Abstract
Reactive oxygen species comprise oxygen-based free radicals and non-radical species such as peroxynitrite and electronically excited (singlet) oxygen. These reactive species often have short lifetimes, and much of our understanding of their formation and reactivity in biological and especially medical environments has come from complimentary fast reaction methods involving pulsed lasers and high-energy radiation techniques. These and related methods, such as EPR, are discussed with particular reference to singlet oxygen, hydroxy radicals, the superoxide radical anion, and their roles in medical aspects, such as cancer, vision and skin disorders, and especially pro- and anti-oxidative processes.
1. Introduction
Reactive oxygen species (ROS) are involved in many biological and medical processes, ranging from photosynthesis to skin photosensitivity, vision, cancer, and sometimes areas of major commercial interest, such as skin protection and the preservation of items of historical importance. Much of this review concerns areas related to medicine, where ROS can have both deleterious and beneficial effects. It is because of the many deleterious processes that arise that anti-oxidants are of such importance in the study of ROS.
The wide range of ROS, from charged and neutral free radicals to non-radical oxidants—for example, singlet oxygen (1O2)—have been reviewed previously; see, for example, [1,2,3].
In the recent work of Halliwell [3], the role of the carbonate radical anion (CO3•−) with DNA was reported. Pulse radiolysis has shown that this radical can be generated from hydroxyl radical (•OH) reacting with the bicarbonate anion (HCO2−), and the relatively wide-ranging reactivity of •OH with DNA was compared with the more specific reactivity (with guanine) of CO3•−.
Di Mascio and co-workers have studied the generation of singlet oxygen (1O2) in biological systems—see, for example, [4]—showing the generation of 1O2 from lipid hydroperoxides and nitronium ions. This group has also shown that 1O2 is involved in the formation of covalent crosslinks involving histidine, lysine, and tryptophan [5].
Pospisil and co-workers have extensively studied the molecular mechanisms involving ROS, especially 1O2, including scavenging in photosynthetic systems such as photosystem II [6,7].
Dikalov and co-workers have reported on the induction of cardiovascular oxidative stress—for example, resulting from tobacco smoking (a source of ROS), which is shown to promote endothelial dysfunction and hypertension enhancement [8].
The major oxy-radicals of biological and medical interest in this review are the superoxide radical and its protonated form (O2•−/HO2•, pKa = 4.75 [9]), the hydroxyl radical (•OH), and various alkyl, allyl and sulfur-based radicals which add oxygen to give the corresponding peroxyl radicals (RO•, RO2•, and RSO2•). Meanwhile a (stable) free radical of current environmental as well as biological importance is nitrogen dioxide (NO2•, often written as just NO2).
Many studies have used fast reaction methods to study the radicals generated by the high-energy radiolysis of aqueous systems. The overall process leads to three free radicals (eaq, •OH, and hydrogen atoms H•) and several less reactive non-radical species. For the radiation methods relevant to this review, equal yields of eaq and •OH are generated, and the yield of H• is much smaller, nearly an order of magnitude less than the combined yield of eaq and •OH. All of the eaq and H• add to oxygen, producing the superoxide radical under normal conditions (both reactions are diffusion-controlled with rate constants of 2 × 1010 M−1s−1 in water [10]).
While not directly related to this review, the •OH radicals are frequently used to generate other, non-oxygen-containing radicals (see later), which, nevertheless, are oxidising and more specific in reactivity than •OH itself. Although these are non-oxygen-containing radicals, their study often helps to unravel the molecular mechanisms involving pro- and anti-oxidants with ROS.
The major non-radical species of interest is the activated oxygen molecule known as ‘singlet oxygen’ (1O2).
Usually, this particularly important oxidising species, 1O2, is generated via light absorption by a substrate, (Sub). This produces an activated reactive (triplet) state (3Sub*) via various fast intra-molecular processes, followed by a diffusional controlled energy transfer from the substrate triplet state to ground state oxygen, producing the reactive 1O2 and regenerating the substrate.
Sub + light → 3Sub* (the triplet state)
3Sub* + O2 → Sub + 1O2
The triplet lifetimes of most substrates of biological (and commercial) interest are sufficiently long for the energy transfer to be a significant process, thus generating the reactive (damaging) 1O2 that leads to major oxidative damage.
Other non-radical species can also be important, such as peroxynitrite/peroxynitrous acid (ONOO−/ONOOH) [1,11]. The pKa for these is 6.8, so both forms will arise in vivo.
2. Effect of Environment on ROS
Many ROS have short lifetimes and much of our understanding of their formation has come from complimentary fast reaction methods involving pulsed lasers and high energy techniques. The environment/solvent can be of particular importance for such studies.
2.1. Excited States
As noted above, the major interest is 1O2 generated via an energy transfer from an (excited) triplet state. The 1O2 lifetime is very dependent on its environment (the solvent), e.g., from 3.3 μs in water (H2O) to 55 μs in deuterated water (D2O) [12], and can be much longer in some deuterated hydrocarbons and carbon tetrachloride (26 ms) [13]. Nevertheless, even with a rather short lifetime in water, 1O2 can lead to the damage of a wide range of substrates/materials, from the skin, eyes, hair, and plants, to valuable paintings, fabrics, and other artefacts. Aspects of protection against such damage are discussed below.
The effect of environment on 1O2 lifetime may well explain the differing observations regarding 1O2 and β-carotene (β-CAR). Ogilby and co-workers [14] were able to detect 1O2 in individual HeLa mammalian cells with a microscope-based technique, but they could observe no quenching of the 1O2 by β-carotene. However, Telfer [15] and co-workers showed that carotenoids quench some, but not all, of the 1O2 in cells, due to the distance of the carotenoids from the source of the 1O2. This work, in photosynthetic systems, suggests that in a specific biological environment, quenching will only be efficient when the carotenoid is sufficiently close to the source of the 1O2.
For some substrates, the properties of the triplet state precursors of 1O2 can also be significantly dependent on the solvent. Good examples are the retinoids, including the visual pigments and their models.
All-trans- and 11-cis-retinal are, of course, the key chromophores in vision, and laser flash photolysis has been used extensively to investigate the production of triplet states and 1O2 from these pigments (see [16] for a recent review). Furthermore, retinol and other retinoids are important in the cosmetic industry, as well as in the treatment of some skin diseases. All isomeric forms of retinal have five conjugated carbon–carbon double bonds, plus a carbonyl group. The oxygen has non-bonding electrons (n), which lead to n-π* excited states, as well as the π-π* states associated with the conjugated carbon–carbon double bonds.
Pulsed laser studies in non-polar solvents lead to high yields of triplet states and 1O2 from all the retinal isomers. However, the environment (solvent dependence) is important with the triplet yields being much lower in polar solvents than in non-polar solvents.
In polar solvents, a state of 1π-π* character is located below 3n-π* and is the lowest excited singlet state. However, in non-polar, non-H-bonding solvents, a 1n-π* state which has a higher energy level than the 3π-π* is the lowest excited singlet state, meaning that a high yield of 1O2 is generated, leading to significant photo-oxidative damage. The 1O2 yield is much lower in polar solvents [17]; for example, in methyl alcohol, it is 0.08 for all-trans-retinal, whereas it is in the range of 0.2–0.5 in non-polar solvents [11].
2.2. Radicals
Generally, radicals do not significantly form in non-polar solvents via laser flash photolysis. However, laser excitation of substrates in polar solvents, such as alcohols, often leads to radical production, as well as the generation of excited states, and hence this environment is a route to study such radicals’ reactivity with a wide range of bio-molecules; see, for example, [18].
The fast reaction technique of pulse radiolysis (see below) has been extensively used to study specific free radicals, and, to some extent, excited states. Polar solvents, such as water and alcohols, support only extremely low yields of solute excited states and relatively high yields of radical ions, whereas non-polar solvents such as benzene and cyclohexane support high yields of solute excited states and relatively low yields of radical ions. Intermediate solvents, such as acetone, support similar yields of both excited states and radical ions.
The details of the processes leading to these environment/solvent effects have been reviewed [1,11].
The most important solvent for biological and medical interest is water, and in this review, we give only a summary of the aspects of free radical generation in aqueous conditions.
The early steps following the absorption of high-energy photons or ions (the radiolysis of water) involve the production of excited states of water; cations and electrons are also produced. Then, a variety of reactions occur, generating hydrogen atoms and hydroxyl radicals, and the electron loses energy via the excitation and ionisation of other molecules and becomes solvated (eaq).
