
Fast-Tracking Isolation, Identification and Characterization of New Microalgae for Nutraceutical and Feed Applications



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Sample Collection and Isolation
2.2. Standardized Cultivation and Growth Experiments
2.3. DNA Isolation, Sequencing and Identification
2.4. Fatty Acid Methyl Ester (FAME) Analyses
2.5. Total Protein Analyses
2.6. Total Carotenoid Analyses
2.7. Statistical Analyses
3. Results
3.1. Sample Collection, Isolation and Identification
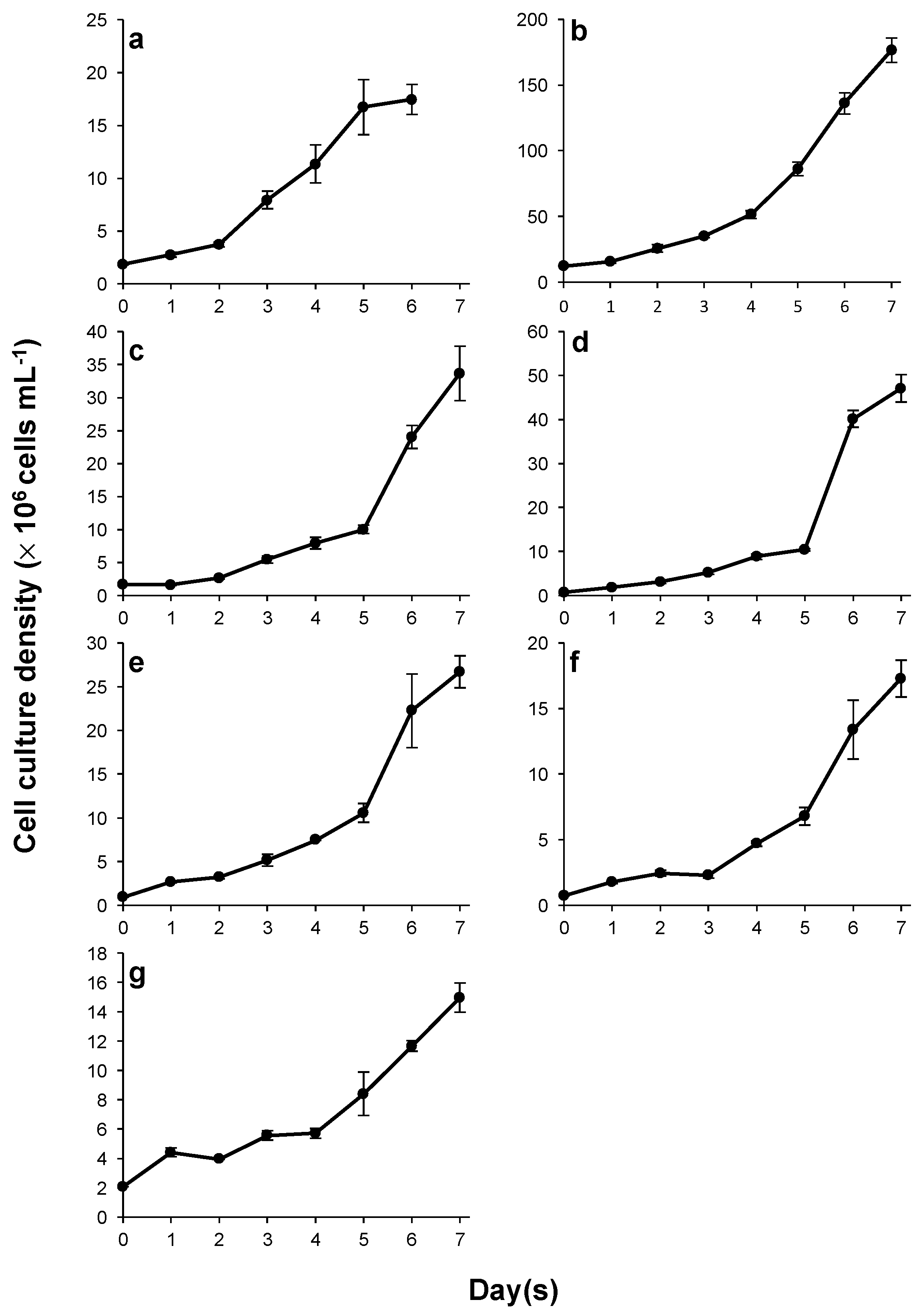
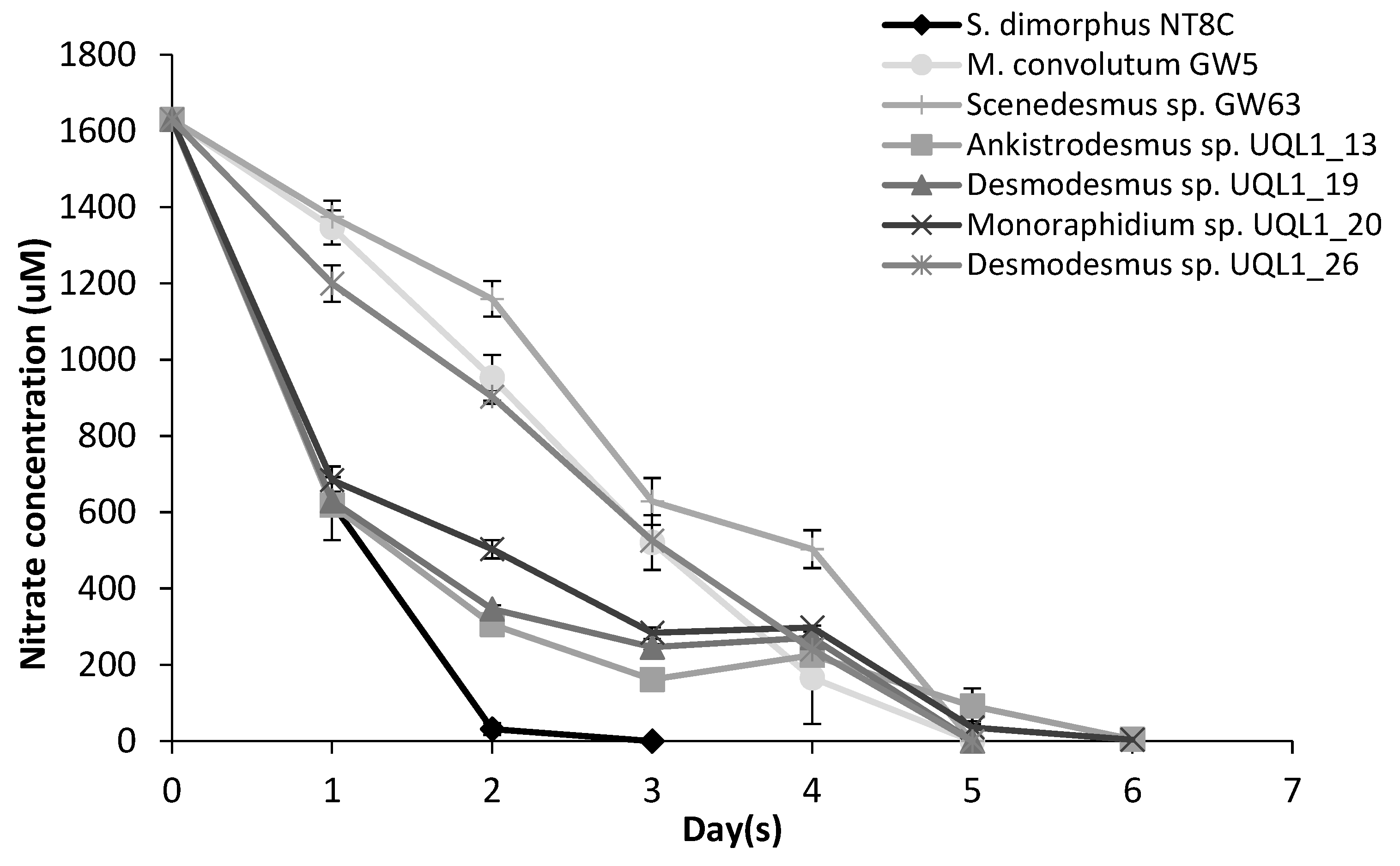
3.2. Cultivation and Growth
3.3. Fatty Acid Content and Composition
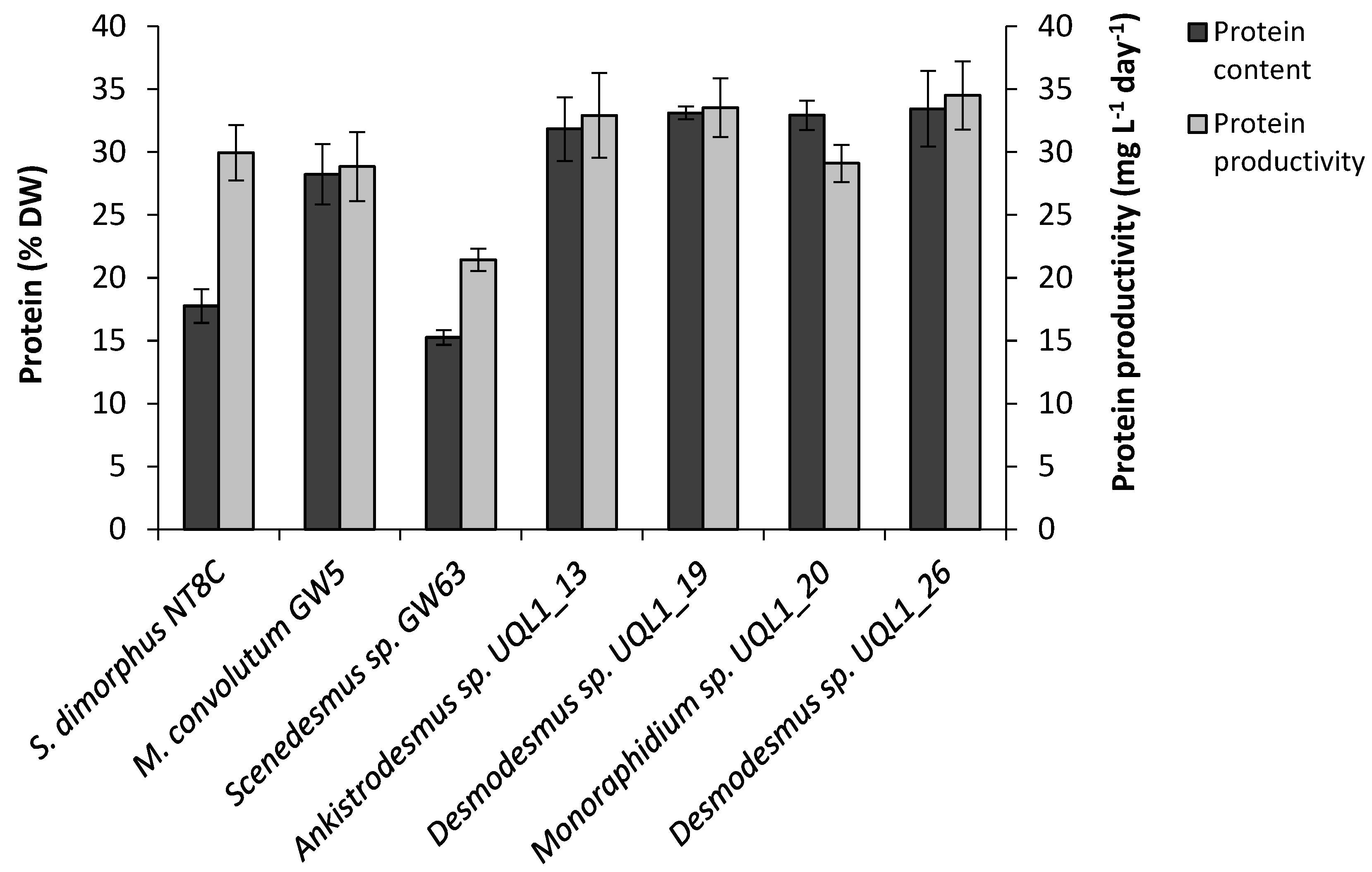
3.4. Total Protein Content
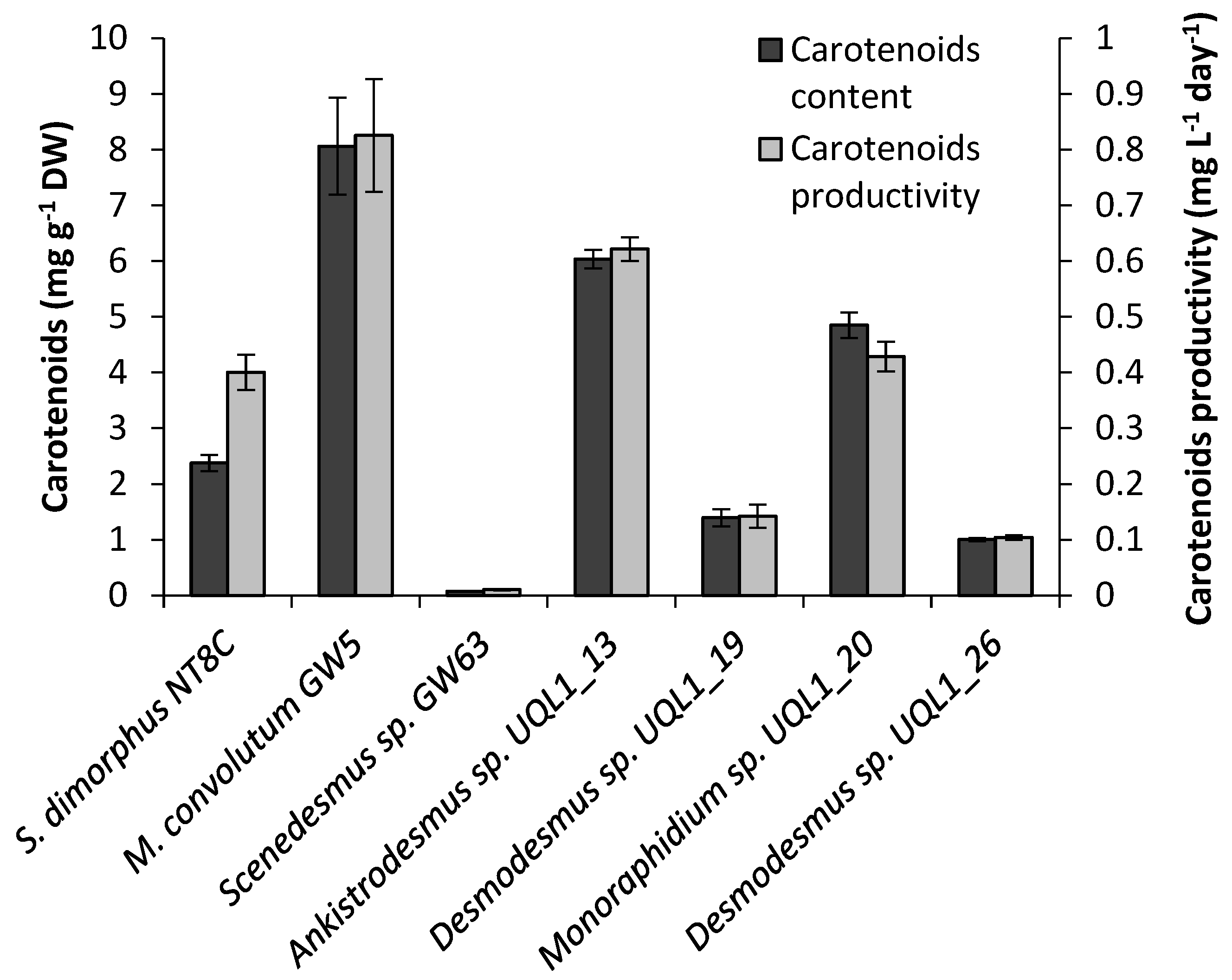
3.5. Total Carotenoid Content
4. Discussion
4.1. Sample Collection, Isolation, Identification and Selection of Microalgal Strains
4.2. Cultivation and Growth
4.3. Fatty Acid Content and Composition
4.4. Total Protein Content
4.5. Total Carotenoid Content
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.K.; Garg, S.; Timmins, M.; Zhang, E.S.; Thomas-Hall, S.R.; Schuhmann, H.; Li, Y.; Schenk, P.M. Isolation and evaluation of oil-producing microalgae from subtropical coastal and brackish waters. PLoS ONE 2012, 7, e40751. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.; Moheimani, N. Sustainable biofuels from algae. Mitig. Adapt. Strateg. Glob. Change 2013, 18, 13–25. [Google Scholar] [CrossRef]
- Schenk, P.M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second generation biofuels: High-efficiency microalgae for biodiesel production. BioEnergy Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Khan, S.A.; Rashmi Hussain, M.Z.; Prasad, S.; Banerjee, U.C. Prospects of biodiesel production from microalgae in India. Renew. Sustain. Energy Rev. 2009, 13, 2361–2372. [Google Scholar] [CrossRef]
- Wang, B.; Li, Y.; Wu, N.; Lan, C.Q. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Vardon, D.R.; Sharma, B.K.; Scott, J.; Yu, G.; Wang, Z.; Schideman, L.; Zhang, Y.; Strathmann, T.J. Chemical properties of biocrude oil from the hydrothermal liquefaction of Spirulina algae, swine manure, and digested anaerobic sludge. Bioresour. Technol. 2011, 102, 8295–8303. [Google Scholar] [CrossRef] [PubMed]
