
Aquatic Macroinvertebrate Community Structure along a Continuum in a Spring Dominated River, Missouri, USA



Abstract
1. Introduction
2. Materials and Methods
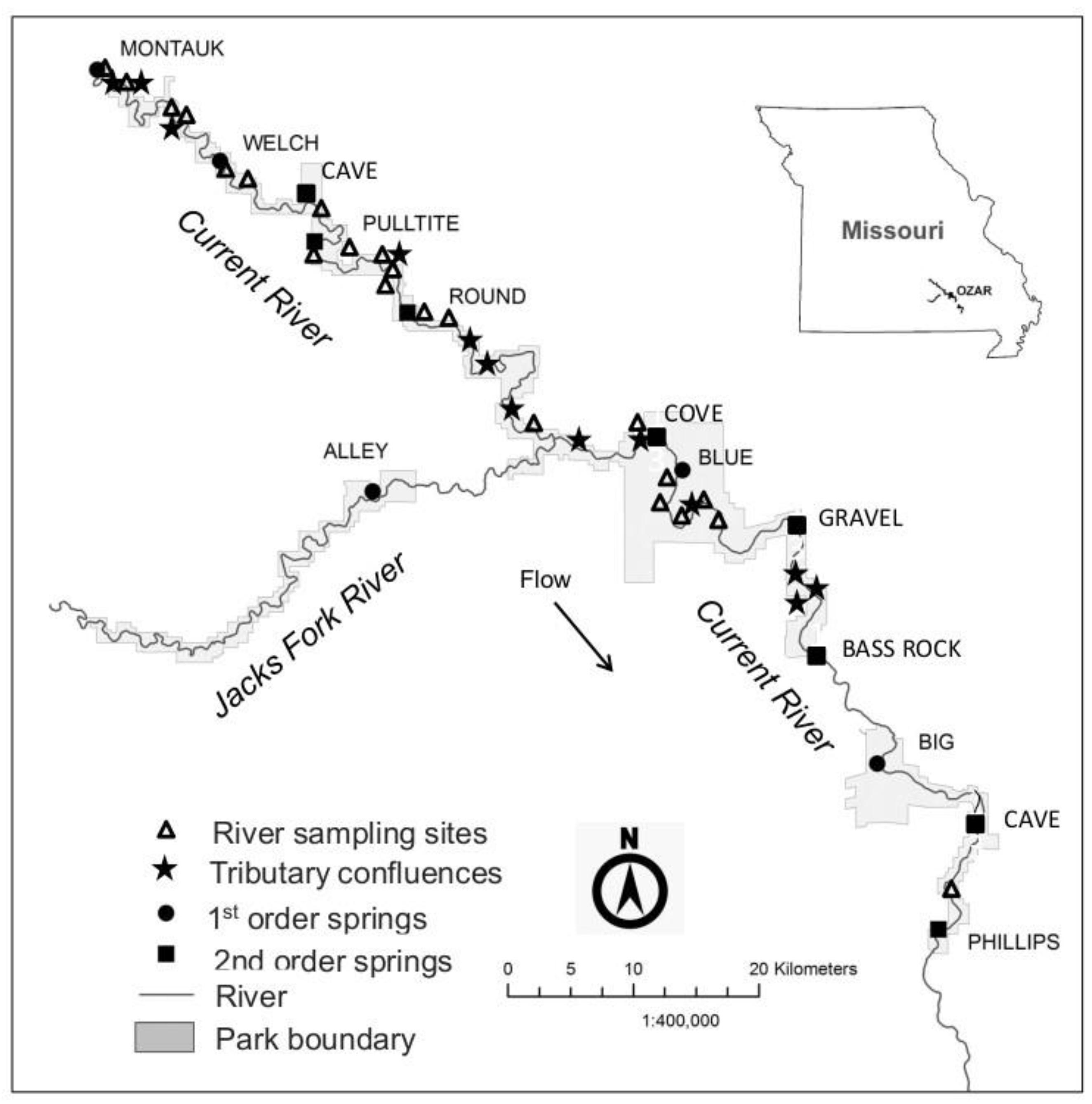
2.1. Study Area and Site Selection
2.2. Macroinvertebrate Sampling
2.3. Habitat and Water Quality
2.4. Statistical Analysis
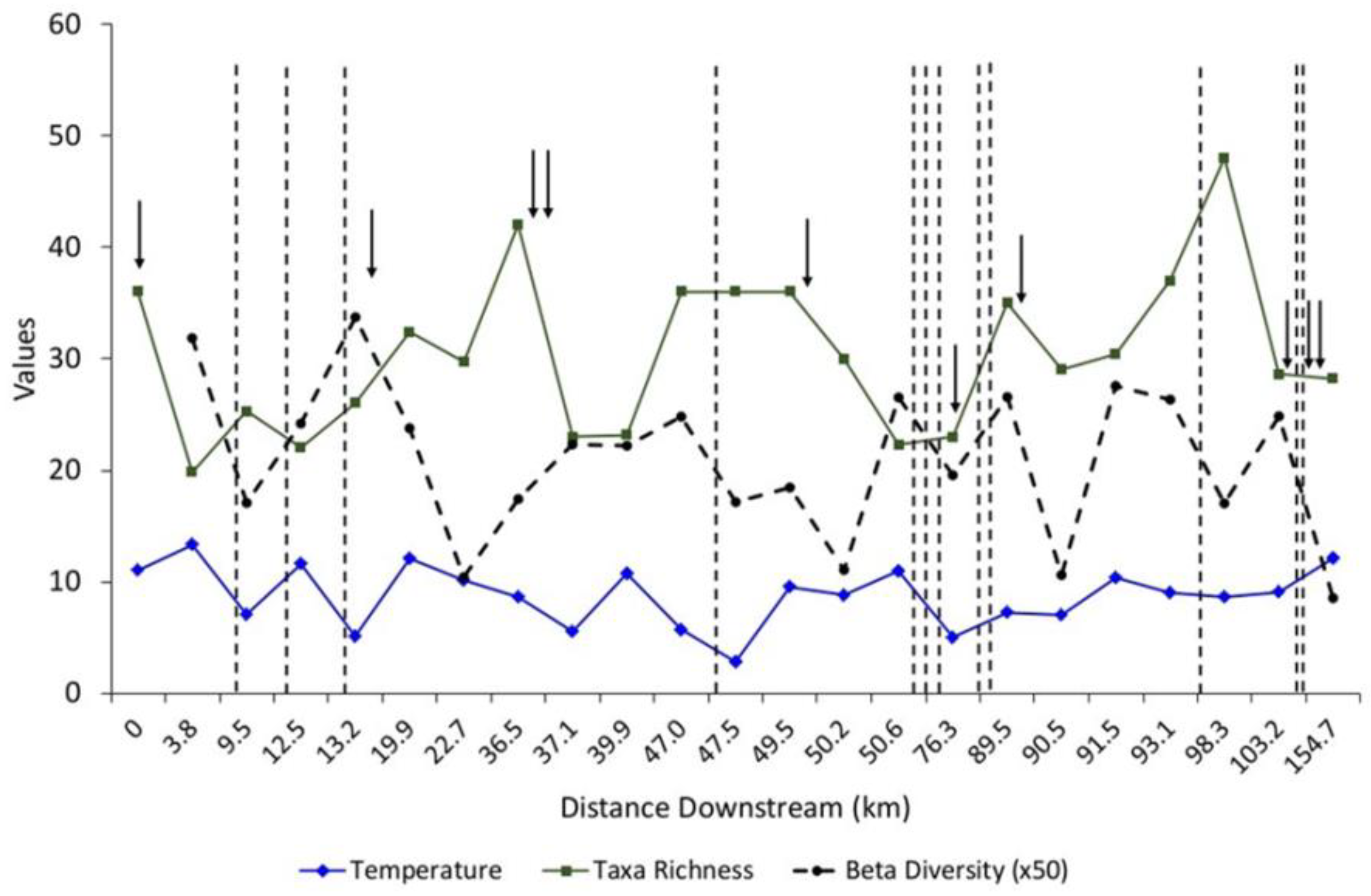
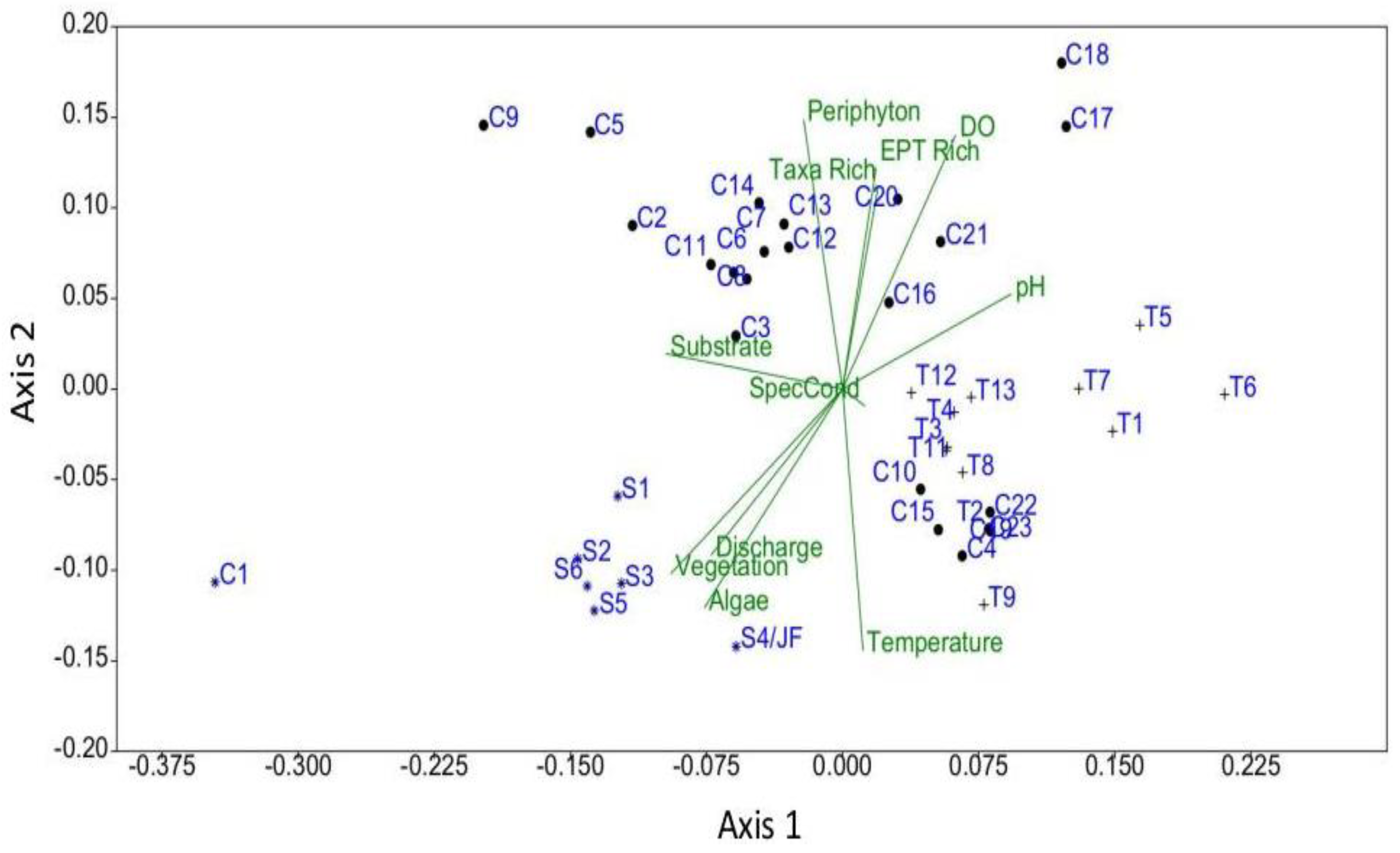
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Site Name | Site Number | Distance between Sites (km) | Springs | Tributaries | River |
|---|---|---|---|---|---|
| Montauk Spring/Current River | C1 | 0 | X | ||
| Current River at Tan Vat | C2 | 3.80 | X | ||
| Ashley Creek * | -- | 3.85 | X | ||
| Current River at Parker | C3 | 5.15 | X | ||
| Schafer Spring Creek | T2 | 0.11 | X | ||
| Current River at Cedar Grove | C4 | 2.92 | X | ||
| Big Creek (West) | T3 | 0.72 | X | ||
| Current River downstream of Big Creek | C5 | 0.30 | X | ||
| Welch Spring | S1 | 6.00 | X | ||
| Current River downstream of Welch Spring | C6 | 0.66 | X | ||
| Current River at Howell | C7 | 2.78 | X | ||
| Cave Spring * | -- | 6.50 | |||
| Current River upstream of Pulltite Spring | C8 | 7.30 | X | ||
| Pulltite Spring | S2 | 0.30 | X | ||
| Current River downstream of Pulltite Spring | C9 | 0.30 | X | ||
| Current River further downstream Pulltite Spring | C10 | 2.79 | X | ||
| Current River Upstream of Sinking Creek | C11 | 7.09 | X | ||
| Sinking Creek | T4 | 0.23 | X | ||
| Current River downstream of Sinking Creek | C12 | 0.28 | X | ||
| Current River upstream of Round Spring | C13 | 2.00 | X | ||
| Round Spring | S3 | 0.54 | X | ||
| Current River downstream of Round Spring | C14 | 0.20 | X | ||
| Current River further downstream of Round Spring | C15 | 0.43 | X | ||
| Big Creek (East) Creek | T5 | 11.25 | X | ||
| Sutton Creek | T6 | 9.90 | X | ||
| Thompson Creek | T7 | 3.90 | |||
| Current River downstream of Thompson Creek | C16 | 0.57 | X | X | |
| Alley Spring/Jacks Fork | S4/JF | 0,38 | X | ||
| Blair Creek | T8 | 7.44 | X | ||
| Powder Mill Creek | T9 | 3.54 | X | ||
