
Video Transect-Based Coral Demographic Investigation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Survey and Data Extractions
2.2. Data Analysis
3. Results
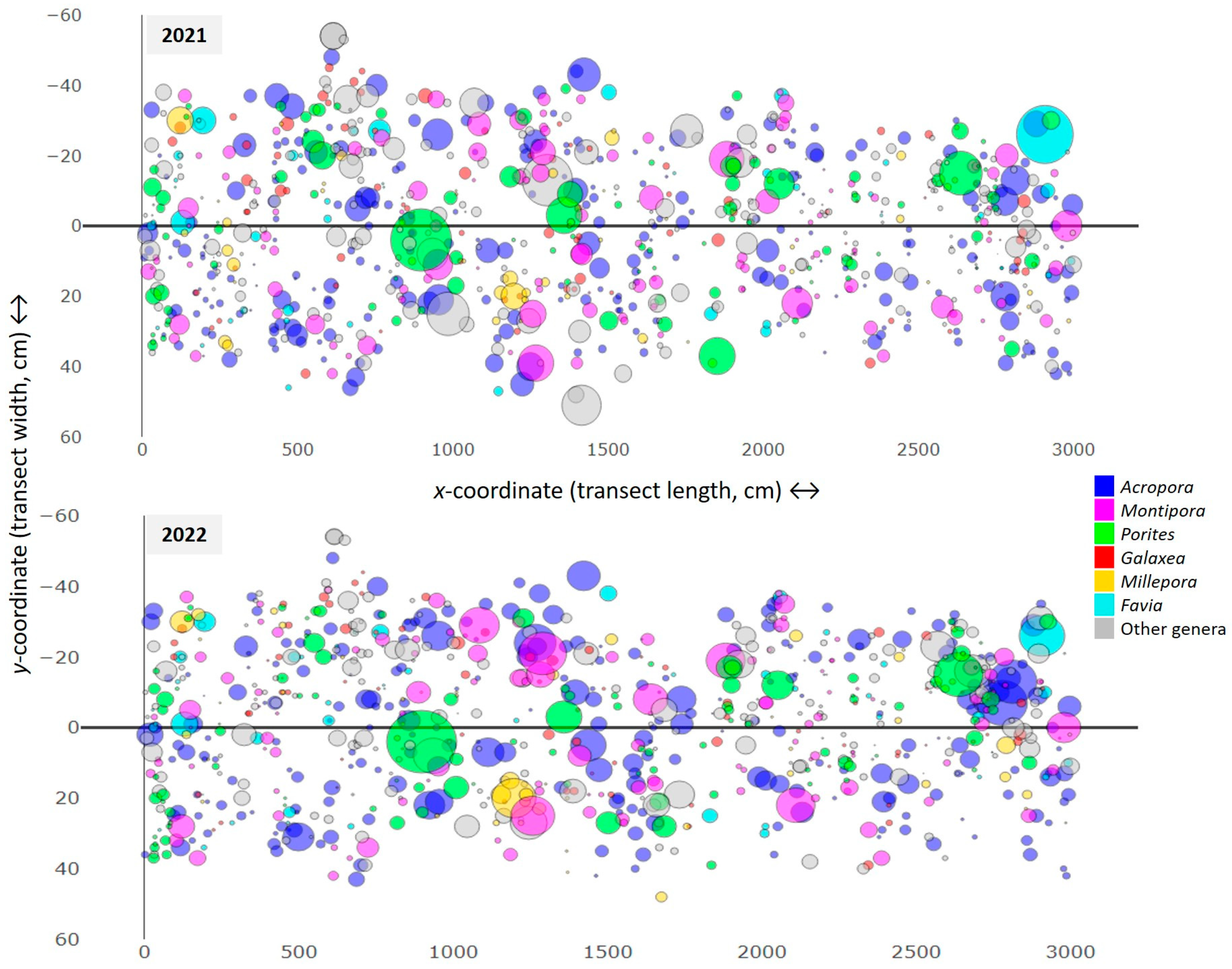
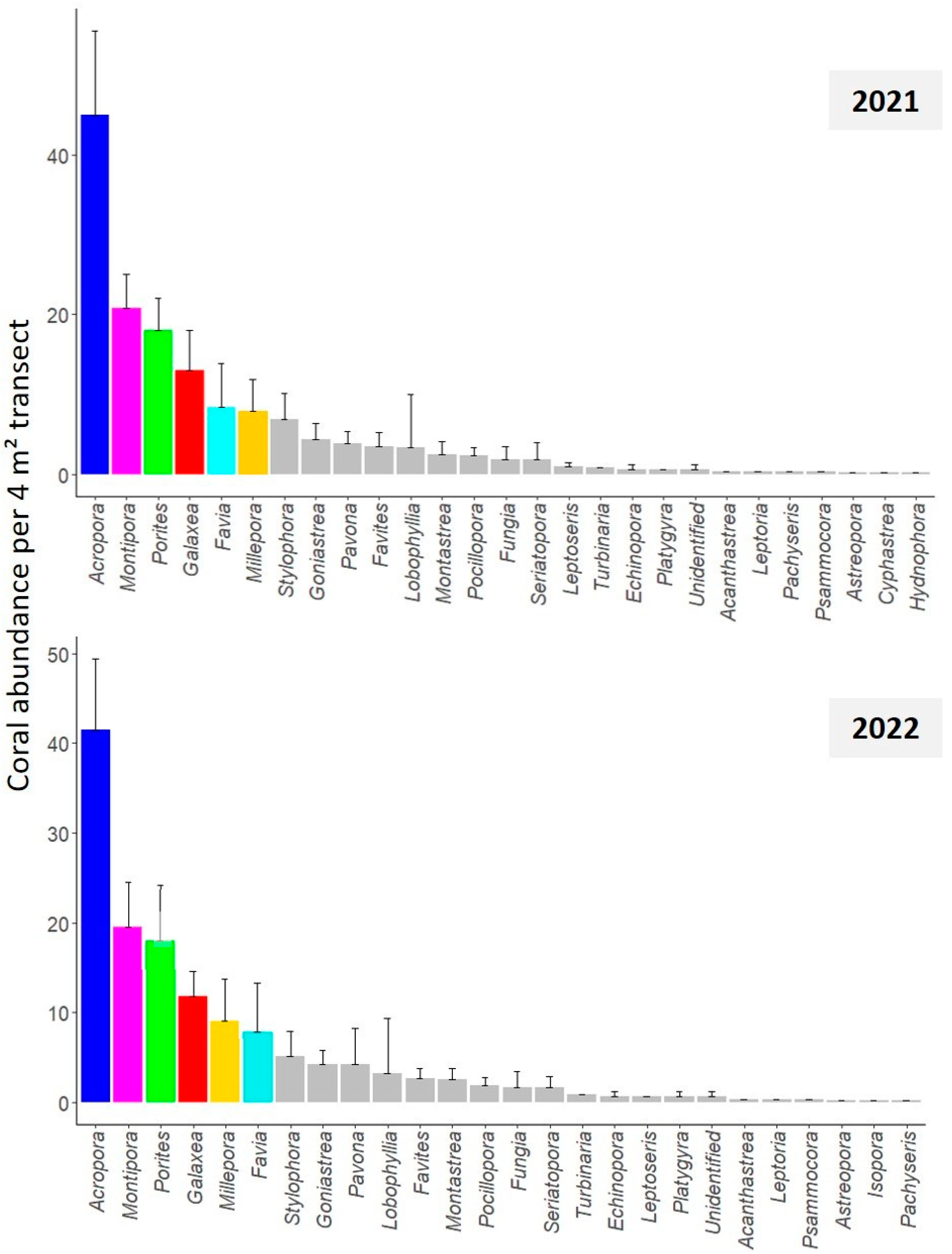
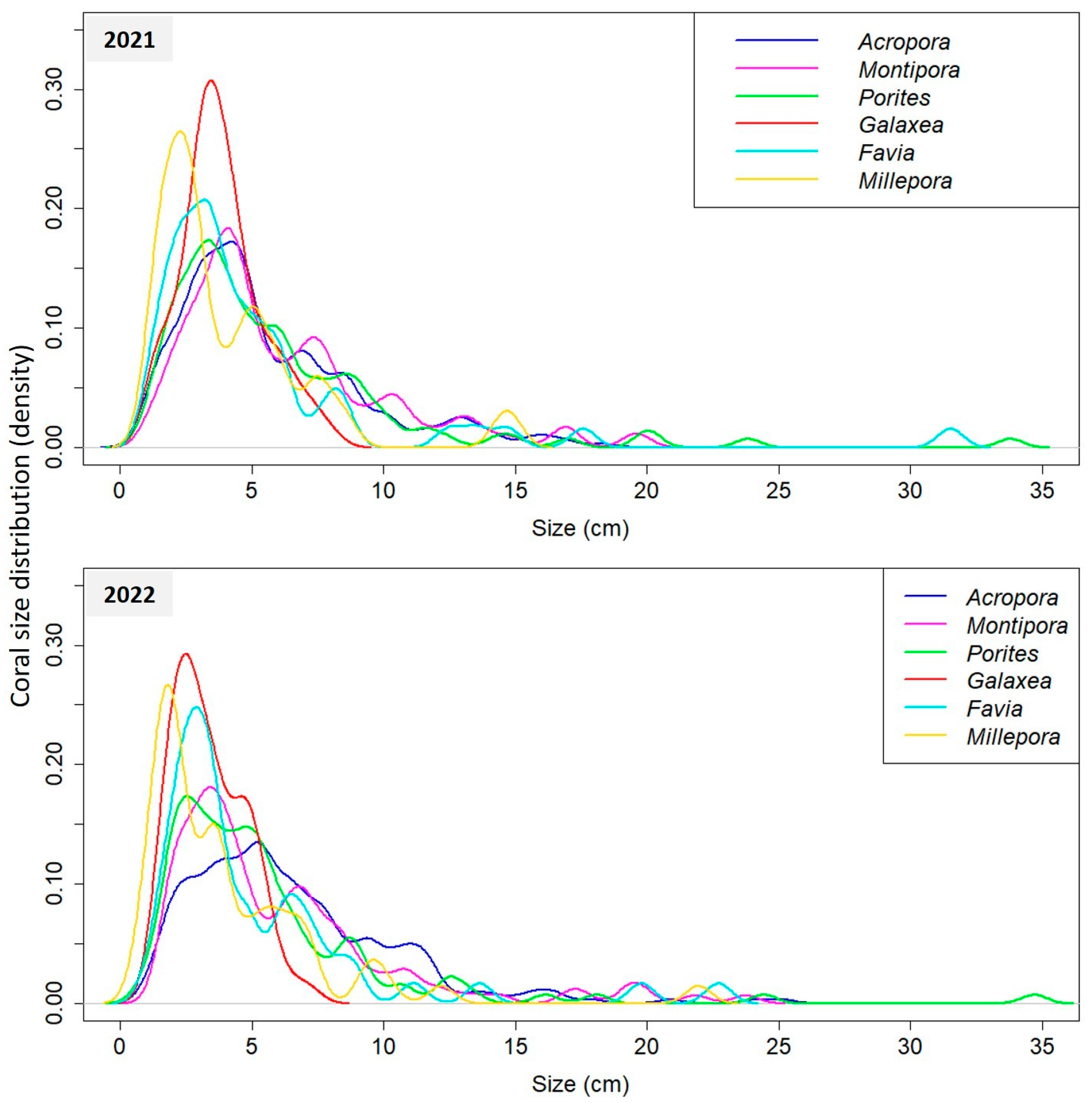
3.1. Coral Community Structure
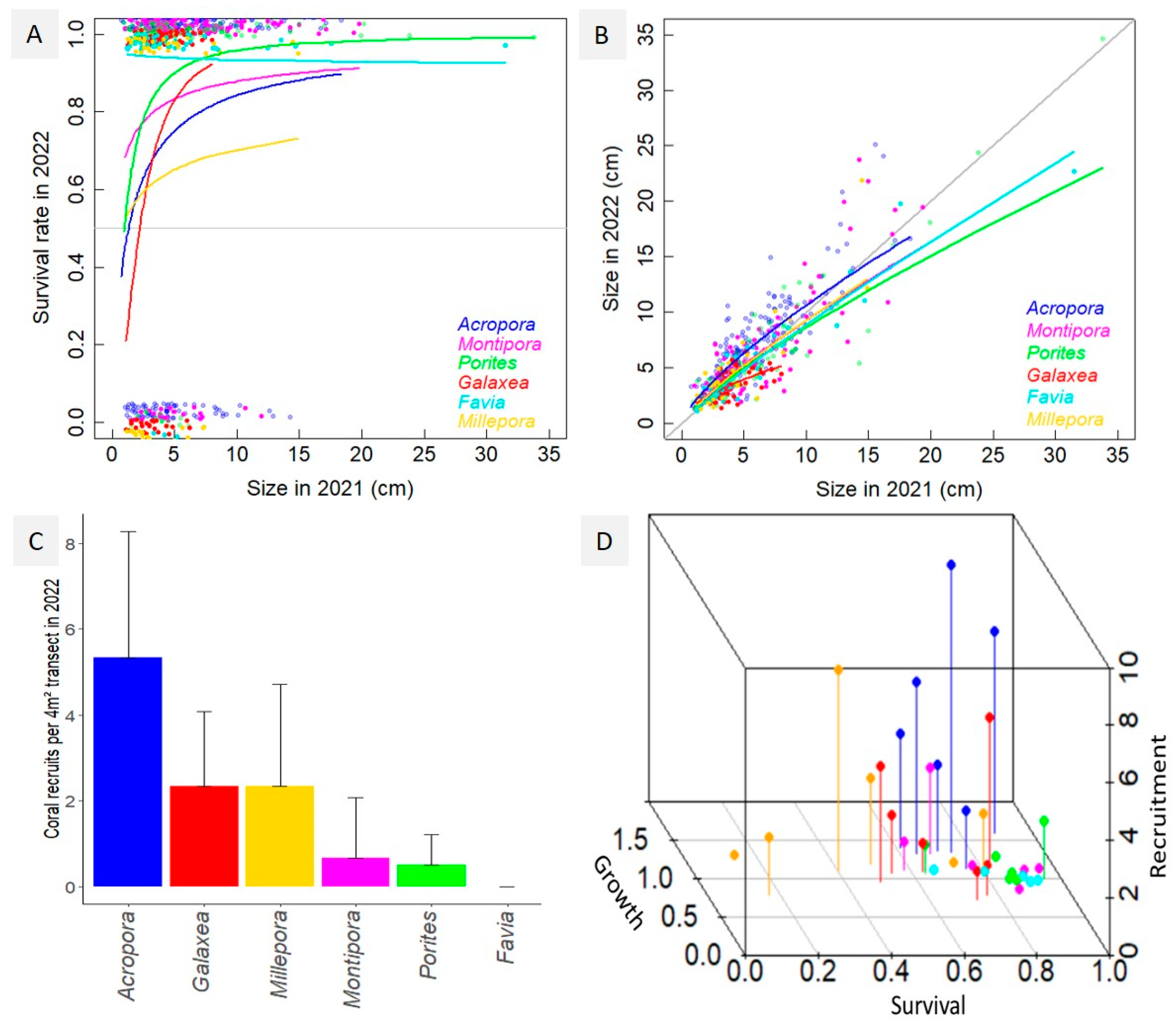
3.2. Coral Demographic Performances
4. Discussion
4.1. Characterizing Coral Demographic Performances
4.2. Accelerating Demographic Investigation Using Imagery
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halpern, B.S.; Frazier, M.; Afflerbach, J.; Lowndes, J.S.; Micheli, F.; O’Hara, C.; Scarborough, C.; Selkoe, K.A. Recent pace of change in human impact on the world’s ocean. Sci. Rep. 2019, 9, 11609. [Google Scholar] [CrossRef]
- Weiskopf, S.R.; Rubenstein, M.A.; Crozier, L.G.; Gaichas, S.; Griffis, R.; Halofsky, J.E.; Hyde, K.J.W.; Morelli, T.L.; Morisette, J.T.; Muñoz, R.C.; et al. Climate change effects on biodiversity, ecosystems, ecosystem services, and natural resource management in the United States. Sci. Total Environ. 2020, 733, 137782. [Google Scholar] [CrossRef]
- Condie, S.A.; Anthony, K.R.; Babcock, R.C.; Baird, M.E.; Beeden, R.; Fletcher, C.S.; Gorton, R.; Harrison, D.; Hobday, A.J.; Plagányi, E.E.; et al. Large-scale interventions may delay decline of the Great Barrier Reef. R. Soc. Open Sci. 2021, 8, 201296. [Google Scholar] [CrossRef]
- Andrello, M.; Darling, E.S.; Wenger, A.; Suárez-Castro, A.F.; Gelfand, S.; Ahmadia, G.N. A global map of human pressures on tropical coral reefs. Conserv. Lett. 2022, 15, e12858. [Google Scholar] [CrossRef]
