enslini Karaman, 1932 (Amphipoda, Niphargidae) in Southern Germany. Subterr. Biol. 2021, 40, 65–89. [Google Scholar] [CrossRef]
- Weber, D.; Flot, J.-F.; Weigand, H.; Weigand, A.M. Demographic History, Range Size and Habitat Preferences of the Groundwater Amphipod Niphargus puteanus (C.L. Koch in Panzer, 1836). Limnologica 2020, 82, 125765. [Google Scholar] [CrossRef]
- Weber, D.; Flot, J.-F. Rote Liste und Gesamtartenliste der Grundwasserkrebse (Niphargidae) des Saarlandes. In Rote Liste Gefährdeter Tiere und Pflanzen des Saarlandes, pdf-Ausgabe; Saarbrücken und Landsweiler-Reden: Schiffweiler, Germany, 2020; pp. 3–9. [Google Scholar]
- Weber, D.; Weigand, A.M. Groundwater Amphipods of the Hyporheic Interstitial: A Case Study from Luxembourg and The Greater Region. Diversity 2023, 15, 411. [Google Scholar] [CrossRef]
- Schminke, H.K. Amphipoda (Flohkrebse). In Grundwasserfauna Deutschlands. Ein Bestimmungswerk; DWA Deutsche Vereinigung für Wasserwirtschaft, Abwasser und Abfall e.V.: Hennef, Germany, 2007; pp. 239–272. [Google Scholar]
- Pust, J. Untersuchungen Zur Systematik, Morphologie und Ökologie der in Westfälischen Höhlen Vorkommenden Aquatischen Höhlentiere; Abhandlungen aus dem Westfälischen Museum für Naturkunde; Westfälisches Museum für Naturkunde: Münster, Germany, 1990; Volume 4. [Google Scholar]
- Ginet, R. Bilan Systematique Du Genre Niphargus En France; Université Claude-Bernard Lyon I: Lyon, France, 1995. [Google Scholar]
- Binczyk, K. Kupfererzbergbau Im Rahmedetal. De Rammuthe 2008, 2008, 16–20. [Google Scholar]
- Chappuis, P.A. Eine Neue Methode zur Untersuchung der Grundwasserfauna. Acta Sci. Math. Et Nat./Univ. Fr. Joseephina 1942, 6, 3–7. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Astrin, J.J.; Stüben, P.E. Phylogeny in Cryptic Weevils: Molecules, Morphology and New Genera of Western Palaearctic Cryptorhynchinae (Coleoptera: Curculionidae). Invertebr. Syst. 2008, 22, 503–522. [Google Scholar] [CrossRef]
- Verovnik, R.; Sket, B.; Trontelj, P. The Colonization of Europe by the Freshwater Crustacean Asellus aquaticus (Crustacea: Iso-poda) Proceeded from Ancient Refugia and Was Directed by Habitat Connectivity. Mol. Ecol. 2005, 14, 4355–4369. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.-F.; Wörheide, G.; Dattagupta, S. Unsuspected Diversity of Niphargus Amphipods in the Chemoautotrophic Cave Ecosystem of Frasassi, Central Italy. BMC Evol. Biol. 2010, 10, 171. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Villesen, P. FaBox: An Online Toolbox for Fasta Sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Statist. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Fišer, C.; Trontelj, P.; Luštrik, R.; Sket, B. Towards a Unified Taxonomy of Niphargus (Crustacea: Amphipoda): A Review of Morphological Variability. Zootaxa 2009, 2061, 1–22. [Google Scholar] [CrossRef]
- Weber, D.; Brad, T.; Weigand, A.; Flot, J.-F. Water Diviners Multiplied: Cryptic Diversity in the Niphargus aquilex Species Complex in Northern Europe. Biorxiv Prepr. Zool. 2023. [Google Scholar] [CrossRef]