- Mallick, N. Biotechnological potential of immobilized algae for wastewater N, P and metal removal: A review. BioMetals 2002, 15, 377–390. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Commercial production of microalgae: Ponds, tanks, tubes and fermenters. J. Biotechnol. 1999, 70, 313–321. [Google Scholar] [CrossRef]
- Stolz, P.; Obermayer, B. Manufacturing microalgae for skincare. Cosmet. Toilet. 2005, 120, 99–106. [Google Scholar]
- Borowitzka, M. High-value products from microalgae—their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Ren. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Milledge, J. Commercial application of microalgae other than as biofuels: A brief review. Rev. Environ. Sci. Bio/Technol. 2011, 10, 31–41. [Google Scholar] [CrossRef]
- Hu, Q. Environmental effects on cell composition. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 114–122. [Google Scholar]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef]
- Adarme-Vega, T.; Lim, D.K.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Factories 2012, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, E.W. Microalgae for human and animal nutrition. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 461–503. [Google Scholar]
- Lopez, C.V.; Garcia Mdel, C.; Fernandez, F.G.; Bustos, C.S.; Chisti, Y.; Sevilla, J.M. Protein measurements of microalgal and cyanobacterial biomass. Bioresour. Technol. 2010, 101, 7587–7591. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Jin, E.; Polle, J.E.; Hong, K.L.; Sang, M.H.; Chang, M. Xanthophylls in microalgae: From biosynthesis to biotechnological mass production and application. J. Microbiol. Biotechnol. 2013, 13, 165–174. [Google Scholar]
- Lorenz, R.T.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.-V.; Parry, D.L. The gross chemical composition and fatty acid composition of 18 species of tropical Australian microalgae for possible use in mariculture. Aquaculture 1999, 170, 147–159. [Google Scholar] [CrossRef]
- Islam, M.; Magnusson, M.; Brown, R.; Ayoko, G.; Nabi, M.; Heimann, K. Microalgal species selection for biodiesel production based on fuel properties derived from fatty acid profiles. Energies 2013, 6, 5676–5702. [Google Scholar] [CrossRef] [Green Version]