- Ellner, S.P.; Childs, D.Z.; Rees, M. Data-Driven Modelling of Structured Populations: A Practical Guide to the Integral Projection Model; Springer International Publishing: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Dumas, P.; Bertaud, A.; Peignon, C.; Leopold, M.; Pelletier, D.A. “Quick and clean” photographic method for the description of coral reef habitats. J. Exp. Mar. Biol. Ecol. 2009, 368, 161–168. [Google Scholar] [CrossRef]
- Heudier, M.; Mouillot, D.; Mannocci, L. Assessing the effects of coral reef habitat and marine protected areas on threatened megafauna using aerial surveys. Aquat. Conserv. Mar. Freshw. 2023, 33, 286–297. [Google Scholar] [CrossRef]
- González-Rivero, M.; Beijbom, O.; Rodriguez-Ramirez, A.; Bryant, D.E.P.; Ganase, A.; Gonzalez-Marrero, Y.; Kennedy, E.V.; Kim, C.J.S.; Lopez-Marcano, S.; Markey, K.; et al. Monitoring of coral reefs using artificial intelligence: A feasible and cost-effective approach. Remote Sens. 2020, 12, 489. [Google Scholar] [CrossRef]
- Urbina-Barreto, I.; Chiroleu, F.; Pinel, R.; Fréchon, L.; Mahamadaly, V.; Elise, S.; Kulbicki, M.; Quod, J.-P.; Dutrieux, E.; Garnier, R.; et al. Quantifying the shelter capacity of coral reefs using photogrammetric 3D modeling: From colonies to reefscapes. Ecol. Indic. 2021, 121, 107151. [Google Scholar] [CrossRef]
- Donovan, M.K.; Alves, C.; Burns, J.; Drury, C.; Meier, O.W.; Ritson-Williams, R.; Cunning, R.; Dunn, R.P.; Goodbody-Gringley, G.; Henderson, L.M.; et al. From polyps to pixels: Understanding coral reef resilience to local and global change across scales. Landsc. Ecol. 2023, 38, 737–752. [Google Scholar] [CrossRef]