Fast radical recombination processes occur, forming water; thus, within a nanosecond, in an argon- or nitrogen-saturated solution, the water radiolysis products are:
H2O ⇝ •OH + H• + eaq + H2O2 + H2 + H3O+
It is the three radical species (•OH + H• + eaq) which are the most reactive. The eaq and hydrogen atom (H•) are extremely reactive reductants, while the hydroxyl radical (•OH) is a highly (and often non-specific) oxidising species. So, as noted above, at all oxygen concentrations of interest, the eaq and H• add to oxygen to generate mainly O2•− at pH values above about 5. To understand the molecular processes involving O2•− (see below), it is normal to totally exclude oxygen, so that no O2•− is formed, and compare, for example, cell kill, with and without oxygen being present.
Normally, mechanistic studies involve using either oxidising or reducing environments, but not both, and specific procedures are used to achieve this.
For example, to selectively produce reducing conditions (often resulting in radical anions of the solutes under investigation), a relatively high concentration of sodium formate can be added. The formate anion is generated and reacts with the oxidising •OH radical and the H•, forming the carbon dioxide radical anion, CO2•−, which is reducing. Thus, the oxidising •OH radical has been replaced with only reducing species (solvated electron and CO2•−) and the overall effect is to generate exclusively the substrate radical anion.
Another method to obtain reducing conditions uses various alcohols which simply react with the •OH (and sometimes also the H•), generating the corresponding neutral radical of the alcohol, which is normally a reducing species.
Overall, oxidising (rather than reducing) environments are of more interest for biological and medical systems, and so it is important to also be able to generate predominantly oxidising conditions. This is normally achieved by saturating the solution with nitrous oxide gas (N2O), which reacts with the eaq to generate further oxidising •OH:
eaq + N2O → •OH + N2 + •OH
The H• also reacts with N2O, but at a slower rate, and this generates additional •OH, nitrogen, and water.
While •OH is regarded as a powerful oxidising species, it often also reacts by adding to the biological substrate of interest rather than the oxidation of the substrate to the corresponding radical cation. To generate such radical cations, weaker oxidant radicals are needed and are discussed below. One such method uses halide ions such as bromide or chloride. Radicals associated with chloride are, of course, important in many different environments including in vivo and in atmospheric and marine water studies. These are not discussed in this review.
3. Experimental Techniques
As noted above, many ROS are often transient species, which can be extremely short-lived, so the major techniques for their study have involved time-resolved methods, using a pulse of radiation to generate a specific ROS and then monitoring the reactivity of this ROS with systems of biological/medical/commercial interest. The techniques which have been extensively used are laser flash photolysis and pulse radiolysis, while 1O2 has been studied mainly by time-resolved near infra-red luminescence at near 1270 nm following pulsed laser activation [11]. These complementary methods are briefly summarised and compared below. In addition, some studies have used continuous radiation methods, using, for example, gamma (γ) rays from a Co-60 source. Moreover, since radicals are paramagnetic, there have been many studies that have used electron spin resonance.
3.1. LFP and PR
As noted above, both LFP and PR use a pulse of radiation to generate the ROS (often a nanosecond-long pulse, but shorter pulse times are also used where appropriate), followed by conventional absorption spectroscopy to monitor the absorption spectra and reactivity of each ROS individually. However, a few alternative monitoring methods have been used, such as resonance Raman spectroscopy [19]. While the initial excitation processes to produce the ROS are quite different from these two methods, both light and ionising radiation often give rise to ultimately similar ROS. Because of the different mechanisms which generate the ROS, it can be sometimes beneficial to apply both techniques. Often, we need information on excited states, especially the lowest triplet states of the solute and also on species such as photo-isomers, photoadducts, and photo-tautomers—flash photolysis is preferred for such studies. However, pulse radiolysis is often the preferred method for the generation of radicals. As noted above, the major method uses N2O to generate oxidising conditions (Equation (4)) but the •OH species often adds to the biological substrate rather than electron transfer to generate the important substrate radical cation. However, hydroxyl radicals can be converted into milder (one-electron) oxidants by the addition of halides (e.g., Br−), thiocyanate (SCN), or azide (N3−) ions, as shown in the following schemes:
•OH + Br− → Br• + •OH
Br• + Br− → Br2•−
•OH + SCN− → SCN• + •OH
SCN• + SCN− → (SCN)2•−
•OH + N3− → N3• + •OH
These milder oxidants (Br2•−, (SCN)2•− and N3•) then react with the biological substrate to generate the corresponding radical cation. Such cations may deprotonate as they are formed.
There are numerous PR studies of a wide range of bio-substrate radical cations obtained via such mild oxidising radicals. A typical example is the oxidation of the amino acid tryptophan to its radical cation, followed by deprotonation to a neutral radical at in vivo pH conditions:
N3• + TrpH → N3− + TrpH•+
TrpH•+ ⇌ Trp• + H+ (pKa = 4.3)
Such radical cations are themselves sufficiently strongly oxidising to react with dietary carotenoids to generate their radical cations via a one-electron transfer, as shown in Figure 1a,b for lycopene (LYC; λmax LYC•+ = 970 nm), and in Equation (12), while the deprotonated neutral tryptophan radical (Trp•) does not react efficiently with the lycopene:
TrpH•+ + LYC ⇌ TrpH + LYC•+
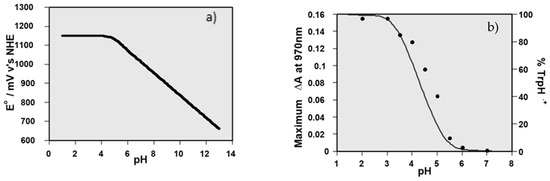
Figure 1.
(a) Plot showing the reduction potential of a tryptophan radical as a function of pH, calculated using a pKa of 4.3; (b) plot of the yield of LYC•+ as a function of pH (black dots) with the calculated percentage of TrpH•+ shown by the black line, based on a pKa of 4.3.
Such reactions were used to establish the reduction potentials of many dietary carotenoid radical cations, by measuring the equilibrium constant for Equation (12) at several pH values, and then obtaining the difference between the reduction potential of the TrpH•+ and the carotenoid radical cation using the Nernst equation [20,21].
3.2. Singlet Oxygen (1O2)
The most frequently used method to study singlet oxygen is based on the pulsed-laser excitation of a solute, as in LFP. The 1O2 is then generated via an energy transfer from the solute excited triplet state to ground state oxygen, but instead of using absorption spectroscopy, the weak but very specific phosphorescence from singlet oxygen—monitored at about 1270 nm, using an appropriate photodiode—is used.
The study of the generation and reactivity of 1O2 is relatively straightforward because the weak phosphorescence from 1O2 at near λmax 1270 nm is totally specific for this species and can be readily measured with time-resolved pulsed-laser excitation and near infra-red diode detectors. As noted above, the lifetime of 1O2 is very solvent-dependent, and D2O (lifetime 55 μs) is often used to study aqueous systems simply because a lifetime of 3.3 μs (in H2O) is too short for many subsequent reactions to be studied unless very high substrate concentrations are used.
As a typical example, Figure 2 shows the quenching of 1O2 (monitored at 1270 nm) by lutein, an important xanthophyll in the macular, and the lack of such quenching by aggregated lutein.
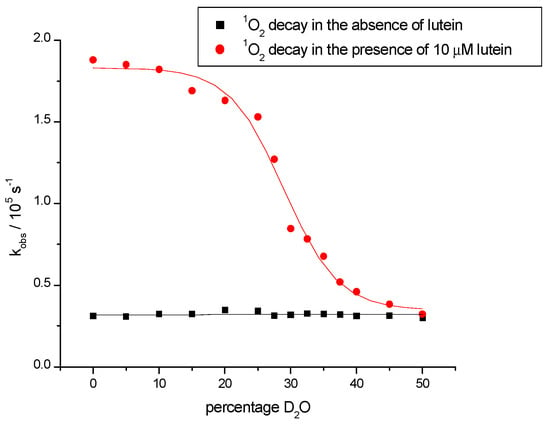
Figure 2.
Figure showing the first-order decay of 1O2 phosphorescence in the absence (black squares) and presence (red circles) of 10 μM lutein as a function of D2O percentage in deuterated methanol, illustrating lower quenching rate constants upon the aggregation of the lutein at higher D2O percentages.
3.3. Electron Paramagnetic Resonance (EPR) and Spin Trapping
A way of studying short-lived radicals or excited states without the need to use time-resolved spectroscopic techniques is to add a spin trap to produce a more stable nitroxyl radical, which can then be studied via standard spectroscopic techniques. This is a technique that is most often used in Electron Paramagnetic Resonance (EPR) spectroscopy (also often called Electron Spin Resonance or ESR).
It was over 50 years ago that free radical addition to nitrosoalkanes and nitrones was shown to produce nitroxide radicals, which are relatively stable due to resonance delocalisation along the N–O bond; see Figure 3 [22,23,24,25,26].
