
Scale-Up Cultivation of the Dinoflagellate Durusdinium glynnii Under Varying Inoculum Percentages: Effects on Growth Performance and Fatty Acid Profile

 , ,
, ,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Scaling-Up Strategies
- (i)
- Test tubes with 50 mL of working volume;
- (ii)
- Erlenmeyer flasks with working volume ranging from 0.25 L to 2 L;
- (iii)
- 5 L transparent plastic bottles;
- (iv)
- A cylindrical pilot-scale photobioreactor, adapted from a 100 L carboy tank.
2.3. Growth Analysis
2.4. Biomass Harvesting and Drying
2.5. Lipid Extraction and Fatty Acid Composition
2.6. Statistical Analysis
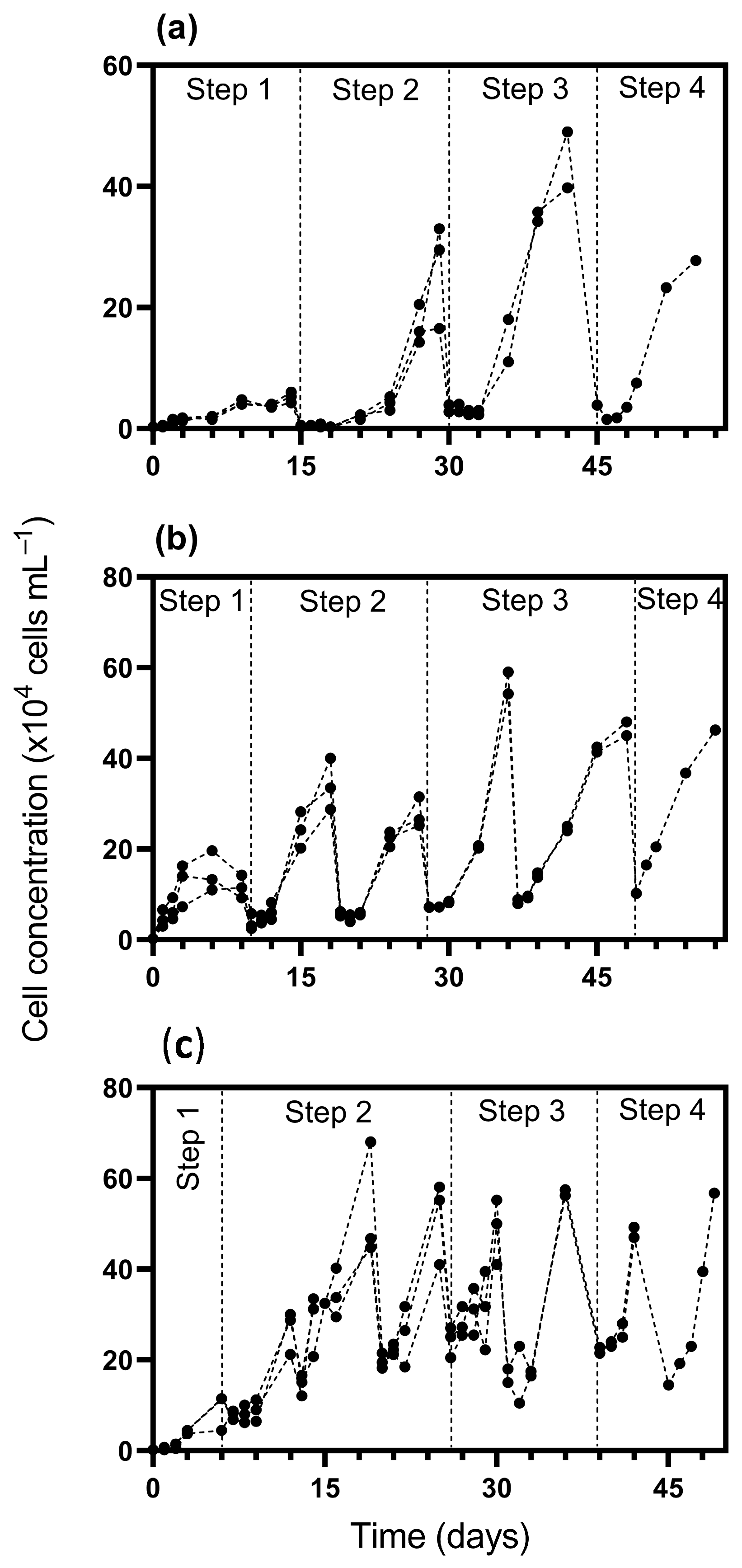
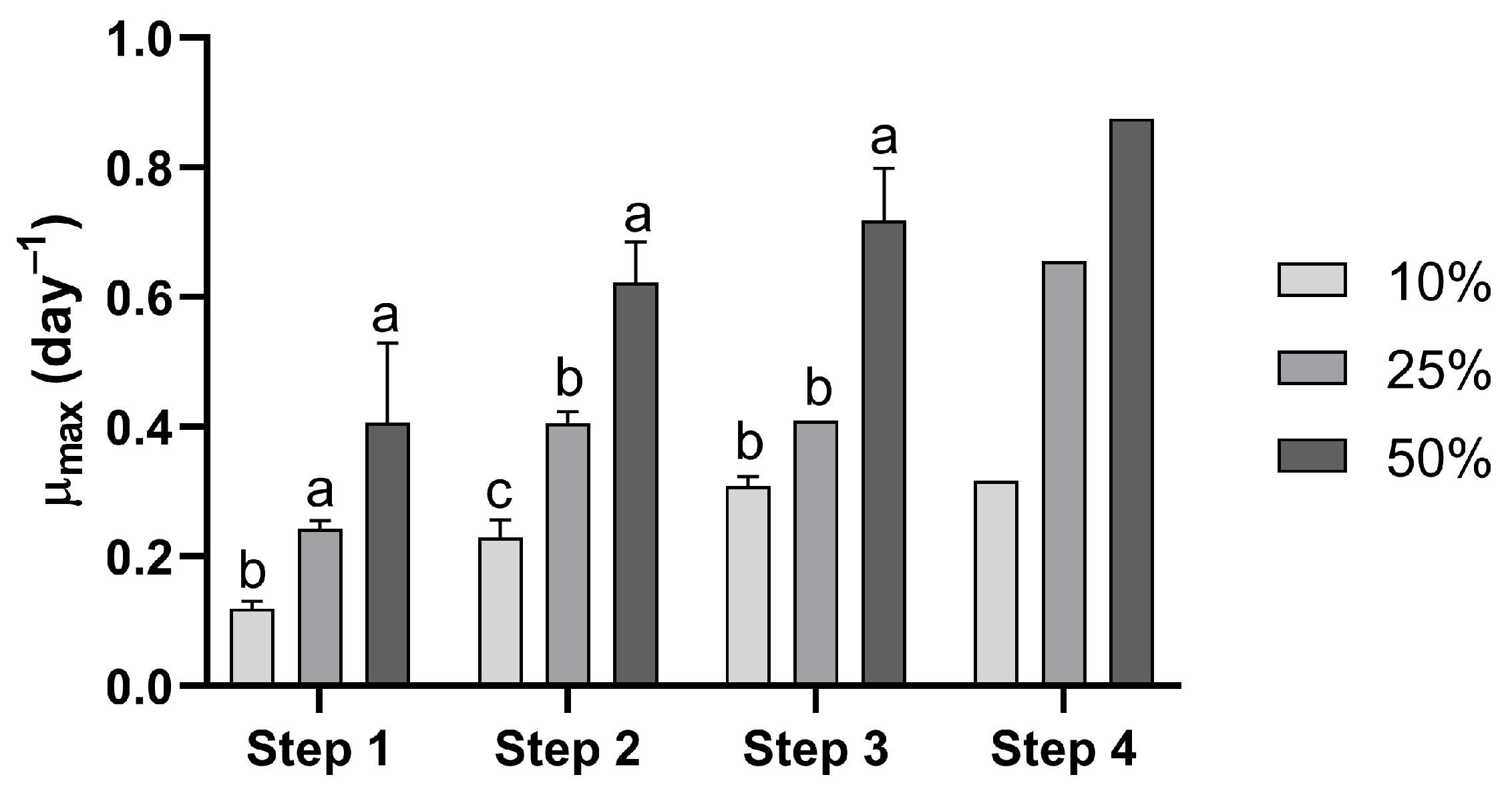
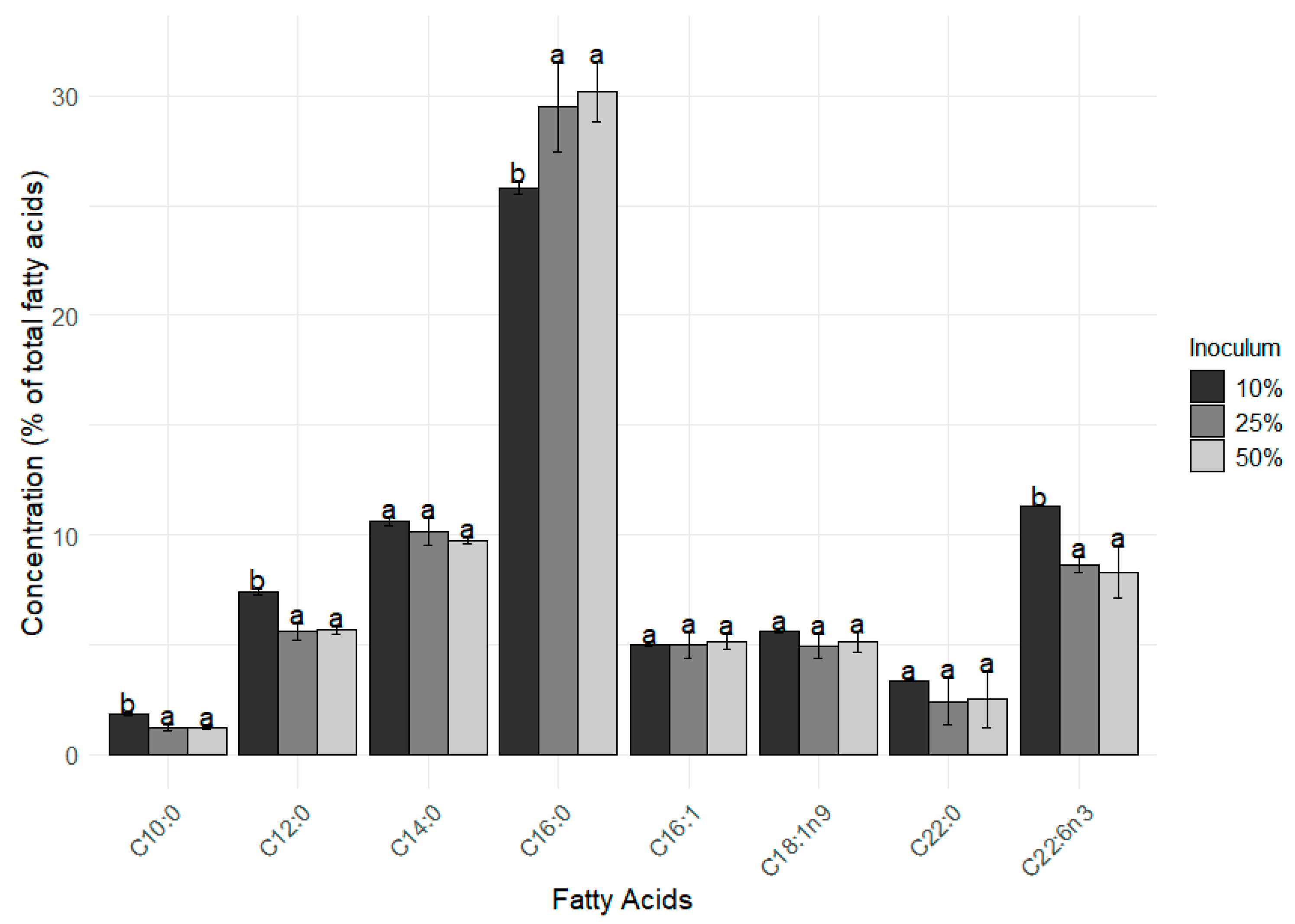
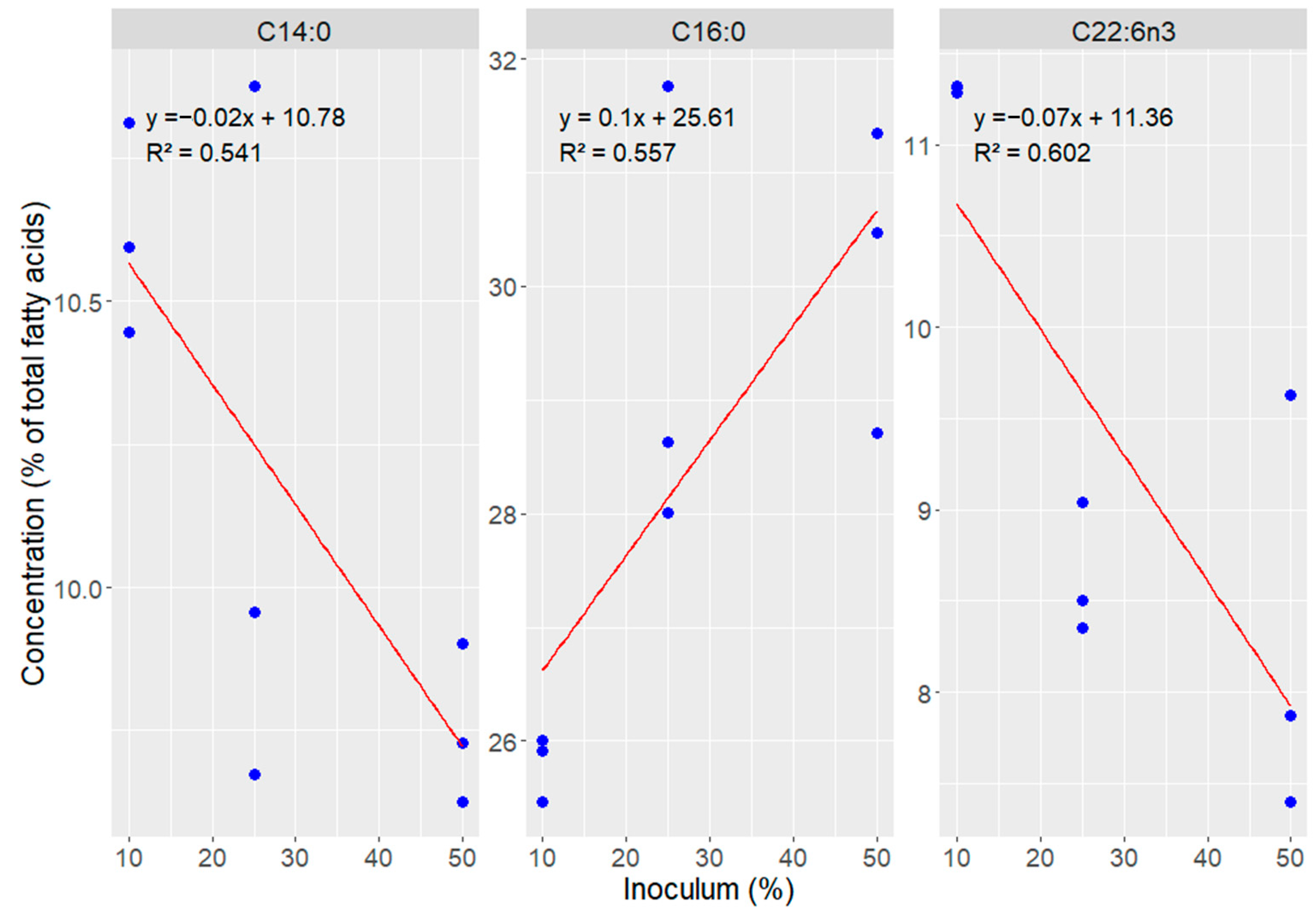
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guiry, M.D. How Many Species of Algae Are There? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Sreeharsha, R.V.; Mohan, S.V. Microbial Photosynthesis. In Photosynthetic Microbes: Evolution, Classification, and Structural Physiology; Springer: Singapore, 2024; pp. 3–22. [Google Scholar] [CrossRef]
- Mordret, S.; Romac, S.; Henry, N.; Colin, S.; Carmichael, M.; Berney, C.; Audic, S.; Richter, D.J.; Pochon, X.; de Vargas, C.; et al. The Symbiotic Life of Symbiodinium in the Open Ocean Within a New Species of Calcifying Ciliate (Tiarina sp.). ISME J. 2015, 10, 1424–1436. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Garcia, L.; Adjallé, K.; Barnabé, S.; Raghavan, G.S.V. Microalgae Biomass Production for a Biorefinery System: Recent Advances and the Way Towards Sustainability. Renew. Sustain. Energy Rev. 2017, 76, 493–506. [Google Scholar] [CrossRef]