- Spence Bate, C. On the Genus Niphargus (Schiödte). Nat. Hist. Rev. Q. J. Sci. 1859, 6, 163–166. [Google Scholar]
- Schellenberg, A. Subterrane Amphipoden Badens, nebst einem Neuen Niphargus Aus Polen. Zool. Anz. 1936, 113, 67–73. [Google Scholar]
- Karaman, G.S. Contribution to the Knowledge of the Amphipoda 150. One New Species of Genus Niphargus (Gammaridea, Niphargidae) from France, Niphargus renei n.sp. Ann. De Limnol. 1986, 22, 17–25. [Google Scholar] [CrossRef]
- Alther, R.; Bongni, N.; Borko, Š.; Fišer, C.; Altermatt, F. Citizen Science Approach Reveals Groundwater Fauna in Switzerland and a New Species of Niphargus (Amphipoda, Niphargidae). Subterr. Biol. 2021, 39, 1–31. [Google Scholar] [CrossRef]
- Fišer, C.; Alther, R.; Zakšek, V.; Borko, Š.; Fuchs, A.; Altermatt, F. Translating Niphargus Barcodes from Switzerland into Taxonomy with a Description of Two New Species (Amphipoda, Niphargidae). ZooKeys 2018, 760, 113–141. [Google Scholar] [CrossRef]
- Zagmajster, M.; Eme, D.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.-F.; Malard, F. Geographic Variation in Range Size and Beta Diversity of Groundwater Crustaceans: Insights from Habitats with Low Thermal Seasonality: Range Size and Beta Diversity in Non-Seasonal Habitats. Glob. Ecol. Biogeogr. 2014, 23, 1135–1145. [Google Scholar] [CrossRef]
- Fišer, C.; Mavrič, B.; Pekolj, A.; Zagmajster, M. Checklist of Amphipod Crustaceans (Crustacea: Amphipoda) in Slovenia. Nat. Slov. 2021, 23, 5–24. [Google Scholar]
- Rapoport, E.H. Areografía. In Estrategias Geográficas de Las Especies; Fondo de Cultura Económica: Mexico City, Mexico, 1975. [Google Scholar]
- Fišer, C.; Sket, B.; Stoch, F. Distribution of Four Narrowly Endemic Niphargus Species (Crustacea: Amphipoda) in the Western Dinaric Region with Description of a New Species. Zool. Anz. 2006, 245, 77–94. [Google Scholar] [CrossRef]
- Fišer, C.; Konec, M.; Alther, R.; Švara, V.; Altermatt, F. Taxonomic, Phylogenetic and Ecological Diversity of Niphargus (Amphipoda: Crustacea) in the Hölloch Cave System (Switzerland). Syst. Biodivers. 2017, 15, 218–237. [Google Scholar] [CrossRef]
- Borowsky, B. Responses to Light in Two Eyeless Cave Dwelling Amphipods (Niphargus ictus and Niphargus frasassianus). J. Crustac. Biol. 2011, 31, 613–616. [Google Scholar] [CrossRef]
- Angyal, D.; Balázs, G.; Zakšek, V.; Krízsik, V.; Fišer, C. Redescription of Two Subterranean Amphipods Niphargus molnari Méhely, 1927 and Niphargus gebhardti Schellenberg, 1934 (Amphipoda, Niphargidae) and their Phylogenetic Position. ZooKeys 2015, 509, 53–85. [Google Scholar] [CrossRef] [PubMed]
- Wichers, H.J. Sur Une Nouvelle Espèce d’Amphipode, Intermédiaire Entre Niphargus et Niphargellus, Du Boulonnais. Beaufortia 1964, 147, 185–198. [Google Scholar]