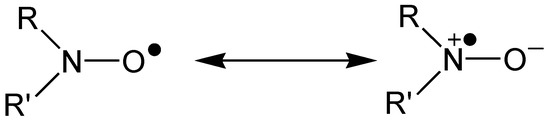
Figure 3.
Resonance stabilisation of a nitroxide radical.
Figure 4 shows how the addition of a free radical, X•, produces nitroxyl radicals from nitrones and nitrosoalkanes. Continuous-wave EPR spectroscopy can be used to detect these stable radicals, and the hyperfine splitting pattern of the resulting spectrum allows for the characterisation of X•:
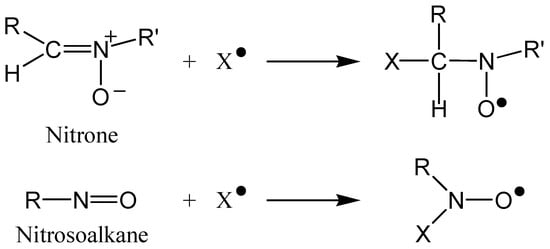
Figure 4.
Reaction of a nitrone and a nitrosoalkane with a free radical, X•, producing stable nitroxyl radicals.
In EPR, when the electron spin interacts with a non-zero nuclear spin, the energy levels are split, and so the spectrum is split into a number of lines. Nitrogen 14 has a nuclear spin (I) of 1, and so a three-line spectrum is observed for nitroxyl radicals. The number of lines observed is equal to 2I + 1.
Additionally, when X• contains nuclei with non-zero spin (e.g., Hydrogen; I = ½), further splitting of the spectrum occurs. The hyperfine splitting constant varies with the electron-withdrawing power of X•, and the magnitude of the splitting depends on the size of the nuclear magnetic moment and the fraction of electron spin density at the particular nucleus.
For example, EPR spin trapping has been used by Sachindra et al. [27] to determine the carotenoid fucoxanthin’s ability to quench both •OH and O2•−, showing the superior quenching of fucoxanthin when compared with two of its metabolites.
EPR has also been used to study 1O2 quenching with the stable nitroxyl radical TEMPO (2,2,6,6-tetramethylpiperidine-N-oxide), which is obtained from the reaction of 1O2 with TEMP (2,2,6,6-tetramethyl-1-piperidine). See, for example, [28], which uses spin trapping to show the scavenging of ROS (including 1O2) by N-substituted indole-2 and 3-carboxamides.
4. Applications
In this review, we discuss in detail five disparate areas where there are substantial new data: carotenoids in the eye, melanins and COVID-19, neonatal jaundice, PUVA treatment for psoriasis, and commercial sunscreens.
4.1. Carotenoids in the Eye
Age-related macular degeneration (AMD) is a major problem for older people. The macular contains three hydroxy-substituted carotenoids. Carotenoids containing oxygen are a sub-section called xanthophylls, and three xanthophylls—lutein (Lut), zeaxanthin (Zea), and meso-zeaxanthin (meso-Zea)—accumulate in the eye from normal diets, though many people also take these as dietary supplements. Another dietary carotenoid, lycopene (Lyc), is also thought to be beneficial against eye disease even though it does not accumulate in the macular—this is discussed below.
A major role of the three xanthophylls is protection, simply by light absorption. One reason for the presence of Lut, with only 10 conjugated C–C double bonds—compared to 11 for virtually all other dietary carotenoids and xanthophylls—is almost certainly its spectral shift. The absorption maximum is typically 445 nm for Lut compared to near 450 nm for Zea and meso-Zea, depending on the solvent [29]. However, these pigments also play other roles—indeed, as noted above, Lyc is not in the macular.
There have been numerous studies on the quenching of 1O2 by a wide range of carotenoids. The most efficient carotenoid quenchers have 11 or more conjugated C–C double bonds—typically, for Zea, about 12 × 109 M−1s−1. However, carotenoids with fewer C–C double bonds are less efficient—typically, for Lut, 6 × 109 M−1s−1. So, if the presence of lutein in the macular is for protection, this is not likely to be via 1O2 quenching.
One complication in such studies, which is still to be resolved, is the role of carotenoid aggregation [29,30]—the xanthophylls in the macular are at a high concentration (up to 1 mM) and aggregated xanthophylls do not quench 1O2 [31]. The role of the micro-environment on the 1O2 quenching efficiency of several carotenoids has also been reviewed recently. An interesting observation compares the efficiency of the carotenoid quenching of 1O2 when generated in the aqueous phase with that of when it is generated inside a liposome. No difference in efficiency was detected, suggesting that the location of carotenoids in cell membranes is not a significant factor with respect to macular protection against 1O2 [16].
More recently, the role of Zea, Lut, and other carotenoids, especially Lyc, with respect to interactions with oxyradicals and eye protection, has become of interest [32]. As noted above, the frequently used term, ROS, ‘hides’ any difference in the behaviour of different radicals, with respect to protection or even damage via their interactions with carotenoid ‘antioxidants.’ Three radicals, NO2•, •OH, and O2•−, have been individually generated, and the effects of a range of carotenoids, with respect to cellular protection/damage, have been reported.
The NO2• radical reacts with carotenoids via electron transfer to generate the radical cation of the carotenoid. Such radical cations are strong oxidising species themselves and can oxidise other biosubstrates [20,21]—clearly, this is a potential route to damage by NO2•. It has been shown that the concentration of oxygen plays no role in this process, but reducing agents such as ascorbic acid dramatically improves the protection of cells from NO2• damage. However, as noted above, there are claims [33] that dietary lycopene can protect against age-related macular deterioration (AMD), even though lycopene does not accumulate in the macular. One proposed mechanism to account for this protection is that Lyc acts as a sacrificial anti-oxidant and that any Lut or Zea lost via the generation of their radical cations is repaired by electron transfer processes such as:
Zea•+/Lut•+ + Lyc → Zea/Lut + Lyc•+
This is followed by the ascorbic acid reduction of Lyc•+ to regenerate the Lyc.
The interaction of •OH radicals with macular carotenoids is quite different to those of NO2• and shows a marked dependence on oxygen concentration. The three most important carotenoids for the eye are Zea, Lut, and Lyc, and here we compare the new results on Zea and Lut with the previous information on Lyc and some other dietary carotenoids [32]. The major technique used to generate the •OH was continuous radiation, using gamma (γ) rays from a Co-60 source. Using dietary carotenoids from commercial supplements, the protection of cell lymphocytes against •OH was measured as a function of the oxygen concentration. Typical results are presented in Figure 5. As can be seen, the cell protection due to Lyc, Zea, and β-Car falls steeply as the oxygen concentration increases and drops typically by up to an order of magnitude at 20% oxygen compared to 0% oxygen. At higher concentrations of oxygen, there is, within experimental error, no protection by these carotenoids. However, for Lut, the protection has only fallen by about 15% from 0% to 20% oxygen and is then more or less constant up to 100% oxygen. This comparison is clearly evident, as shown in Figure 6.
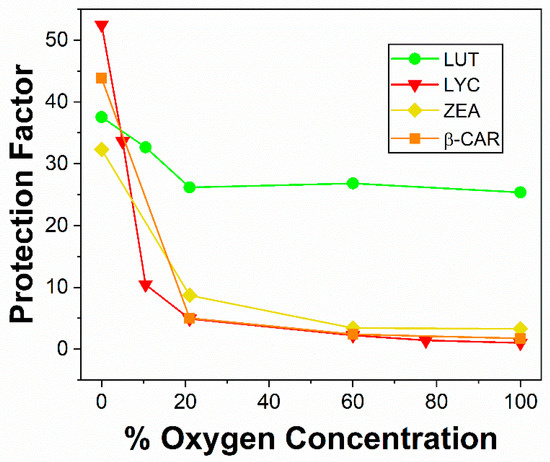
Figure 5.
Variation of mean protection factor for the protection of lymphocyte cells γ-irradiated with 2000 Gy dose with the percentage of oxygen concentration. Lyc (red triangle), β-Car (orange square), Zea (yellow diamond), and lutein (green circle).
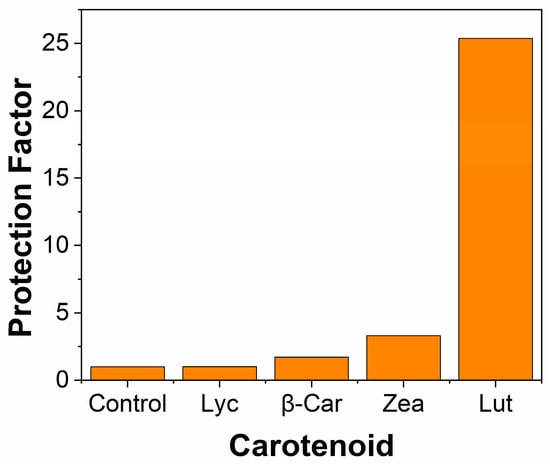
Figure 6.