- Oliveira, A.P.F.; Bragotto, A.P.A. Microalgae-Based Products: Food and Public Health. Future Foods 2022, 6, 100157. [Google Scholar] [CrossRef]
- Rizwan, M.; Mujtaba, G.; Memon, S.A.; Lee, K.; Rashid, N. Exploring the Potential of Microalgae for New Biotechnology Applications and Beyond: A Review. Renew. Sustain. Energy Rev. 2018, 92, 394–404. [Google Scholar] [CrossRef]
- Morse, D.; Tse, S.P.K.; Lo, S.C.L. Exploring Dinoflagellate Biology with High-Throughput Proteomics. Harmful Algae 2018, 75, 16–26. [Google Scholar] [CrossRef]
- Lee, S.K.; Jeong, H.J.; Jang, S.H.; Lee, K.H.; Kang, N.S.; Lee, M.J.; Potvin, E. Mixotrophy in the Newly Described Dinoflagellate Ansanella granifera: Feeding Mechanism, Prey Species, and Effect of Prey Concentration. Algae 2014, 29, 137–152. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jeong, H.J.; Kwon, J.E.; You, J.H.; Kim, S.J.; Ok, J.H.; Kang, H.C.; Park, J.Y. First Report of the Photosynthetic Dinoflagellate Heterocapsa minima in the Pacific Ocean: Morphological and Genetic Characterizations and the Nationwide Distribution in Korea. Algae 2019, 34, 7–21. [Google Scholar] [CrossRef]
- Nitschke, M.R.; Rosset, S.L.; Oakley, C.A.; Gardner, S.G.; Camp, E.F.; Suggett, D.J.; Davy, S.K. The Diversity and Ecology of Symbiodiniaceae: A Traits-Based Review. Adv. Mar. Biol. 2022, 92, 55–127. [Google Scholar]
- Camacho, F.G.; Rodríguez, J.G.; Mirón, A.S.; García, M.C.; Belarbi, E.H.; Chisti, Y.; Grima, E.M. Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Oliveira, C.D.L.; Müller, M.N.; Santos, E.P.; Dantas, D.M.M.; Gálvez, A.O. A Scientometric Overview of Global Dinoflagellate Research. Publications 2020, 8, 50. [Google Scholar] [CrossRef]