- Lang, I.; Hodac, L.; Friedl, T.; Feussner, I. Fatty acid profiles and their distribution patterns in microalgae: A comprehensive analysis of more than 2000 strains from the SAG culture collection. BMC Plant Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, A.P.; Gouveia, L.; Bandarra, N.M.; Franco, J.M.; Raymundo, A. Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res. 2013, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Dectonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Duong, V.T.; Li, Y.; Nowak, E.; Schenk, P.M. Microalgae isolation and selection for prospective biodiesel production. Energies 2012, 5, 1835–1849. [Google Scholar] [CrossRef]
- Duong, V.T.; Bao, B.; Schenk, P.M. Oleaginous microalgae isolation and screening for lipid productivity using a standard protocol. In Hydrocarbon and Lipid Microbiology Protocols; McGenity, T.J., Timmis, K.N., Nogales Fernández, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Duong, V.T.; Ahmed, F.; Thomas-Hall, S.R.; Quigley, S.; Nowak, E.; Schenk, P.M. High protein- and high lipid-producing microalgae from northern australia as potential feedstock for animal feed and biodiesel. Front. Bioeng. Biotechnol. 2015, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, M.; Harrison, S.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. J. Appl. Phycol. 2009, 21, 93–507. [Google Scholar] [CrossRef]
- Wood, A.M.; Everroad, R.C.; Wingard, L.M. Measuring growth rates in microalgal cultures. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Academic Press: Burlington, MA, USA, 2005; pp. 269–285. [Google Scholar]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Huang, J.J.; Cheung, P.C. +UVA treatment increases the degree of unsaturation in microalgal fatty acids and total carotenoid content in Nitzschia closterium (Bacillariophyceae) and Isochrysis zhangjiangensis (Chrysophyceae). Food Chem. 2011, 129, 783–791. [Google Scholar] [CrossRef]
- Porra, R.; Thompson, W.; Kriedemann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- An, S.S.; Friedl, T.; Hegewald, E. Phylogenetic relationships of Scenedesmus and Scenedesmus-like coccoid green algae as Inferred from ITS-2 rDNA sequence comparisons. Plant Biol. 1999, 1, 418–428. [Google Scholar] [CrossRef]