Mean protection factors for the protection of lymphocyte cells γ-irradiated with 2000 Gy dose; 4 carotenoids are compared with the control at 100% oxygen.
The mechanism presented for the effects of oxygen with •OH is based on the formation of peroxyl radicals:
Car + •OH → Car-•OH
Car-•OH + O2 ⇌ Car-OH-OO•
Car-OH-OO• + cell → cell kill
So, we speculate for Lut that the equilibrium of 15 is more to the left than for the other carotenoids. Overall, these results suggest that Lut is a significantly better protector of the eye from •OH at moderate to high oxygen concentrations, as occur in the eye.
The superoxide radical anion, O2•−, is continually generated in normal metabolism. The first and most essential protector is superoxide dismutase (SOD). Studies at various oxygen concentrations showed, as with NO2•, no marked effect of the oxygen concentration. The γ-radiolysis source generates •OH and eaq as the two most significant reactive species and, at all oxygen concentrations [32], the solvated electron, eaq, is totally converted to O2•−. Cell protection for several carotenoids and SOD against a combination of •OH plus O2•− was obtained, as shown in the following table (Table 1). As can be seen, Lut markedly has the highest protection factor, and this is even more marked at higher oxygen concentrations (as seen in Figure 6, above). So, while SOD is, of course, the key in vivo protector against O2•−, it is possible that these ex vivo results also show efficient protection against O2•− by Lut, as well as protection for •OH. In summary, it seems a reasonable speculation that the efficient human cell protection given by Lut at all oxygen concentrations may suggest one reason why the macular accumulates Lut as well as Zea.

Table 1.
Mean protection factors for the protection of lymphocyte cells γ-irradiated with 2000 Gy dose; 4 carotenoids and Superoxide Dismutase (SOD) are compared at 21% oxygen.
4.2. Melanins and COVID-19
The precise structure of melanin is not established, and the term melanin is used to describe a range of pigments distributed widely in nature, including in humans (in the eye, hair, brain, ear, and, of course, the skin). Differences in skin pigmentation in humans are the most obvious sign of ethnic variation. Of importance is the fact that individuals with dark skin rarely get skin cancer, while light-skinned people are skin cancer-prone via light-induced skin damage. While such damage and ageing are not restricted to light-skinned people, it is more prevalent.
It has been recognised for many years that even though melanins are photoprotective they can also be photosenstisers and, therefore, lead to skin damage [34,35].
Basically, there are two types of melanin in the skin: eumelanin (EM), predominantly in dark skin, and pheomelanin (PM) in light skin. These species arise via a divergence in the melanin synthesis. Melanogenesis of both of these types starts with the generation of dopaquinone, the initial product of the oxidation of the amino acid tyrosine by tyrosinase (see, for example, [36]). This is followed by a divergence, with EM being formed via cyclodopa and dopachrome, while, for PM, the sulphur-containing amino acid cysteine adds to produce cysteinyldopa and then various benzothiazoles which are characteristic of PM [36].
This polymerisation takes place in cells called melanocytes, and the melanin is deposited in pigment granules called melanosomes. The degree of an individual’s skin pigmentation depends on several factors, such as the rate of melanin synthesis and the size of the pigment granules.
While EM can lead to a small degree of photosensitisation, it is an extremely efficient photoprotector. However, PM is quite different, and readily undergoes photo-reactions via sunlight absorption. It is these processes that can cause the susceptibility of fair-skinned people to the serious deleterious effects of sunlight absorption. Moreover, of course, there is deeper penetration of light in fair-skinned individuals compared to those with dark skin [37]. It is these processes that have also been speculated to be linked to the incidence and/or severity of COVID-19.
Many discussions and suggestions have concerned ethnic differences in the clinical outcomes of COVID-19 infection, and several possible factors have been discussed. These include socio-economic influences [38], vitamin D [39,40] levels, and the role of melanin due to iron chelation and subsequent trans-epidermal loss [41]. Indeed, a clear association has been observed between the death rate from COVID-19 infection and skin types IV to VI (brown to black) [42,43]. This correlation with skin pigmentation has led to the speculation that melanin is in some way implicated.
Overall, it seems likely that more than one factor is involved, and it is not yet well understood why COVID-19 is so damaging for BAME people. Therefore, it seems worthwhile to consider mechanisms involving epidermal pigmentation and photochemical processes in the skin, releasing ROS being one such possibility. Of course, there is an indirect photochemical link associated with vitamin D syntheses, but a more direct mechanism via photoreactions of the melanins and the generation of ROS is quite different and also worthy of consideration.
At least two photochemical-induced processes can arise. One is direct photo-production of various ROS species due to the effects of sunlight on PM and the second is related to light-induced structural rearrangements of the melanins to release the ROS which have accumulated during their biosynthesis.
Drawing on data from previous melanin/radical studies, a speculative suggestion [44] is that ROS, including singlet oxygen and superoxide, might escape melanocytes more easily in those individuals who have a higher percentage of PM in their skin pigmentation. These radicals could then cause subsequent damage to SARS-CoV-2, the virus which causes COVID-19. Possibly related is the complex behaviour of ROS with pathogens, where the precise conditions can lead to ROS-mediated pathogen killing or adaption, allowing pathogens to defend or thrive under ROS conditions [45].
Additionally, it has been postulated that carbon and sulphur peroxy radicals, may be produced in the skin via oxygen addition to pheomelanin radicals formed by carbon-sulphur bond cleavage following light absorption. The subsequent structural rearrangement of the large melanin molecules could then lead to the release of pre-absorbed ROS, also promoting enhanced COVID-19 protection in the fair-skinned. Another possibly significant difference for EM and PM arises with the carbonate radical anion. In the presence of •OH, the carbonate radical anion is converted to the strongly oxidising CO3•−, as noted above [3]. Using dopa melanin as a model of black eumelanin and cysteinyl-dopa-melanin as a model of red/blond phaeomelanin, we have reported huge differences in their reactivity with CO3•− (3 × 105 M−1s−1 and 2 × 108 M−1s−1, respectively) [46]. Such a difference (near 103) is much more marked than with other bio-molecules (typically 10 times for amino acids).
Certainly, photochemical studies of PM models have shown the increased production of singlet oxygen, superoxide, and both carbon and sulphur-based radicals [47,48,49], while the aggregation of EM has been shown to diminish the photogeneration of ROS [50].
Recent EPR studies have also shown the photo-production of ROS from melanins in melanocytes and skin itself [51,52]. In Kassouf et al. [51], the spin trapping of radicals produced via UVA irradiation of melanocytes and melanoma cells showed higher levels of spin trapped carbon-centred radicals in lightly pigmented cells than in darkly pigmented or even non-pigmented cells. Albrecht et al. [52] used EPR spin trapping to detect radicals directly in irradiated skin and found radical production when the skin was irradiated with either UV, visible, or even infra-red light. Radical production in skin types IV–V (dark skin) was found to be only around 60% of the total amount observed in skin type II (fair skin).
In summary, we speculate there may be a link between the increased photo-induced radicals in fair-skinned individuals compared to dark-skinned, leading to higher severity of COVID-19 in the dark-skinned BAME community, with partial protection for fair-skinned individuals arising from ROS, generated from both the photolysis of PM and its photo-induced rearrangement.
4.3. Psoriasis and UVA
The skin disease psoriasis has been treated by a photochemotherapeutic system for many years. The possible deleterious effects involving free radicals and 1O2 are important and are discussed below. The treatment is known as PUVA, where the ‘P’ refers to the drug 8-methoxypsoralen (8-MOP), and UVA to long-wavelength ultra-violet radiation (320–400 nm). The 8-MOP is administered by either oral or topical application, followed (typically 45–60 min later) by UVA irradiation. While much of the underlying biological processes are still to be understood, one mechanism invokes UVA absorption, producing the excited triplet state of 8-MOP, which then interacts (possibly via a cyclobutane formation) with the epidermal DNA, inhibiting further DNA replication. However, there are unwanted side effects, and one such problem is the generation of various ROS, such as •OH, O2•−, and 1O2, as shown by EPR studies [53,54] and the time-resolved luminescence of 1O2 [55].
Such ROS may lead to pro-oxidative effects and to carcinogenesis and mutagenesis. For that reason, PUVA is often restricted in use for some age groups.
A second side effect is an increase in skin pigmentation. While the mechanism of this skin-darkening effect is not fully understood, one possibility involves an electron transfer between the 8-MOP triplet state (3MOP) and the melanin precursor 3,4-dihydroxy-phenylalanine (dopa) [56] this can be summarised as:
3MOP + dopa 🡢🡢 dopachrome 🡢🡢 melanin
Dopachrome is a well-known key intermediate in the free radical polymerisation of dopa to melanin [36].