- McInerney, C.E.; Maurice, L.; Robertson, A.L.; Knight, L.R.F.D.; Arnscheidt, J.; Venditti, C.; Dooley, J.S.G.; Mathers, T.; Matthijs, S.; Erikkson, K.; et al. The Ancient Britons: Groundwater Fauna Survived Extreme Climate Changes over Tens of Millions of Years across NW Europe. Mol. Ecol. 2014, 23, 1153–1166. [Google Scholar] [CrossRef]
- Weber, D. Niphargus Tonywhitteni Fišer et al. 2018 Recorded for the First Time in Bavaria (Malacostraca, Amphipoda, Niphargidae); Spixiana: München, Germany, 2023. [Google Scholar]
- Weber, D.; Stoch, F.; Flot, J.-F. New Insight into the Phylogeny and Biogeography of the Subterranean Niphargus virei Clade (Amphipoda: Niphargidae) in Western Europe. In 3rd Central European Symposium for Aquatic Macroinvertebrate Research, Lodz; University of Lodz: Lodz, Poland, 2018. [Google Scholar]











| Inv.No. | Date | Pres. | Spec. | Isolate | COI | 28S | ITS2 |
|---|---|---|---|---|---|---|---|
| DW180512-003 | 12 May 2018 | ET96 | Weber | ULB | OR644127 | OR646283 | |
| DW180512-004 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-005 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-006 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-007 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-008 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-009 | 12 May 2018 | ET96 | Weber | ||||
| DW180512-010 | 12 May 2018 | ET96 | ISER | ||||
| DW180512-011 | 12 May 2018 | ET96 | ISER | SDEI | OR644128 | ||
| DW220205-001 * | 5 February 2022 | KGG | ISER | ||||
| DW220205-002 | 5 February 2022 | PG | -** | SDEI | OR644129 | OR711534 | |
| DW220205-003 | 5 February 2022 | ET96 | Weber | ||||
| DW220205-004 | 5 February 2022 | ET96 | Weber | ||||
| DW220906-001 | 6 September 2022 | PG | ISER | ||||
| DW220907-001 | 7 September 2022 | KGG | ISER | ||||
| DW220907-002 | 7 September 2022 | PG | ISER | ||||
| DW220907-003 | 7 September 2022 | PG | ISER | ||||
| DW220907-004 | 7 September 2022 | PG | ISER | ||||
| DW220907-005 | 7 September 2022 | ET96 | ISER | ||||
| DW220907-006 | 7 September 2022 | ET96 | ISER | ||||
| DW220907-007 | 7 September 2022 | ET96 | ISER |
| Primer | Nucleotide Sequence (5′ to 3′) | PCR | Seq | Ref. |
|---|---|---|---|---|
| LCO1490-JJ | CHA CWA AYC ATA AAG ATA TYG G | x | x | [29] |
| HCO2198-JJ | AWA CTT CVG GRT GVC CAA ARA ATC A | x | x | [29] |
| Niph15 | CAA GTA CCG TGA GGG AAA GTT | x | x | [30] |
| Niph16 | AGG GAA ACT TCG GAG GGA ACC | x | [30] | |
| Niph20 | AAA CAC GGG CCA AGG AGT AT | x | [31] | |
| Niph21 | TAT ACT CCT TGG CCC GTG TT | x | [31] | |
| ITSsf2 = Niph22 | AAGGCTATAGCTGGCGATCA | x | x | [31] |
| ITSsr2 = Niph23 | TCAGCGGGTAACCTCTCCTA | x | x | [31] |
| Appendage | mm | Appendage | mm | ||
|---|---|---|---|---|---|
| Body length | 8.93 | PIII | Coxal plate depth | 0.62 | |
| Head length | 0.66 | Coxal plate width | 0.49 | ||
| Antenna I | Peduncle length | 1.17 | Basipodite | 0.96 | |
| Flagellum length | 2.19 | Ischiopodite | 0.13 | ||
| Art 1 acc. flagellum | 0.13 | Meropodite | 0.67 | ||
| Antenna II | Peduncle length | 1.32 | Carpopodite | 0.43 | |
| Flagellum length | 0.70 | Propodite | 0.49 | ||