- Borowitzka, M. Species and strain selection. In Developments in Applied Phycology; Borowitzka, M.A., Moheimani, N.R., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 5, pp. 77–89. [Google Scholar]
- Barclay, W.; Apt, K. Strategies for bioprospecting microalgae for potential commercial applications. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 69–79. [Google Scholar]
- Friedl, T.; O’Kelly, C.J. Phylogenetic relationships of green algae assigned to the genus Planophila (Chlorophyta): Evidence from 18S rDNA sequence data and ultrastructure. Eur. J. Phycol. 2002, 37, 373–384. [Google Scholar] [CrossRef]
- Krienitz, L.; Bock, C.; Nozaki, H.; Wolf, M. SSU rRNA gene phylogeny of morphospecies affiliated to the bioassay alga “Selenastrum capricornutum” recovered the polyphyletic origin of crescent-shaped Chlorophyta. J. Phycol. 2011, 47, 880–893. [Google Scholar] [CrossRef]
- Leliaert, F.; Verbruggen, H.; Vanormelingen, P.; Steen, F.; López-Bautista, J.M.; Zuccarello, G.C.; De Clerck, O. DNA-based species delimitation in algae. Eur. J. Phycol. 2014, 49, 179–196. [Google Scholar] [CrossRef] [Green Version]
- Moon-van der Staay, S.Y.; De Wachter, R.; Vaulot, D. Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 2001, 409, 607–610. [Google Scholar] [CrossRef]
- Piganeau, G.; Eyre-Walker, A.; Grimsley, N.; Moreau, H. How and why DNA barcodes underestimate the diversity of microbial eukaryotes. PLoS ONE 2011, 6, e16342. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.A. The microalgal cell. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 1–20. [Google Scholar]
- Lürling, M. Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction of defensive morphology. Ann. Limnol. Int. J. Limnol. 2009, 39, 85–101. [Google Scholar] [CrossRef] [Green Version]
- Alpermann, T.J.; Tillmann, U.; Beszteri, B.; Cembella, A.D.; John, U. Phenotypic variation and genotypic diversity in a planktonic population of the toxigenic marine dinoflagellate Alexandrium tamarense (Dinophyceae). J. Phycol. 2010, 46, 18–32. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Pflugmacher, S.; Proschold, T.; Walz, N.; Krienitz, L. Genotype versus phenotype variability in Chlorella and Micractinium (Chlorophyta, Trebouxiophyceae). Protist 2006, 157, 315–333. [Google Scholar] [CrossRef]