This deleterious effect can lead to reduced PUVA efficiency, and, therefore, the requirement to increase the UVA dose, thus increasing the potential skin carcinogenicity/mutagenicity risk.
4.4. Neonatal Jaundice
The phototherapy of newborn infants with blue/green light is the established (often called ‘gold standard’) therapy for neonatal hyperbilirubinemia following its introduction nearly 60 years ago. While the detailed molecular mechanism is not fully understood, clearly this phototherapy depends on the photochemical isomerisation of bilirubin (BR) within light-exposed tissues, leading to excretion without having to undergo further metabolic modification. Overall, while BR shows no fluorescence itself, three photochemical reactions of bilirubin occur: photooxidation, configurational isomerization, and structural isomerization [57]. The configurational isomerization leads to the production of Z,E isomers (from the original Z,Z isomer) and a structural isomer called Lumirubin (LR), as well as BR fragments (mono-, di-, and tripyrrolic species) via oxidation. The configurational isomers are likely to be established very quickly (maybe sub-nanosecond) and be in equilibrium—it seems unlikely that they would lead to ROS such as singlet oxygen or oxy radicals. However, the structural isomer Z-LR contains a seven-membered ring and significant double bond/single bond conjugation (see Figure 7).
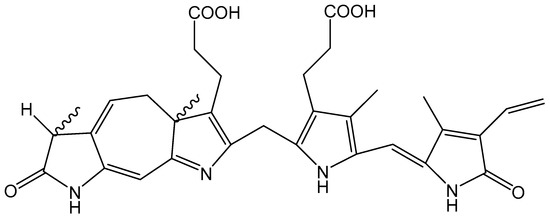
Figure 7.
Chemical structure of Lumirubin.
The UV/visible spectrum has recently been reported [58], showing light absorption between about 380 and 500 nm for both BR and LR (with a minor spectral shift due to binding to human serum albumin (HSA)). Subsequently, oxidation processes from further light absorption by LR seem the most likely route to the overall destruction of the free BR and its excretion. Overall, we would suggest processes such as:
this is followed by an energy transfer to molecular oxygen to produce the potentially damaging species, singlet oxygen:
with other reactions, such as electron transfer processes, this possibly leads to other (radical) ROS.
LR + light 🡢 1LR* fluorescence (λmax = 415 nm) + LR triplet (3LR*)
3LR* + O2 🡢 LR + 1O2
1O2 + BR 🡢 BR photoproducts
Singlet oxygen has a very specific luminescence in the infra-red at near 1270 nm. The observation of this luminescence means 1O2 is being formed. In one experiment, BR dissolved in carbon tetrachloride was irradiated with 514.5 nm light (argon ion laser) and a very marked increase in the luminesce signal at 1270 nm was observed over a 30 min period, proving the generation of 1O2. In addition, during this period, the solution became colourless, proving the destruction of the BR and LR pigments [59]. More recently, the effects of BR and LR (the major BR photo-oxidation product) on metabolic and oxidative stress markers have been compared [60]. These researchers estimated the effects of BR and LR in a cell-based study on peroxy radicals, O2•− and nitric oxide. LR stability was tested under different oxygen concentrations—showing an increase in the LR degradation as the oxygen concentration was increased from 1% to 21%—and seemed likely to result in the production of mono-, di- and tripyrrolic fragments. Collectively, these data point to the biological effects of BR and its photo-oxidation products, which might have clinical relevance in phototherapy-treated hyperbilirubinemic neonates and adult patients.
While phototherapy treatment on children with enhanced non-bound BR is clearly far more valuable than any possible drawbacks, the potential deleterious side effects may be linked to LR light excitation, leading to ROS production as an early step.
4.5. Commercial Sunscreens
The first report on the use of sunscreens to protect the skin was nearly 100 years ago—although, of course, humans have used various ways of protecting themselves from the sun since antiquity. Studies on sunscreens up to 1990 have been thoroughly reviewed [61]. Commercial sunscreens can be divided into two broad categories: inorganic and organic. Inorganic sunscreens mainly scatter UV radiation, while the organic systems absorb the radiation and then lose the energy as heat. In practice, mixtures of both are normally formulated together for the commercial market.
4.5.1. Inorganic Sunscreens
The use of inorganic sunscreens began around 60 years ago, and both titanium dioxide (TiO2) and zinc oxide (ZnO) are used [62]. TiO2 is by far the most important inorganic sunscreen and gives protection against both UVB and UVA2 (320–340 nm); the protection ranges from 315 to 340 nm, and there is no protection against UVA1 wavelengths (340–400 nm). One important aspect of both TiO2 and ZnO is that they are inert, and so do not interfere with the other (organic) sunscreens in a typical commercial product. Early formulations of TiO2 used a particle size which led to, at least for adults, an unacceptable white cosmetic appearance upon the skin. Nowadays, smaller, so-called micronized TiO2, which is transparent, with particle sizes typically around 10–20 nm, is used. However, such preparations have been questioned as a source of potentially damaging (ROS) species when irradiated, possibly because of the increased surface area compared to larger particles. Of course, TiO2 is widely used as a photosensitiser for many industrial applications, such as water treatment (see, for example, [63]), and as a route to solar-generated electricity. Clearly, the photosensitised generation of ROS must be avoided for sunscreen use, and there has been much debate on this possible deleterious effect: [64,65,66].
A major factor concerns the type of TiO2 used. There are three types: rutile, anatase, and brookite. Rutile has the lowest photoactivity and so is the preferred choice for sunscreens and cosmetic applications, while anatase, a very photoactive form, is used where photoactivity is required, such as for the degradation of pollutants in water. Another important factor is surface coatings—many materials are used and can not only reduce photoactivity but also help to avoid other unwanted commercial problems, such as the aggregation of the TiO2 particles. Another important aspect is the addition of antioxidants to the commercial product, such as ascorbic acid and vitamin E. A major discussion topic is the degree of penetration of the inorganic sunscreens into the skin. Probably most important is that, in order for ROS to be a damaging factor, the TiO2 and ZnO would have to penetrate beyond the outer layer of the skin, which seems unlikely [62].
4.5.2. Organic Sunscreens
Numerous organic molecules have been developed and are in current use as sunscreens. Generally, most are regarded as UVA or UVB absorbers, though there are many more UVB absorbers in use than UVA absorbers. A major concern is the photostability of such sunscreens, with much current interest concerning the important UVA sunscreen 4-tert-butyl-4′-methoxydibenzoylmethane (BM-DBM). This widely used sunscreen has several different trade names in different parts of the world; the most common are Avobenzone and Parsol 1789.
The strong absorbance of BM-DBM has a λmax which depends, to some degree, on the solvent, but is about 360 nm. However, this widely used UVA sunscreen is unstable, mainly because it degrades upon exposure to UV radiation. BM-DBM was approved for use in the EU in 1978 and by the FDA in 1988. However, shortly afterwards, based on studies of model compounds, its safety with respect to phototoxicity and photo-mutagenicity was brought into question [67].
Several studies of the photochemical degradation of BM-DBM have been undertaken with considerable interest in methods to reduce such degradation. Studies on the effect of solvents showed that BM-DBM was more or less photostable in the polar protic solvent methanol but was photoisomerised in the polar aprotic solvent dimethylsulfoxide, and it photodegraded significantly in the nonpolar solvent cyclohexane [68]. Both photoisomerisation and photodegradation occurred to a similar extent in the moderately polar aprotic solvent ethyl acetate [68].
BM-DBM is a complex molecule and, in solution, it is an equilibrium mixture of enolic and diketo forms. The enols can be both chelated and non-chelated ‘rotamers,’ as shown in Figure 8, but the chelated enol form, absorbing around 360 nm, is the dominant form, with the keto isomer produced during UV irradiation.
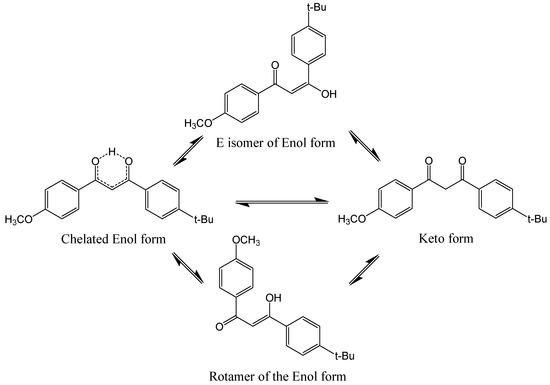
Figure 8.
Photoinduced ketonization of BM-DBM via an intermediate excited state rotamer transient.