- Assunção, J.; Guedes, A.C.; Malcata, F.X. Biotechnological and Pharmacological Applications of Biotoxins and Other Bioactive Molecules from Dinoflagellates. Mar. Drugs 2017, 15, 393. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.H.; Gibson, C.H. Quantified Small-Scale Turbulence Inhibits a Red Tide Dinoflagellate, Gonyaulax polyedra Stein. Deep Sea Res. I Oceangr. Res. Pap. 1990, 37, 1583–1593. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.J.G.; Mirón, A.S.; Belarbi, E.H.; Chisti, Y.; Grima, E.M. Photobioreactor Scale-Up for a Shear-Sensitive Dinoflagellate Microalga. Process Biochem. 2011, 46, 936–944. [Google Scholar] [CrossRef]
- Greenhough, H.; Waugh, C.; van Ginkel, R.; Bowater, J.; Kaur, G.; Oakly, J.; Plouviez, M.; Ingebrigtsen, R.A.; Svenson, J.; Selwood, A.I. Mass Cultivation of the Dinoflagellate Alexandrium pacificum for Gonyautoxin-1,4 Production. Sci. Rep. 2025, 15, 7430. [Google Scholar] [CrossRef]
- De Cassia, S.; Brandão, B.; de Abreu, J.L.; Oliveira, D.W.S.; da Silva Campos, C.V.F.; de Aguiar, I.M.T.; de Sena, P.R.; Gálvez, A.O.; Oliveira, C.Y.B. New Findings on the Survival of Durusdinium glynnii Under Different Acclimation Methods to Low Salinities. Microorganisms 2025, 13, 946. [Google Scholar] [CrossRef]
- Molina-Miras, A.; Bueso-Sánchez, A.; Cerón-García, M.C.; Sánchez-Mirón, A.; Contreras-Gómez, A.; García-Camacho, F. Effect of Nitrogen, Phosphorus, and Light Colimitation on Amphidinol Production and Growth in the Marine Dinoflagellate Microalga Amphidinium carterae. Toxins 2022, 14, 594. [Google Scholar] [CrossRef]
- Rathinavel, L.; Ravikumar, S.M.; Jothinathan, D.; Paul, S.J.; Pandey, A.; Mahata, C. Extraction and Enrichment of Fatty Acids from Marine Microalgae. In Marine Molecules from Algae and Cyanobacteria: Extraction, Purification, Toxicology and Applications; Elsevier: Amsterdam, The Netherlands, 2025; Chapter 3; pp. 41–57. [Google Scholar] [CrossRef]