| Current River upstream of Blue Spring | C17 | 1.92 | X | ||
| Blue Spring | S5 | 0.14 | X | ||
| Current River downstream of Blue Spring | C18 | 0.79 | X | ||
| Current River further downstream of Blue Spring | C19 | 1.00 | X | ||
| Current River upstream of Rocky Creek | C20 | 1.66 | X | ||
| Rocky Creek | T10 | 2.07 | X | ||
| Current River Downstream of Rocky Creek | C21 | 3.08 | X | ||
| Current River near Log Yard | C22 | 4.91 | X | ||
| Carr Creek | T11 | 1.20 | X | ||
| Gravel Spring * | -- | 10.19 | X | ||
| Rogers Creek | T12 | 9.50 | X | ||
| Mill Creek | T13 | 5.80 | X | ||
| Bass Rock Spring * | -- | 1.50 | |||
| Big Spring | S6 | 9.40 | X | ||
| Current River downstream of Big Spring | C23 | 13.94 | X |
References
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Heino, J.; Parviainen, J.; Paavola, R.; Jehle, M.; Louhi, P.; TMuotka, T. Characterizing macroinvertebrate assemblage structure in relation to stream size and tributary position. Hydrobiologia 2005, 539, 121–130. [Google Scholar] [CrossRef]
- Hellmann, J.K.; Erikson, J.S.; Queenborough, S.A. Evaluating macroinvertebrate community shifts in the confluence of freestone and limestone streams. J. Limnol. 2015, 74, 64–74. [Google Scholar] [CrossRef]
- Fleishman, E.; Noss, R.F.; Noon, B.R. Utility and limitations of species richness metrics for conservation planning. Ecol. Indic. 2006, 6, 543–553. [Google Scholar] [CrossRef]
- Heino, J.; Muotka, T.; Paavola, R. Determinants of macroinvertebrate diversity in headwater streams: Regional and local influences. J. Anim. Ecol. 2003, 72, 425–434. [Google Scholar] [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 280–338. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef]
- Astorga, A.; Death, R.; Paavola, R.; Chakraborty, M.; Muotka, T. Habitat heterogeneity drives the geographical distribution of beta diversity: The case of New Zealand stream invertebrates. Ecol. Evol. 2014, 4, 2693–2702. [Google Scholar] [CrossRef]
- Ward, J.V.; Stanford, J.A. Thermal responses in the evolutionary ecology of aquatic insects. Annu. Rev. Entomol. 1982, 27, 97–117. [Google Scholar] [CrossRef]