While the keto form has a λmax near 260 nm (UVC), it has an absorption ‘tail’ extending through the UVB. Laser flash photolysis studies show the existence of short-lived transient forms which are assigned to the non-chelated rotamers [69,70], as shown in Figure 8, with a λmax near 300 nm. The efficiency of the conversion from the initial enol form to these short-lived transient forms is quite high, at 0.25 [71]. The decay of the short-lived transients are assumed to be both reversions back to the original enol and also to the formation of the diketo form—it is this form that almost certainly leads to the destruction of the sunscreen BM-DBM. Further evidence of this is based on model studies on the ‘locked’ form of the diketo isomer, where a simple methyl substitution results in a 100% diketo form. A quantum yield of 1O2 formation of 0.3 from this model was reported [72]. Consistent with this quite high efficiency of 1O2 formation, it has been shown that when the diketo form is generated by the pre-irradiation of BM-DBM [70], subsequent pulsed laser irradiation (266 nm, UVC) leads to the detection of damaging 1O2 (monitored as described above at 1270 nm), which leads to the oxidation of BM-DBM. The decay of the enol form and the corresponding formation of the diketo form has been also demonstrated by others [73]. Furthermore, these diketones are liable to also be degraded by other photochemical processes, such as α-cleavage (also called Norrish Type 1 processes), generating a range of undesirable free radicals. Overall, this is an extremely complex system, and recent attempts to unravel the underlying photochemistry have involved gas-phase studies of protonated BM-DBM [74].
As noted above, the deleterious effects of such processes are reduced by antioxidants such as vitamin C and E. However, there is still considerable interest in the deleterious effects associated with 1O2 generation for the diketo form of BM-DBM. Of course, 266 nm (UVC) is not a wavelength that reaches earth, but, as noted above, while the BM-DBM diketo has a λmax near 260 nm, it also has weak absorption in the UVB and even UVA spectral region (and even out to 450 nm). It seems possible this long-wavelength weak absorption is a key factor in the final photodegradation of BM-DBM. The generation of 1O2 from diketo absorption occurs via the production of the triplet form of the diketo, which undergoes an energy transfer with oxygen to give the damaging 1O2, as discussed above, and shown in the simplified Equations (21) and (22), below:
BM-DBM + Light 🡢🡢 3BM-DBM*
3BM-DBM* + O2 🡢 BM-DBM + 1O2
Clearly, to overcome this problem, other (anti-oxidant) molecules which can efficiently remove (quench) the triplet states are worthwhile. Many combinations of sunscreens—usually mixing UVB absorbers with BM-DBM—have been considered [72,75] to improve photostability.
Of course, for such a mixture, an increase in the stability of BM-DBM can be expected upon the addition of a second sunscreen, arising simply due to the additive effect of the two absorbances. However, if the protection of the BM-DBM is increased by more than this ‘additive’ effect, it would suggest the occurrence of other interactions involving these sunscreen mixtures. One such possibility is that the BM-DBM triplet state undergoes a triplet–triplet energy transfer to the second, protecting sunscreen. Of course, for this to arise, the triplet energy level of the second sunscreen needs to be below that of the BM-DBM diketo form. Unfortunately, the triplet energy levels of UVB sunscreens are not well established.
One important combination is BM-DBM with octocrylene—while there does not seem to be an experimental value of the octocrylene triplet level available, the theoretical value (60.7 kcal·mol−1) sets the octocrylene triplet level well below that of the diketo triplet (established via phosphorescence as 70 kcal·mol−1 [76]) and so would allow for a very efficient energy transfer from the diketo triplet to octocrylene, and hence the protection of BM-DBM from photodegradation [77].
The triplet lifetime of the keto form is expected to be shorter than that of the enol form because of the short-lived character of the 3nπ* state. However, the observed triplet lifetime of the keto form of BM-DBM is about six times longer than that of the enol form [77]. This unexpected effect of the tautomerization on the triplet lifetime of BM-DBM may arise from a fast, nonradiative decay channel in the 3nπ* state of the enol form. Such an increase in lifetime, if also applicable to real-life situations, may well lead to the increased photodegradation of the sunscreen.
Of course, in real-life situations, the rate of collision between different sunscreens will be well below that observed in many solution-based model studies and, clearly, it will also depend on the concentrations used. Another (possibly more likely) process is that the diketo form produces 1O2 but this (mobile) ROS species is significantly quenched by reacting with the secondary UVB sunscreens.
Overall, more data is needed to unravel the photodegradation of BM-DBM. Indeed, it is perhaps surprising that UVB degrades the BM-DBM diketo form at all, given that it is a UVC absorber.
5. Summary
ROS consist of a complex mixture of species ranging from non-radical 1O2 to a wide range of oxy-radicals. Overall, the most important and studied species are 1O2, NO2•, •OH, O2•−, and various peroxyl species (RO2•).
Usually, ROS are damaging species, but in some situations, they play a useful, beneficial role rather than a deleterious role—for example, in water purification and in the treatment of some diseases such as skin cancer. In some cases, there is a possible minor deleterious effect associated with an important beneficial treatment, as in the photochemotherapy of psoriasis and possibly the phototherapy of neonatal jaundice (hyperbilirubinemia).
In this review, we have attempted to unravel the molecular mechanisms of both deleterious and beneficial effects via studies of some of the important individual ROS species. Such studies require the generation of ROS and, usually, very short-lived specific radicals or 1O2 are studied rather than the ROS mixture itself. For such fast-reaction studies, pulse radiation methods are most useful and generally use pulsed lasers to study 1O2 and pulse radiolysis to study radicals, although there is an overlap in the use of these fast reaction techniques, and often both techniques are used to give complementary information. However, in some cases, EPR spin trapping can be used and fast techniques are not required.
Clearly, as we have shown, more work is needed to maximise the beneficial effects of individual ROS species while minimising any unwanted deleterious side effects.
Author Contributions
Conceptualization, R.E. and T.G.T.; writing—original draft preparation, R.E. and T.G.T.; writing—review and editing R.E. and T.G.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
R.E. acknowledges the support of The University of Manchester’s Dalton Cumbrian Facility (DCF), a partner in the National Nuclear User Facility, the EPSRC UK National Ion Beam Centre, and the Henry Royce Institute.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Edge, R. Radiolytic and photolytic production of free radicals and reactive oxygen species: Interactions with antioxidants and biomolecules. In Applied Photochemistry; Evans, R.C., Douglas, P., Burrows, H.D., Eds.; Springer Science: Dordrecht, The Netherlands, 2013; pp. 305–330. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Halliwell, B.; Adhikary, A.; Dingfelder, M.; Dizdaroglu, M. Hydroxyl radical is a significant player in oxidative DNA damage in vivo. Chem. Soc. Rev. 2021, 50, 8355–8360. [Google Scholar] [CrossRef]
- Prado, F.M.; Scalfo, A.C.; Miyamoto, S.; Medeiros, M.H.G.; Di Mascio, P. Generation of Singlet Molecular Oxygen by Lipid Hydroperoxides and Nitronium Ion. Photochem. Photobiol. 2020, 96, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Marques, E.F.; Medeiros, M.H.G.; Di Mascio, P. Singlet oxygen-induced protein aggregation: Lysozyme crosslink formation and nLC-MS/MS characterization. J. Mass. Spec. 2019, 54, 894–905. [Google Scholar] [CrossRef]