- Remize, M.; Brunel, Y.; Silva, J.L.; Berthon, J.-Y.; Filaire, E. Microalgae n-3 PUFAs Production and Use in Food and Feed Industries. Mar. Drugs 2021, 19, 113. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Jacob, A.; Nader, C.; Oliveira, C.D.L.; Matos, Â.P.; Araújo, E.S.; Shabnam, N.; Ashok, B.; Gálvez, A.O. An Overview on Microalgae as Renewable Resources for Meeting Sustainable Development Goals. J. Environ. Manag. 2022, 320, 115854. [Google Scholar] [CrossRef]
- Gui, J.; Chen, S.; Luo, G.; Wu, Z.; Fan, Y.; Yao, L.; Xu, H. Nutrient Deficiency and an Algicidal Bacterium Improved the Lipid Profiles of a Novel Promising Oleaginous Dinoflagellate, Prorocentrum donghaiense, for Biodiesel Production. Appl. Environ. Microbiol. 2021, 87, e0115921. [Google Scholar] [CrossRef]
- Chen, W.; Li, T.; Du, S.; Chen, H.; Wang, Q. Microalgal Polyunsaturated Fatty Acids: Hotspots and Production Techniques. Front. Bioeng. Biotechnol. 2023, 11, 1146881. [Google Scholar] [CrossRef] [PubMed]
- Graeff, J.E.; Leblond, J.D. Composition of Galactolipids, Betaine Lipids and Triglyceride-Associated Fatty Acids of the Symbiotic Dinoflagellate Zooxanthella (Brandtodinium) Nutricula: A Glimpse into Polyunsaturated Fatty Acids Available to Its Polycystine Radiolarian Host. Phycol. Res. 2023, 71, 175–181. [Google Scholar] [CrossRef]
- Didrihsone, E.; Dubencovs, K.; Grube, M.; Shvirksts, K.; Suleiko, A.; Suleiko, A.; Vanags, J. Crypthecodinium cohnii Growth and Omega Fatty Acid Production in Mediums Supplemented with Extract from Recycled Biomass. Mar. Drugs 2022, 20, 68. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Lorenzen, C.J. Yellow-green algae with chlorophyllide c. J. Phycol. 