- Fawley, M.W.; Dean, M.L.; Dimmer, S.K.; Fawley, K.P. Evaluating the Morphospecies Concept in the Selenastraceae (Chlorophyceae, Chlorophyta)1. J. Phycol. 2005, 42, 142–154. [Google Scholar] [CrossRef]
- Krienitz, L.; Ustinova, I.; Friedl, T.; Huss, V.A.R. Traditional generic concepts versus 18S rRNA gene phylogeny in the green algal family selenastraceae (chlorophyceae, chlorophyta). J. Phycol. 2001, 37, 852–865. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Nakajima, H.; Anno, Y. Potential of Monoraphidium sp. GK12 for energy-saving astaxanthin production. J. Chem. Technol. Biotechnol. 2008, 83, 1578–1584. [Google Scholar] [CrossRef]
- Ruangsomboon, S.; Ganmanee, M.; Choochote, S. Effects of different nitrogen, phosphorus, and iron concentrations and salinity on lipid production in newly isolated strain of the tropical green microalga, Scenedesmus dimorphus KMITL. J. Appl. Phycol. 2012, 25, 867–874. [Google Scholar] [CrossRef]
- Vidyashankar, S.; Deviprasad, K.; Chauhan, V.S.; Ravishankar, G.A.; Sarada, R. Selection and evaluation of CO2 tolerant indigenous microalga Scenedesmus dimorphus for unsaturated fatty acid rich lipid production under different culture conditions. Bioresour. Technol. 2013, 144, 28–37. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, P.; He, C.; Li, J.; Tang, X.; Zhou, J.; Huang, Z. Isolation of a novel strain of Monoraphidium sp. and characterization of its potential application as biodiesel feedstock. Bioresour. Technol. 2012, 121, 256–262. [Google Scholar] [CrossRef]
- Radzun, K.A.; Wolf, J.; Jakob, G.; Zhang, E.; Stephens, E.; Ross, I.; Hankamer, B. Automated nutrient screening system enables high-throughput optimisation of microalgae production conditions. Biotechnol. Biofuels 2015, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerlimann, R.; de Nys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 107, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Lawton, R.J.; de Nys, R.; Paul, N.A. Selecting reliable and robust freshwater macroalgae for biomass applications. PLoS ONE 2013, 8, e64168. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, I.A.; Marques, S.S.I.; Cabanelas, I.T.D.; Pereira, S.A.; Druzian, J.I.; de Souza, C.O.; Vich, D.V.; de Carvalho, G.C.; Nascimento, M.A. Screening microalgae strains for biodiesel production: Lipid productivity and estimation of fuel quality aased on fatty acids profiles as selective criteria. BioEnergy Res. 2012, 6, 1–13. [Google Scholar] [CrossRef]
- Voltolina, D.; Cordero, B.; Nieves, M.; Soto, L.P. Growth of Scenedesmus sp. in artificial wastewater. Bioresour. Technol. 1999, 68, 265–268. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef]
- Via, S.; Lande, R. Genotype-environment interaction and the evolution of phenotypic plasticity. Evolution 1985, 39, 505–522. [Google Scholar] [CrossRef]
- Chen, C.Y.; Yeh, K.L.; Aisyah, R.; Lee, D.J.; Chang, J.S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Bischoff, H.W.; Bold, H.C. Some Soil Algae from Enchanted Rock and Related Algal Species; University of Texas: Austin, TX, USA, 1963; Volume 4. [Google Scholar]
- Allen, M.M.; Stanier, R.Y. Growth and division of some unicellular blue-green algae. J. Gen. Microbiol. 1968, 51, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Tang, H.; Ma, H.; Holland, T.C.; Ng, K.Y.; Salley, S.O. Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresour. Technol. 2011, 102, 1649–1655. [Google Scholar] [CrossRef]
- Samori, G.; Samori, C.; Guerrini, F.; Pistocchi, R. Growth and nitrogen removal capacity of Desmodesmus communis and of a natural microalgae consortium in a batch culture system in view of urban wastewater treatment: Part I. Water Res. 2013, 47, 791–801. [Google Scholar] [CrossRef]
- Richmond, A. Biological Principles of Mass Cultivation of Photoautotrophic Microalgae. In Handbook of Microalgal Culture; Richmond, A., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 169–204. [Google Scholar]
- Gosch, B.J.; Magnusson, M.; Paul, N.A.; de Nys, R. Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012, 4, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Kent, M.; Welladsen, H.M.; Mangott, A.; Li, Y. Nutritional evaluation of Australian microalgae as potential human health supplements. PLoS ONE 2015, 10, e0118985. [Google Scholar] [CrossRef]