- Kayal, M.; Vercelloni, J.; Wand, M.P.; Adjeroud, M. Searching for the best bet in life-strategy: A quantitative approach to individual performance and population dynamics in reef-building corals. Ecol. Complex. 2015, 23, 73–84. [Google Scholar] [CrossRef]
- Darling, E.S.; McClanahan, T.R.; Maina, J.; Gurney, G.G.; Graham, N.A.J.; Januchowski-Hartley, F.; Cinner, J.E.; Mora, C.; Hicks, C.C.; Maire, E.; et al. Social-environmental drivers inform strategic management of coral reefs in the Anthropocene. Nat. Ecol. Evol. 2019, 3, 1341–1350. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock–recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Pendleton, L.; Kaup, A. People and the changing nature of coral reefs. Reg. Stud. Mar. Sci. 2019, 30, 100699. [Google Scholar] [CrossRef]
- Eddy, T.D.; Lam, V.W.; Reygondeau, G.; Cisneros-Montemayor, A.M.; Greer, K.; Palomares, M.L.D.; Bruno, J.F.; Ota, Y.; Cheung, W.W.L. Global decline in capacity of coral reefs to provide ecosystem services. One Earth 2021, 4, 1278–1285. [Google Scholar] [CrossRef]
- Carlot, J.; Vousdoukas, M.; Rovere, A.; Karambas, T.; Lenihan, H.S.; Kayal, M.; Adjeroud, M.; Pérez-Rosales, G.; Hedouin, L.; Parravicini, V. Coral reef structural complexity loss exposes coastlines to waves. Sci. Rep. 2023, 13, 1683. [Google Scholar] [CrossRef] [PubMed]