- Barquín, J.; Death, R.G. Spatial patterns of macroinvertebrate diversity in New Zealand springbrooks and rhithral streams. J. N. Am. Benthol. Soc. 2006, 25, 768–786. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Barquín, J.; Death, R.G. Downstream changes in spring-fed stream invertebrate communities: The effect of increased temperature range? J. Limnol. 2011, 70, 134–146. [Google Scholar] [CrossRef][Green Version]
- Ebersole, J.L.; Wigington, P.L., Jr.; Leibowitz, S.G.; Comeleo, R.L.; Van Sickle, J. Predicting the occurrence of cold-water patches at intermittent and ephemeral tributary confluences with warm rivers. Freshw. Sci. 2015, 34, 111–124. [Google Scholar] [CrossRef]
- Mattson, R.A.; Epler, J.H.; Hein, M.K. Description of benthic communities in karst, spring-fed streams of north central Florida. J. Kans. Entomol. Soc. 1995, 68, 18–41. [Google Scholar]
- Benda, L.; Andras, K.; Miller, D.; Bigelow, P. Confluence effects in rivers: Interactions of basin scale, network geometry, and disturbance regimes. Water Resour. Bull. 2004, 40, 1–15. [Google Scholar] [CrossRef]
- Benda, L.; Poff, N.L.; Miller, D.; Dunne, T.; Reeves, G.; Pess, G.; Pollock, M. The network dynamics hypothesis: How channel networks structure riverine habitats. BioScience 2004, 54, 413–427. [Google Scholar] [CrossRef]
- Scarsbrook, M.; Barquin, J.; Gray, D. New Zealand Coldwater Springs and Their Biodiversity. Science for Conservation 278; Science & Technology Publishing, Department of Conservation: Wellington, New Zealand, 2007; p. 72. [Google Scholar]
- Smith, H.; Wood, P.J.; Gunn, J. The influence of habitat structure and flow permanence on invertebrate communities in karst spring systems. Hydrobiologia 2003, 510, 53–66. [Google Scholar] [CrossRef]
- Smith, H.; Wood, P.J. Flow permanence and macroinvertebrate community variability in limestone spring systems. Hydrobiologia 2002, 487, 45–58. [Google Scholar] [CrossRef]
- Lusardi, R.A.; Bogan, M.T.; Moyle, P.B.; Dahlgren, R.A. Environment shapes invertebrate assemblage structure differences between volcanic spring-fed and runoff rivers in northern California. Freshw. Sci. 2016, 35, 1010–1022. [Google Scholar] [CrossRef]