- Martínez-Fernández, E.; Acosta-Salmón, H.; Southgate, P.C. The nutritional value of seven species of tropical microalgae for black-lip pearl oyster (Pinctada margaritifera, L.) larvae. Aquaculture 2006, 257, 491–503. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquacult. Int. 2006, 15, 1–9. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Waanders, J.; Ward, L.; Brown, L. Lipid redistribution by alpha-linolenic acid-rich chia seed inhibits stearoyl-CoA desaturase-1 and induces cardiac and hepatic protection in diet-induced obese rats. J. Nutr. Biochem. 2012, 23, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Dubois, V.; Breton, S.; Linder, M.; Fanni, J.; Parmentier, M. Fatty acid profiles of 80 vegetable oils with regard to their nutritional potential. Eur. J. Lipid Sci. Technol. 2007, 109, 710–732. [Google Scholar] [CrossRef]
- Martins, D.A.; Custodio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Bell, M.; Tocher, D. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new direction. In Lipids in Aquatic Ecosystems; Kainz, M., Brett, M.T., Arts, M.T., Eds.; Springer: New York, NY, USA, 2009; pp. 211–236. [Google Scholar]
- Hempel, N.; Petrick, I.; Behrendt, F. Biomass productivity and productivity of fatty acids and amino acids of microalgae strains as key characteristics of suitability for biodiesel production. J. Appl. Phycol. 2012, 24, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High lipid induction in microalgae for biodiesel production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Mendiola, J.; Plaza, M.; Ibañez, E. Screening for bioactive compounds from algae. In Advanced Biofuels and Bioproducts; Lee, J.W., Ed.; Springer: New York, NY, USA, 2013; pp. 833–872. [Google Scholar]
- Wright, T.; Lackey, R. Definitions of Feed Manufacturing and Livestock Nutrition Terms, Ontario Ministry of Agriculture, Food, and Rural Affairs. 2008. Available online: http://www.omafra.gov.on.ca/english/livestock/dairy/facts/08-039.pdf (accessed on 18 October 2015).
- Fujii, K.; Nakashima, H.; Hashidzume, Y.; Uchiyama, T.; Mishiro, K.; Kadota, Y. Potential use of the astaxanthin-producing microalga, Monoraphidium sp. GK12, as a functional aquafeed for prawns. J. Appl. Phycol. 2009, 22, 363–369. [Google Scholar] [CrossRef]