| Mandible palp | Art 1 | 0.10 | Dactylopodite 1 | 0.11 | |
| Art 2 | 0.20 | Dactylopodite 2 | 0.13 | ||
| Art 3 | 0.26 | PIV | Coxal plate depth | 0.54 | |
| Gnathopod I | Propodite length | 0.57 | Coxal plate width | 0.55 | |
| Propodite width | 0.53 | Basipodite | 0.95 | ||
| Propodite depth | 0.40 | Ischiopodite | 0.13 | ||
| Dactylopodite 1 | 0.38 | Meropodite | 0.55 | ||
| Dactylopodite 2 | 0.20 | Carpopodite | 0.41 | ||
| Carpopodite length | 0.30 | Propodite | 0.43 | ||
| Basipodite width | 0.28 | Dactylopodite 1 | 0.08 | ||
| Basipodite length | 0.66 | Dactylopodite 2 | 0.10 | ||
| Coxal plate depth | 0.51 | PV | Basipodite length | 0.83 | |
| Coxal plate width | 0.36 | Basipodite width | 0.53 | ||
| Gnathopod II | Propodite length | 0.74 | Ischiopodite | 0.15 | |
| Propodite width | 0.70 | Meropodite | 0.50 | ||
| Propodite depth | 0.47 | Carpopodite | 0.63 | ||
| Dactylopodite 1 | 0.56 | Propodite | 0.71 | ||
| Dactylopodite 2 | 0.24 | Dactylopodite 1 | 0.15 | ||
| Carpopodite length | 0.37 | Dactylopodite 2 | 0.08 | ||
| Basipodite width | 0.26 | PVI | Basipodite length | 0.97 | |
| Basipodite length | 0.82 | Basipodite width | 0.57 | ||
| Coxal plate depth | 0.48 | Ischiopodite | 0.15 | ||
| Coxal plate width | 0.50 | Meropodite | 0.62 | ||
| Uropod I | Basipodite length | 0.70 | Carpopodite | 0.91 | |
| Endopodite length | 0.57 | Propodite | 1.10 | ||
| Exopodite length | 0.46 | Dactylopodite 1 | 0.20 | ||
| Uropod II | Basipodite length | 0.47 | Dactylopodite 2 | 0.10 | |
| Endopodite length | 0.38 | PVII | Basipodite length | 0.91 | |
| Exopodite length | 0.33 | Basipodite width | 0.59 | ||
| Uropod III | Basipodite length | 0.34 | Ischiopodite | 0.19 | |
| Endopodite length | 0.14 | Meropodite | 0.54 | ||
| Exopodite art 1 | 1.00 | Carpopodite | 0.87 | ||
| Exopodite art 2 | 0.18 | Propodite | 0.89 | ||
| Telson | Cleft length | 0.23 | Dactylopodite 1 | 0.21 | |
| Telson width | 0.26 | Dactylopodite 2 | 0.10 | ||
| Telson length | 0.33 | ||||
| Longest spine length | 0.19 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, D.; Brad, T. Niphargus carolinensis sp. nov. (Amphipoda: Niphargidae), an Endemic Species in the Northern Distribution Area of the Genus. Hydrobiology 2023, 2, 554-574. https://doi.org/10.3390/hydrobiology2040037
Weber D, Brad T. Niphargus carolinensis sp. nov. (Amphipoda: Niphargidae), an Endemic Species in the Northern Distribution Area of the Genus. Hydrobiology. 2023; 2(4):554-574. https://doi.org/10.3390/hydrobiology2040037
Chicago/Turabian StyleWeber, Dieter, and Traian Brad. 2023. "Niphargus carolinensis sp. nov. (Amphipoda: Niphargidae), an Endemic Species in the Northern Distribution Area of the Genus" Hydrobiology 2, no. 4: 554-574. https://doi.org/10.3390/hydrobiology2040037
APA StyleWeber, D., & Brad, T. (2023). Niphargus carolinensis sp. nov. (Amphipoda: Niphargidae), an Endemic Species in the Northern Distribution Area of the Genus. Hydrobiology, 2(4), 554-574. https://doi.org/10.3390/hydrobiology2040037