- Kayal, M.; Lenihan, H.S.; Brooks, A.J.; Holbrook, S.J.; Schmitt, R.J.; Kendall, B.E. Predicting coral community recovery using multi-species population dynamics models. Ecol. Lett. 2018, 21, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Madin, J.S.; Baird, A.H.; Dornelas, M.; Connolly, S.R. Mechanical vulnerability explains size-dependent mortality of reef corals. Ecol. Lett. 2014, 17, 1008–1015. [Google Scholar] [CrossRef]
- Riegl, B.; Johnston, M.; Purkis, S.; Howells, E.; Burt, J.; Steiner, S.C.C.; Sheppard, C.R.C.; Bauman, A. Population collapse dynamics in Acropora downingi, an Arabian/Persian Gulf ecosystem-engineering coral, linked to rising temperature. Glob. Chang. Biol. 2018, 24, 2447–2462. [Google Scholar] [CrossRef] [PubMed]
- Carlot, J.; Kayal, M.; Lenihan, H.S.; Brandl, S.J.; Casey, J.M.; Adjeroud, M.; Cardini, U.; Mercière, A.; Espiau, B.; Barneche, D.R.; et al. Juvenile corals underpin coral reef carbonate production after disturbance. Glob. Chang. Biol. 2021, 27, 2623–2632. [Google Scholar] [CrossRef]
- Fenner, D.; Muir, P. Reef Corals of the Northwestern Lagoon of Grande-Terre, New Caledonia. In A Rapid Marine Biodiversity Assessment of the Coral Reefs of the Northwest Lagoon, between Koumac and Yandé, Province Nord, New Caledonia; Conservation International: Arlington, TX, USA, 2008. [Google Scholar] [CrossRef]
- Andréfouët, S.; Cabioch, G.; Flamand, B.; Pelletier, B. A reappraisal of the diversity of geomorphological and genetic processes of New Caledonian coral reefs: A synthesis from optical remote sensing, coring and acoustic multibeam observations. Coral Reefs 2009, 28, 691–707. [Google Scholar] [CrossRef]
- Adjeroud, M.; Poisson, E.; Peignon, C.; Penin, L.; Kayal, M. Spatial patterns and short-term changes of coral assemblages along a cross-shelf gradient in the southwestern lagoon of New Caledonia. Diversity 2019, 11, 21. [Google Scholar] [CrossRef]