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Vineyard, J.D.; Feder, G.L.; Pflieger, W.L.; Lipscomb, R.G. Springs of Missouri with Sections on Fauna and Flora; Water Resources Report No. 29; Missouri Geological Survey and Water Resources: Rolla, MO, USA, 1974; p. 212. [Google Scholar]
- Bowles, D.E.; Dodd, H.R. Floristics and community ecology of aquatic vegetation occurring in seven large springs at Ozark National Scenic Riverways, Missouri (U.S.A.), 2007–2012. J. Bot. Res. Inst. Tex. 2015, 9, 235–249. [Google Scholar]
- Meinzer, O.E. Large Springs in the United States; U.S. Geological Survey Water-Supply Paper 557; United States Geological Survey: Washington, DC, USA, 1927; p. 94. [Google Scholar]
- Mugel, D.N.; Richards, J.M.; Schumacher, J.G. Geohydrologic Investigations and Landscape Characteristics of Areas Contributing Water to Springs, the Current River, and Jacks Fork, Ozark National Scenic Riverways, Missouri; U.S. Geological Survey Scientific Investigations Report 2009–5138; United States Geological Survey: Washington, DC, USA, 2009; p. 80. [Google Scholar]
- Bowles, D.E.; Luraas, J.A.; Morrison, L.W.; Dodd, H.R.; Williams, M.H.; Rowell, G.A.; DeBacker, M.D.; Hinsey, J.A.; Usrey, F.D.; Haack, J.L. Protocol for Monitoring Aquatic Invertebrates at Ozark National Scenic Riverways, Missouri, and Buffalo National River, Arkansas; Natural Resource Report NPS/HTLN/NRR—2007/00; US National Park Service: Fort Collins, CO, USA, 2007; p. 138. [Google Scholar]
- Moulton, S.R., II; Kennen, J.G.; Goldstein, R.M.; Hambrook, J.A. Revised Protocols for Sampling Algal, Invertebrate, and Fish Communities as Part of the National Water-Quality Assessment Program. Open-File Report 02-150; US Geological Survey: Reston, VT, USA, 2022; p. 75. [Google Scholar]
- Heth, R.L. Diversity of Macroinvertebrates in Tributaries of the Jacks Fork and Current Rivers, Ozark National Scenic Riverways, Missouri and Efficacy of Spring-Fed Tributaries as Refugia. Ph.D. Dissertation, University of Missouri-Columbia, Columbia, MO, USA, 2015; p. 223. [Google Scholar]