- Pospíšil, P.; Ankush Prasad, A. Formation of singlet oxygen and protection against its oxidative damage in Photosystem II under abiotic stress. J. Photochem. Photobiol. B Biol. 2014, 137, 39–48. [Google Scholar] [CrossRef]
- Pospíšil, P. Molecular mechanisms of production and scavenging of reactive oxygen species by photosystem II. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 218–231. [Google Scholar] [CrossRef]
- Dikalov, S.; Itani, H.; Richmond, B.; Vergeade, A.; Rahman, S.M.J.; Boutaud, O.; Blackwell, T.; Massion, P.P.; Harrison, D.G.; Dikalova, A. Tobacco smoking induces cardiovascular mitochondrial oxidative stress, promotes endothelial dysfunction, and enhances hypertension. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H639–H646. [Google Scholar] [CrossRef]
- Bielski, B.H.J.; Allen, A.O. Mechanism of the disproportionation of superoxide radicals. J. Phys. Chem. 1977, 81, 1048–1050. [Google Scholar] [CrossRef]
- Bielski, B.H.J.; Cabelli, D.E.; Arudi, R.L.; Ross, A.B. Reactivity of HO2/O−2 radicals in aqueous solution. J. Phys. Chem. Ref. Data 1985, 14, 1041–1100. [Google Scholar] [CrossRef]
- Bensasson, R.V.; Land, E.J.; Truscott, T.G. Excited States in Biology and Medicine; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Rodgers, M.A.J. Time resolved studies of 1.27 μm luminescence from singlet oxygen generated in homogeneous and microheteregeneous fluids. Photochem. Photobiol. 1983, 37, 99–103. [Google Scholar] [CrossRef]
- Salokhiddinov, K.I.; Dzhagarov, B.M.; Byteva, I.M.; Gurinovich, G.P. Photosensitized luminescence of singlet oxygen in solutions at 1588 nm. Chem. Phys. Lett. 1980, 76, 85–87. [Google Scholar] [CrossRef]
- Bosio, G.N.; Breitenbach, T.; Parisi, J.; Reigosa, M.; Blaikie, F.H.; Pedersen, B.W.; Silva, E.F.F.; Martire, D.O.; Ogilby, P.R. Antioxidant β-carotene does not quench singlet oxygen in mammalian cells. J. Am. Chem. Soc. 2013, 135, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Telfer, A. Singlet oxygen production by PSII under light stress: Mechanism, detection and the protective role of β-carotene. Plant Cell Physiol. 2014, 55, 1216–1223. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. Singlet oxygen and free radical reactions of retinoids and carotenoids—A review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef]
- Dillon, J.; Gaillard, E.R.; Bilski, P.; Chignell, C.F.; Reszka, K.J. The photochemistry of the retinoids as studied by steady-state and pulsed methods. Photochem. Photobiol. 1996, 63, 680–685. [Google Scholar] [CrossRef]
- El-Agamey, A.; Fukuzumi, S. Laser flash photolysis study on the retinol radical cation in polar solvents. Org. Biomol. Chem. 2011, 9, 6437–6446. [Google Scholar] [CrossRef]
- Teraoka, J.; Hashimoto, H.; Matsudaira, S.; Koyama, Y. Resonance Raman spectra of excited triplet-states of β-carotene isomers. Chem Lett. 1985, 14, 311–314. [Google Scholar] [CrossRef]
- Edge, R.; Land, E.J.; McGarvey, D.J.; Burke, M.; Truscott, T.G. The reduction potential of the β-Carotene•+/β-Carotene couple in an aqueous micro-heterogeneous environment. FEBS Lett. 2000, 471, 125–127. [Google Scholar] [CrossRef]
- Burke, M.; Edge, R.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. One-electron reduction potentials of dietary carotenoid radical cations in aqueous micellar environments. FEBS Lett. 2001, 500, 132–136. [Google Scholar] [CrossRef]
- Mackor, A.; Wajer, T.A.J.W.; de Boer, T.J.; van Voorst, J.D.W. C-nitroso compounds. Part I. The formation of nitroxides by photolysis of nitroso compounds as studied by electron spin resonance. Tetrahedron Lett. 1966, 7, 2115–2123. [Google Scholar] [CrossRef]
- Mackor, A.; Wajer, T.A.J.W.; de Boer, T.J.; van Voorst, J.D.W. C-nitroso compounds. Part III. Alkoxy-alkyl-nitroxides as intermediates in the reaction of alkoxy-radicals with nitroso compounds. Tetrahedron Lett. 1967, 8, 385–390. [Google Scholar] [CrossRef]
- Mackor, A.; Wajer, T.A.J.W.; de Boer, T.J. C-nitroso compounds—VI: Acyl-alkyl-nitroxides from acyl radicals and nitroso compounds as studied by ESR. Tetrahedron Lett. 1968, 9, 1623–1631. [Google Scholar] [CrossRef]
- Janzen, E.G.; Blackburn, B.J. Detection and identification of short-lived free radicals by an electron spin resonance trapping technique. J. Am. Chem. Soc. 1968, 90, 5909–5910. [Google Scholar] [CrossRef]
- Iwamura, M.; Inamoto, N. Reactions of Nitrones with Free Radicals. II. Formation of Nitroxides. Bull. Chem. Soc. Jpn. 1970, 43, 860–863. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef]
- Aboul-Enein, H.Y.; Kruk, I.; Lichszteld, K.; Michalska, T.; Kladna, A.; Marczynski, S.; Ölgen, S. Scavenging of reactive oxygen species by N-substituted indole-2 and 3-carboxamides. Luminescence 2004, 19, 1–7. [Google Scholar] [CrossRef]
- Polivka, T. Effects of self-assembled aggregation on excited states. In Carotenoids: Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; Chapter 8; pp. 137–157. [Google Scholar]
- Sliwka, H.-R.; Partali, V.; Lockwood, S.F. Hydrophilic carotenoids: Carotenoid aggregates. In Carotenoids: Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; Chapter 3; pp. 31–58. [Google Scholar]
- Edge, R.; Truscott, T.G. Properties of carotenoid radicals and excited states and their potential role in biological systems. In Carotenoids: Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; Chapter 14; pp. 283–307. [Google Scholar]
- Boehm, F.; Edge, R.; Truscott, T.G. Anti- and pro-oxidative mechanisms comparing the macular carotenoids zeaxanthin and lutein with other dietary carotenoids—A singlet oxygen, free-radical in vitro and ex vivo study. Photochem. Photobiol. Sci. 2020, 19, 1001–1008. [Google Scholar] [CrossRef]
- Mares-Perlman, J.A.; Brady, W.E.; Klein, R.; Klein, B.E.; Bowen, P.; Stacewicz-Sapuntzakis, M.; Palta, M. Serum antioxidants and age-related macular degeneration in a population-based case-control study. Arch. Ophthalmol. 1995, 113, 1518–1523. [Google Scholar] [CrossRef] [PubMed]
- Hill, H.Z.; Li, W.; Xin, P.; Mitchell, D.L. Melanin: A two edged sword? Pigment Cell Res. 1997, 10, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Premi, S.; Wallisch, S.; Mano, C.M.; Weiner, A.B.; Bacchiocchi, A.; Wakamatsu, K.; Bechara, E.J.H.; Halaban, R.; Douki, T.; Brash, D.E. Chemiexcitation of melanin derivatives induces DNA photoproducts long after UV exposure. Science 2015, 347, 842–847. [Google Scholar] [CrossRef]
- Thomson, A.; Land, E.J.; Chedekel, M.R.; Subbaro, K.V.; Truscott, T.G. A pulse radiolysis investigation of the oxidation of the melanin precursors 3,4-dihydroxyphenylalanine (dopa) and the cysteinyldopas. Biochim. Et Biophys. Acta 1985, 843, 49–57. [Google Scholar] [CrossRef]
- Mustafa, F.H.; Jaafar, M.S. Comparison of wavelength-dependent penetration depths of lasers in different types of skin in photodynamic therapy. Indian J. Phys. 2013, 87, 203–209. [Google Scholar] [CrossRef]
- Khunti, K.; Platt, L.; Routen, A.; Abbasi, K. COVID-19 and ethnic minorities: An urgent agenda for overdue action. BMJ 2020, 369, m2503. [Google Scholar] [CrossRef] [PubMed]
- Ilie, P.C.; Stefanescu, S.; Smith, L. The role of vitamin D in the prevention of coronavirus disease 2019 infection and mortality. Aging Clin. Exp. Res. 2020, 32, 1195–1198. [Google Scholar] [CrossRef]
- Israel, A.; Cicurel, A.A.; Feldhamer, I.; Dror, Y.; Giveon, S.M.; Gillis, D.; Strich, D.; Lavie, G. The link between vitamin D deficiency and COVID-19 in a large population. medRxiv bioRxiv 2020. [Google Scholar] [CrossRef]
- Riley, P.A.; Truscott, T.G. Does iron chelation by melanin explain the ethnic link with COVID-19 fatality? Biomed. J. Sci. Tech. Res. 2020, 31, 24527–24528. [Google Scholar]
- Khunti, K.; Platt, L.; Routen, A.; Abbasi, K. Ethnicity and COVID-19. BMJ 2020, 369, m2282. [Google Scholar]