- Kayal, M.; Adjeroud, M. The war of corals: Patterns, drivers, and implications of changing coral competitive performances across reef environments. R. Soc. Open Sci. 2022, 9, 220003. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models, R Package Version 3.1-162. 2023. Available online: https://CRAN.R-project.org/package=nlme (accessed on 14 February 2023).
- Adjeroud, M.; Kayal, M.; Iborra-Cantonnet, C.; Vercelloni, J.; Bosserelle, P.; Liao, V.; Chancerelle, Y.; Claudet, J.; Penin, L. Recovery of coral assemblages despite acute and recurrent disturbances on a South Central Pacific reef. Sci. Rep. 2018, 8, 9680. [Google Scholar] [CrossRef] [PubMed]
- Dietzel, A.; Bode, M.; Connolly, S.R.; Hughes, T.P. The population sizes and global extinction risk of reef-building coral species at biogeographic scales. Nat. Ecol. Evol. 2021, 5, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Tokeshi, M. Species Coexistence: Ecological and Evolutionary Perspectives; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Holt, R.D. Species Coexistence. In Reference Module in Life Sciences; Elsevier: Amsterdam, Netherlands, 2017. [Google Scholar] [CrossRef]
- Adjeroud, M.; Peignon, C.; Gauliard, C.; Penin, L.; Kayal, M. Extremely high but localized pulses of coral recruitment in the southwestern lagoon of New Caledonia and implications for conservation. Mar. Ecol. Prog. Ser. 2022, 692, 67–79. [Google Scholar] [CrossRef]