1972, 8, 10–14. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Brandão, B.d.C.S.; Jannuzzi, L.G.d.S.; Oliveira, D.W.S.; Yogui, G.T.; Müller, M.N.; Gálvez, A.O. New insights on the role of nitrogen in the resistance to environmental stress in an endosymbiotic dinoflagellate. Environ. Sci. Pollut. Res. 2023, 30, 82142–82151. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Axelsson, M.; Gentili, F. A single-step method for rapid extraction of total lipids from green microalgae. PLoS ONE 2014, 9, e89643. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Abreu, J.L.; Santos, E.P.; Matos, Â.P.; Tribuzi, G.; Oliveira, C.D.L.; Veras, B.O.; Bezerra, R.S.; Müller, M.N.; Gálvez, A.O. Light induces peridinin and docosahexaenoic acid accumulation in the dinoflagellate Durusdinium glynnii. Appl. Microbiol. Biotechnol. 2022, 106, 6263–6276. [Google Scholar] [CrossRef]
- López-Rosales, L.; García-Camacho, F.; Sánchez-Mirón, A.; Martín Beato, E.; Chisti, Y.; Molina Grima, E. Pilot-scale bubble column photobioreactor culture of a marine dinoflagellate microalga illuminated with light emission diodes. Bioresour. Technol. 2016, 216, 845–855. [Google Scholar] [CrossRef]
- Alizade, A.; Jantschke, A. Dinoflagellates as sustainable cellulose source: Cultivation, extraction, and characterization. Int. J. Biol. Macromol. 2023, 242, 125116. [Google Scholar] [CrossRef] [PubMed]
- Vidyarathna, N.K.; Smith, L.E.; Miller, K.R.; Coyne, K.J.; Cohen, J.H.; Warner, M.E. Short-term and long-term exposure to combined elevated temperature and CO₂ leads to differential growth, toxicity, and fatty acid profiles in the harmful dinoflagellate Karlodinium veneficum. Front. Mar. Sci. 2024, 11, 1305495. [Google Scholar] [CrossRef]