- Pareek, M.; Bangash, M.N.; Pareek, N.; Pan, D.S.; Sze, S.; Minhas, J.S.; Hanif, W.; Khunti, K. Ethnicity and COVID-19: An urgent public health research priority. Lancet 2020, 395, 1421–1422. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. COVID-19 and the ethnicity link—Is there a photochemical link? Photochem. Photobiol. Sci. 2021, 20, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, X.; Huang, Y.; Liao, B.; Cheng, L.; Ren, B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front. Microbiol. 2021, 11, 622534. [Google Scholar] [CrossRef]
- Clarke, K.; Edge, R.; Johnson, V.; Land, E.J.; Navaratnam, S.; Truscott, T.G. The carbonate Radical: Its reactivity with oxygen, ammonia, amino acids and melanins. J. Phys. Chem. A 2008, 112, 10147–10151. [Google Scholar] [CrossRef] [PubMed]
- Chedekel, M.R.; Smith, S.K.; Post, P.W.; Pokora, A.; Vessell, D.L. Photodestruction of pheomelanin: Role of oxygen. Proc. Natl. Acad. Sci. USA 1978, 75, 5395–5399. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Sinclair, R.S.; Truscott, T.G.; Land, E.J.; Chedekel, M.R.; Liu, C.-T. Photochemistry of benzothiazole models of pheomelanin. Photochem. Photobiol. 1984, 39, 5–10. [Google Scholar] [CrossRef]
- Land, E.J.; Thompson, A.; Truscott, T.G.; Subbarao, K.V.; Chedekel, M.R. Photochemistry of melanin precursors: Dopa, 5-S-cysteinyldopa and 2,5-S S’-dicysteinyldopa. Photochem. Photobiol. 1986, 44, 697–702. [Google Scholar] [CrossRef]
- Nofsinger, J.B.; Lui, Y.; Simon, J.D. Aggregation of eumelanin mitigates photogeneration of reactive oxygen species. Free Radic. Biol. Med. 2002, 32, 720–730. [Google Scholar] [CrossRef]
- Kassouf, N.; Kay, C.W.M.; Volkov, A.; Chiang, S.-H.; Birch-Machin, M.A.; El-Khamisy, S.F.; Haywood, R.M. UVA-induced carbon-centred radicals in lightly pigmented cells detected using ESR spectroscopy. Free Radic. Biol. Med. 2018, 126, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, S.; Jung, S.; Müller, R.; Lademann, J.; Zuberbier, T.; Zastrow, L.; Reble, C.; Beckers, I.; Meinke, M.C. Skin type differences in solar-simulated radiation-induced oxidative stress. Br. J. Dermatol. 2019, 180, 597–603. [Google Scholar] [CrossRef]
- Aboul-Enein, H.Y.; Kladna, A.; Kruk, I.; Lichszteld, K.; Michalska, T. Effect of psoralens on Fenton-like reaction generating reactive oxygen species. Biopolym. Biospectroscopy 2003, 72, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Collet, M.; Hoebeke, M.; Piette, J.; Jakobs, A.; Lindqvist, L.; Van de Vorst, A. Photosensitized generation of hydroxyl radical by eight new sulfur and selenium analogs of psoralen. J. Photochem. Photobiol. B Biol. 1996, 35, 221–231. [Google Scholar] [CrossRef]
- Knox, C.N.; Land, E.J.; Truscott, T.G. Singlet oxygen generation by furocoumarins triplet states—1. Linear furocoumarins (Psoralens). Photochem. Photobiol. 1986, 43, 359–363. [Google Scholar] [CrossRef]
- Craw, M.; Chedekal, M.R.; Truscott, T.G.; Land, E.J. The photochemical interaction between the triplet state of 8-methoxypsoralen and the melanin precursor L-3,4 dihydroxyphenylalanine. Photochem. Photobiol. 1984, 39, 155–159. [Google Scholar] [CrossRef]
- Bacci, M.; Linari, R. UV excitable fluorescence of lumirubin. J. Photochem. Photobiol. B Biol. 1989, 3, 419–427. [Google Scholar] [CrossRef]
- Jasprova, J.; Dal Ben, M.; Vianello, E.; Goncharova, I.; Urbanova, M.; Vyroubalova, K.; Gazzin, S.; Tiribelli, C.; Sticha, M.; Cerna, M.; et al. The biological effects of bilirubin photoisomers. PLoS ONE 2016, 11, e0148126. [Google Scholar] [CrossRef]
- Böhm, F.; Drygalla, F.; Charlesworth, P.; Böhm, K.; Truscott, T.G.; Jokiel, K. Bilirubin phototoxicity to human cells by green light phototherapy in vitro. Photochem. Photobiol. 1995, 62, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, A.; Pospíšilová, K.; Žížalová, K.; Capková, N.; Muchová, L.; Vecka, M.; Vrzáčková, N.; Křížová, J.; Zelenka, J.; Vítek, L. The effects of bilirubin and lumirubin on metabolic and oxidative stress markers. Front. Pharmacol. 2021, 12, 567001. [Google Scholar] [CrossRef]
- Lowe, N.J.; Shaath, N.A. Sunscreens: Development, Evaluation and Regulatory Aspects; Marcel Dekker: New York, NY, USA, 1990. [Google Scholar]
- Trivedi, M.; Murase, J. Titanium dioxide in sunscreen. In Application of Titanium Dioxide; Janus, M., Ed.; IntechOpen: London, UK, 2017; Available online: https://www.intechopen.com/chapters/55103 (accessed on 10 August 2021).
- Wu, M.J.; Bak, T.; O’Doherty, P.J.; Moffitt, M.C.; Nowotny, J.; Bailey, T.D.; Kersaitis, C. Photocatalysis of titanium dioxide for water disinfection: Challenges and future perspectives. Int. J. Photochem. 2014, 2014, 973484. [Google Scholar] [CrossRef]
- Newman, M.D.; Stotland, M.; Ellis, J.I. The safety of nanosized particles in titanium dioxide- and zinc oxide-based sunscreens. J. Am. Acad. Dermatol. 2009, 61, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Serpone, N.; Salinaro, A.; Emeline, A. Deleterious effects of sunscreen titanium dioxide nanoparticles on DNA: Efforts to limit DNA damage by particle surface modification. In Proceedings of the BiOS 2001 the International Symposium on Biomedical Optics, San Jose, CA, USA, 20 January 2001; Volume 4258, pp. 86–98. [Google Scholar]
- Hidaka, H.; Kobayashi, H.; Koike, T.; Serpone, N. DNA damage photoinduced by cosmetic pigments and sunscreen agents under solar exposure and artificial UV illumination. J. Oleo Sci. 2006, 55, 249–261. [Google Scholar] [CrossRef][Green Version]
- Knowland, J.; McKenzie, E.A.; McHugh, P.J.; Cridland, N.A. Sunlight-induced mutagenicity of a common sunscreen ingredient. FEBS Lett. 1993, 324, 309–313. [Google Scholar] [CrossRef]
- Mturi, G.J.; Martincigh, B. Photostability of the sunscreening agent 4- tert-butyl-4′-methoxydibenzoylmethane (avobenzone) in solvents of different polarity and proticity. J. Photochem. Photobiol. A Chem. 2008, 200, 410–420. [Google Scholar] [CrossRef]
- Andrae, I.; Bringhen, A.; Böhm, F.; Gonzenbach, H.; Hill, T.; Mulroy, L.; Truscott, T.G. A UVA filter (4-tert-butyl-4′-methoxydibenzoylmethane): Photoprotection reflects photophysical properties. J. Photochem. Photobiol. B Biol. 1997, 37, 147–150. [Google Scholar] [CrossRef]
- Cantrell, A.; McGarvey, D.J. Photochemical studies of 4-tert-butyl-4′-methoxydibenzoylmethane (BM-DBM). J. Photochem. Photobiol. B Biol. 2001, 64, 117–122. [Google Scholar] [CrossRef]
- Hill, T.J. Molecular Mechanisms of Photoprotection. Ph.D. Thesis, Keele University, Keele, UK, 1994. [Google Scholar]
- Paris, C.; Lhiaubet-Vallet, V.; Jimenez, O.; Trullas, C.; Miranda, M.A. A blocked diketo form of avobenzone: Photostability, photosensitizing properties and triplet quenching by a triazine-derived UVB-filter. Photochem. Photobiol. 2009, 85, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Lhiaubet-Vallet, V.; Marin, M.; Jimenez, O.; Gorchs, O.; Trullas, C.; Miranda, M.A. Filter–filter interactions. Photostabilization, triplet quenching and reactivity with singlet oxygen. Photochem. Photobiol. Sci. 2010, 9, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Berenbeim, J.A.; Wong, N.G.K.; Cockett, M.C.R.; Berden, G.; Oomens, J.; Rijs, A.M.; Dessent, C.E.H. Unravelling the keto–enol tautomer dependent photochemistry and degradation pathways of the protonated UVA filter avobenzone. J. Phys. Chem. A 2020, 124, 2919–2930. [Google Scholar] [CrossRef]
- Damiani, E.; Rosati, L.; Castagna, R.; Carloni, P.; Greci, L. Changes in ultraviolet absorbance and hence in protective efficacy against lipid peroxidation of organic sunscreens after UVA irradiation. J. Photochem. Photobiol. B Biol. 2006, 82, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, A.; Oguchi, N.; Yagi, M. Optical and electron paramagnetic resonance studies of the excited states of 4-tert-butyl-4′-methoxydibenzoylmethane and 4-tert-butyl-4′-methoxydibenzoylpropane. J. Phys. Chem. A 2009, 113, 13492–13497. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, T.; Pan, J.; Scheurich, R.; Pfluecker, F.; Graf, R.; Epstein, H. Superior two step approach to completely photoprotect avobenzone with a designed organic redox pair. SOFW J. 2009, 135, 14–18. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).