- Dubé, C.E.; Ziegler, M.; Mercière, A.; Boissin, E.; Planes, S.; Bourmaud, C.F.; Voolstra, C.R. Naturally occurring fire coral clones demonstrate a genetic and environmental basis of microbiome composition. Nat. Commun. 2021, 12, 6402. [Google Scholar] [CrossRef]
- Jamil, S.; Rahman, M.; Haider, A. Bag of features (BoF) based deep learning framework for bleached corals detection. Big Data Cogn. Comput. 2021, 5, 53. [Google Scholar] [CrossRef]
- Dietzel, A.; Bode, M.; Connolly, S.R.; Hughes, T.P. Long-term shifts in the colony size structure of coral populations along the Great Barrier Reef. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201432. [Google Scholar] [CrossRef] [PubMed]
- Track Changes: Coastal Marine Ecosystem Transformation in the Face of Environmental Changes. Available online: https://umr-entropie.ird.nc/index.php/home-en/home/research/portfolio/ongoing-projects/projet-track-changes (accessed on 14 February 2023).



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayal, M.; Mevrel, E.; Ballard, J. Video Transect-Based Coral Demographic Investigation. Coasts 2023, 3, 370-382. https://doi.org/10.3390/coasts3040022
Kayal M, Mevrel E, Ballard J. Video Transect-Based Coral Demographic Investigation. Coasts. 2023; 3(4):370-382. https://doi.org/10.3390/coasts3040022
Chicago/Turabian StyleKayal, Mohsen, Eva Mevrel, and Jane Ballard. 2023. "Video Transect-Based Coral Demographic Investigation" Coasts 3, no. 4: 370-382. https://doi.org/10.3390/coasts3040022
APA StyleKayal, M., Mevrel, E., & Ballard, J. (2023). Video Transect-Based Coral Demographic Investigation. Coasts, 3(4), 370-382. https://doi.org/10.3390/coasts3040022