- Heth, R.L.; Bowles, D.E.; Havel, J.E. Potential impacts of stream crossing traffic on macroinvertebrate communities in a Missouri Ozark River. River Res. Appl. 2016, 32, 925–934. [Google Scholar] [CrossRef]
- Vinson, M.R.; Hawkins, C.P. Effects of sampling area and subsampling procedure on comparisons of taxa richness among streams. J. N. Am. Benthol. Soc. 1996, 15, 392–399. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Carter, R.W.; Davidian, J. General Procedure for Gaging Streams. Book 3, Chapter A6 of Techniques of Water-Resources Investigations of the United States Geological Survey; United States Government Printing Office: Washington, DC, USA, 1966; p. 13. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Wilson, M.V.; Shmida, A. Measuring beta diversity with presence-absence data. J. Ecol. 1984, 72, 1055–1064. [Google Scholar] [CrossRef]
- Bowles, D.E.; Morrison, L.W.; Cribbs, J.T. Aquatic invertebrate community structure, biological condition, habitat, and water-quality at Ozark National Scenic Riverways, Missouri, 2005–2014, Missouri, 2005–2014. J. Ark. Acad. Sci. 2018, 72, 67–80. [Google Scholar]
- Bowles, D.E.; Dodd, H.R.; Hinsey, J.A.; Cribbs, J.T.; Luraas, J.A. Spring Communities Monitoring at Ozark National Scenic Riverways, Missouri: 2007–2009 Status Report; Natural Resource Technical Report NPS/OZAR/NRTR—2011/511; National Park Service: Fort Collins, CO, USA, 2011; p. 41. [Google Scholar]
- Bowles, D.E.; Dodd, H.R.; Hinsey, J.A.; Cribbs, J.T.; Williams, J.M. Aquatic Invertebrate Community Structure in Springs at Ozark National Scenic Riverways, Missouri, 2007–2016; Natural Resource Data Series NPS/HTLN/NRDS—2018/1161; National Park Service: Fort Collins, CO, USA, 2018; p. 37. [Google Scholar]
- Buria, L.; Albarino, R.; Villanueva, V.D.; Modenutti, B.; Balseiro, E. Impact of exotic rainbow trout on the benthic macroinvertebrate community from Andean-Patagonian headwater streams. Fundam. Appl. Limnol. Arch. Fur Hydrobiol. 2007, 168, 145–154. [Google Scholar] [CrossRef]