- Aboualaalaa, H.; Leblad, B.R.; Elkbiach, M.L.; Ibghi, M.; Boutaib, R.; Maamour, N.; Savar, V.; Masseret, E.; Abadie, E.; Rolland, J.L.; et al. Effect of temperature, salinity and nutrients on the growth and toxin content of the dinoflagellate Gymnodinium catenatum from the southwestern Mediterranean. Sci. Total Environ. 2024, 945, 174094. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Wu, K.-C.; Chan, S.C.-Y.; Yau, Y.-H.; Chan, K.-K.; Lee, F.W.-F. Investigation of growth, lipid productivity, and fatty acid profiles in marine bloom-forming dinoflagellates as potential feedstock for biodiesel. J. Mar. Sci. Eng. 2020, 8, 381. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Y.; Song, S.; Li, C.; Tang, Y.Z.; Yu, Z. The effects of major environmental factors and nutrient limitation on growth and encystment of planktonic dinoflagellate Akashiwo sanguinea. Harmful Algae 2015, 46, 62–70. [Google Scholar] [CrossRef]
- Tsirigoti, A.; Tzovenis, I.; Koutsaviti, A.; Economou-Amilli, A.; Ioannou, E.; Melkonian, M. Biofilm cultivation of marine dinoflagellates under different temperatures and nitrogen regimes enhances DHA productivity. J. Appl. Phycol. 2020, 32, 865–880. [Google Scholar] [CrossRef]
- Ali, O.; Szabó, A. Review of eukaryote cellular membrane lipid composition, with special attention to the fatty acids. Int. J. Mol. Sci. 2023, 24, 15693. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K. Fatty acids of microalgae: Diversity and applications. Rev. Environ. Sci. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Sharma, T.; Das, N.; Kakkar, P.M.; Mohapatra, R.K.; Pamidimarri, S.; Singh, R.K.; Kumar, M.; Guldhe, A.; Nayak, M. Microalgae as an emerging alternative raw material of docosahexaenoic acid and eicosapentaenoic acid—A review. Crit. Rev. Food Sci. Nutr. 2025, 1–20. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sena, P.R.; Lira, M.E.S.S.; Oliveira, D.W.S.; Brandão, B.d.C.S.; Abreu, J.L.d.; Gama, W.A.; Araújo, E.S.; Tribuzi, G.; Gálvez, A.O.; Oliveira, C.Y.B. Scale-Up Cultivation of the Dinoflagellate Durusdinium glynnii Under Varying Inoculum Percentages: Effects on Growth Performance and Fatty Acid Profile. Phycology 2025, 5, 21. https://doi.org/10.3390/phycology5020021
de Sena PR, Lira MESS, Oliveira DWS, Brandão BdCS, Abreu JLd, Gama WA, Araújo ES, Tribuzi G, Gálvez AO, Oliveira CYB. Scale-Up Cultivation of the Dinoflagellate Durusdinium glynnii Under Varying Inoculum Percentages: Effects on Growth Performance and Fatty Acid Profile. Phycology. 2025; 5(2):21. https://doi.org/10.3390/phycology5020021
Chicago/Turabian Stylede Sena, Pedro Rodrigues, Maria Eunice S. S. Lira, Deyvid Willame S. Oliveira, Barbara de Cassia S. Brandão, Jessika L. de Abreu, Watson Arantes Gama, Evando S. Araújo, Giustino Tribuzi, Alfredo O. Gálvez, and Carlos Yure B. Oliveira. 2025. "Scale-Up Cultivation of the Dinoflagellate Durusdinium glynnii Under Varying Inoculum Percentages: Effects on Growth Performance and Fatty Acid Profile" Phycology 5, no. 2: 21. https://doi.org/10.3390/phycology5020021
APA Stylede Sena, P. R., Lira, M. E. S. S., Oliveira, D. W. S., Brandão, B. d. C. S., Abreu, J. L. d., Gama, W. A., Araújo, E. S., Tribuzi, G., Gálvez, A. O., & Oliveira, C. Y. B. (2025). Scale-Up Cultivation of the Dinoflagellate Durusdinium glynnii Under Varying Inoculum Percentages: Effects on Growth Performance and Fatty Acid Profile. Phycology, 5(2), 21. https://doi.org/10.3390/phycology5020021