- Minshall, G.W.; Cummins, K.W.; Peterson, R.C.; Cushing, C.E.; Bruns, D.A.; Sedell, J.R.; Vannote, R.L. Developments in stream ecosystem theory. Can. J. Fish. Aquat. Sci. 1985, 42, 1045–1055. [Google Scholar] [CrossRef]
- Ward, J.V.; Stanford, J.A. The serial discontinuity concept of lotic ecosystems. In Dynamics of Lotic Ecosystems; Fontaine, T.D., Bartell, S.M., Eds.; Ann Arbor Scientific Publishers: Ann Arbor, MI, USA, 1983; pp. 29–42. [Google Scholar]
- Stanford, J.A.; Ward, J.V. Revisiting the serial discontinuity concept. River Res. Appl. 2001, 17, 303–310. [Google Scholar] [CrossRef]
- Svendsen, K.M.; Renshaw, C.E.; Magilligan, F.J.; Nislow, K.H.; Kaste, J.M. Flow and sediment regimes at tributary junctions on a regulated river: Impact on sediment residence time and benthic macroinvertebrate communities. Hydrol. Process. 2009, 23, 284–296. [Google Scholar] [CrossRef]
- Westhoff, J.T.; Paukert, C.P. Climate change simulations predict altered biotic response in a thermally heterogeneous stream system. PLoS ONE 2014, 9, e111438. [Google Scholar] [CrossRef]
- Dong, R.; Wang, Y.; Lu, C.; Lei, G.; Wen, L. The seasonality of macroinvertebrate β diversity along the gradient of hydrological connectivity in a dynamic river-floodplain system. Ecol. Indic. 2021, 121, 107112. [Google Scholar] [CrossRef]
- Gray, D.; Harding, J. Braided river benthic diversity at multiple spatial scales: A hierarchical analysis of ß diversity in complex floodplain systems. J. N. Am. Benthol. Soc. 2009, 28, 537–551. [Google Scholar] [CrossRef]
- Melo, A.S.; Schneck, F.; Hepp, L.U.; Simões, N.R.; Siqueira, T.; Bini, L.M. Focusing on variation: Methods and applications of the concept of beta diversity in aquatic ecosystems. Acta Limnol. Bras. 2011, 23, 318–331. [Google Scholar] [CrossRef][Green Version]
- Castro, D.M.P.; Callisto, M.; Solar, R.R.C.; Macedo, D.R.; Fernandes, G.W. Beta diversity of aquatic invertebrates increases along an altitudinal gradient in a Neotropical mountain. Biotropica 2019, 51, 399–411. [Google Scholar] [CrossRef]
- Gonzales-Yrujillo, J.D.; Alonso-Moreno, Y.L. Habitat simplification changes temporal patterns of invertebrate beta diversity in a high-Andean stream. Neotrop. Biodivers. 2020, 6, 206–216. [Google Scholar] [CrossRef]
- Bowles, D.E.; Dodd, H.R. Aquatic Vegetation Monitoring in Springs at Ozark National Scenic Riverways, 2007–2015; Natural Resource Data Series NPS/OZAR/NRDS—2016/1044; National Park Service: Fort Collins, CO, USA, 2016; p. 21. [Google Scholar]
- Bowles, D.E. Resiliency and recovery of aquatic vegetation following scouring floods in two first-magnitude springs, Missouri, USA. Hydrobiology 2022, 1, 164–182. [Google Scholar] [CrossRef]
- Wang, H.; He, G. Rivers: Linking nature, life, and civilization. River 2022, 1, 25–3612. [Google Scholar] [CrossRef]
- Lusardi, R.A.; Nichols, A.L.; Willis, A.D.; Jeffries, C.A.; Kiers, A.H.; Van Nieuwenhuyse, E.E.; Dahlgren, R.A. Not all rivers are created equal: The importance of spring-fed rivers under a changing climate. Water 2021, 13, 1652. [Google Scholar] [CrossRef]
- Cui, Y.; Mahoney, E.; Herbowicz, T. Economic Benefits to Local Communities from National Park Visitation, 2011; Report No. 48824–6446; Michigan State University, Department of Community, Agriculture, Recreation, and Resource Studies: East Lansing, MI, USA, 2013; p. 36. [Google Scholar]
- Cullinane, T.C.; Huber, C.; Koontz, L. 2012 National Park Visitor Spending Effects: Economic Contributions to Local Communities, States, and the Nation; Natural Resource Report NPS/NRSS/EQD/NRR 2014/765; National Park Service: Fort Collins, CO, USA, 2014; p. 42. [Google Scholar]
- National Park Service (NPS). Ozark National Scenic Riverways, Final General Management Plan/Environmental Impact Statement; Ozark National Scenic Riverways: Van Buren, MO, USA, 2014; p. 616. [Google Scholar]



| Parameter | Springs | Tributaries | River | Kruskal-Wallis Test | Dunn’s Test (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| N | 6 | 13 | 23 | p-value | H | S-C | S-T | T-C |
| Discharge (m3/s) | 3.95 (1.69) | 0.35 (0.11) | 8.01 (2.74) | 0.008 | 9.51 | 0.35 | 0.006 | 0.01 |
| Temperature (°C) | 13.02 (0.44) | 11.57 (0.52) | 8.64 (0.58) | 0.0001 | 17.97 | 0.0012 | 0.19 | 0.003 |
| Dissolved oxygen (mg/L) | 9.36 (0.16) | 10.13 (0.25) | 11.64 (0.28) | 0.0003 | 16.43 | 0.0004 | 0.23 | 0.003 |
| Specific conductance (µm/cm) | 274.97 (13.44) | 323.09 (22.83) | 258.12 (7.47) | 0.005 | 10.40 | 0.21 | 0.21 | 0.001 |
| pH | 7.30 (0.02) | 7.94 (0.07) | 7.92 (0.08) | 0.0006 | 14.94 | 0.0002 | 0.0005 | 0.92 |
| % Periphyton | 27.11 (4.68) | 24.56 (0.82) | 43.43 (4.64) | 0.003 | 11.75 | 0.02 | 0.90 | 0.002 |
| % Vegetation | 45.11 (8.07) | 1.59 (1.29) | 2.08 (0.79) | 0.0007 | 13.40 | 0.001 | 0.0002 | 0.38 |
| % Filamentous green algae | 12.68 (1.88) | 3.60 (1.46) | 5.23 (1.69) | 0.002 | 12.11 | 0.002 | 0.0008 | 0.48 |
| Dominant substrate size (mm) | 59.91 (8.98) | 36.63 (2.11) | 42.12 (2.68) | 0.01 | 8.40 | 0.04 | 0.004 | 0.16 |
| % Embeddedness | 39.14 (2.90) | 36.12 (2.37) | 33.60 (2.39) | 0.29 | 2.42 | 0.13 | 0.39 | 0.46 |
| Taxa richness | 19.8 (2.94) | 23.67 (1.07) | 30.81 (1.52) | 0.0004 | 15.73 | 0.0003 | 0.16 | 0.008 |
| EPT richness | 6.80 (1.21) | 9.89 (0.79) | 17.13 (0.90) | 0.000002 | 25.74 | 0.000016 | 0.26 | 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heth, R.L.S.; Bowles, D.E. Aquatic Macroinvertebrate Community Structure along a Continuum in a Spring Dominated River, Missouri, USA. Hydrobiology 2022, 1, 518-530. https://doi.org/10.3390/hydrobiology1040031
Heth RLS, Bowles DE. Aquatic Macroinvertebrate Community Structure along a Continuum in a Spring Dominated River, Missouri, USA. Hydrobiology. 2022; 1(4):518-530. https://doi.org/10.3390/hydrobiology1040031
Chicago/Turabian StyleHeth, Rachel L. S., and David E. Bowles. 2022. "Aquatic Macroinvertebrate Community Structure along a Continuum in a Spring Dominated River, Missouri, USA" Hydrobiology 1, no. 4: 518-530. https://doi.org/10.3390/hydrobiology1040031
APA StyleHeth, R. L. S., & Bowles, D. E. (2022). Aquatic Macroinvertebrate Community Structure along a Continuum in a Spring Dominated River, Missouri, USA. Hydrobiology, 1(4), 518-530. https://doi.org/10.3390/hydrobiology1040031