- Seyfabadi, J.; Ramezanpour, Z.; Amini Khoeyi, Z. Protein, fatty acid, and pigment content of Chlorella vulgaris under different light regimes. J. Appl. Phycol. 2011, 23, 721–726. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Chen, W.; Shen, H.; Han, D.; Li, Y.; Jones, H.D.T.; Timlin, J.A.; Hu, Q. Basic culturing and analytical measurement techniques. In Handbook of Microalgal Culture; Richmond, A., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 37–68. [Google Scholar]
- Pignolet, O.; Jubeau, S.; Vaca-Garcia, C.; Michaud, P. Highly valuable microalgae: Biochemical and topological aspects. J. Ind. Microbiol. Biotechnol. 2013, 40, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.-C.; Ho, S.-H.; Lee, D.-J.; Chen, C.-Y.; Huang, C.-C.; Chang, J.-S. Characterization, extraction and purification of lutein produced by an indigenous microalga Scenedesmus obliquus CNW-N. Biochem. Eng. J. 2013, 78, 24–31. [Google Scholar] [CrossRef]
- Schüler, L.; Greque de Morais, E.; Trovão, M.; Machado, A.; Carvalho, B.; Carneiro, M.; Maia, I.; Soares, M.; Duarte, P.; Barros, A.; et al. Isolation and characterization of novel Chlorella vulgaris mutants with low chlorophyll and improved protein contents for food applications. Front. Bioeng. Biotechnol. 2020, 8, 469. [Google Scholar] [CrossRef]
- Mendes-Pinto, M.M.; Raposo, M.F.J.; Bowen, J.; Young, A.J.; Morais, R. Evaluation of different cell disruption processes on encysted cells of Haematococcus pluvialis: Effects on astaxanthin recovery and implications for bio-availability. J. Appl. Phycol. 2001, 13, 19–24. [Google Scholar] [CrossRef]
- Boussiba, S. Carotenogenesis in the green alga Haematococcus pluvialis: Cellular physiology and stress response. Physiol. Plant. 2000, 108, 111–117. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Dunaliella: Biology, production, and markets. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 359–368. [Google Scholar]
- Han, D.; Li, Y.; Hu, Q. Biology and commercial aspects of Haematococcus pluvialis. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2013; pp. 388–405. [Google Scholar]
- Latasa, M.; Van Lenning, K.; Garrido, J.; Scharek, R.; Estrada, M.; Rodríguez, F.; Zapata, M. Losses of chlorophylls and carotenoids in aqueous acetone and methanol extracts prepared for RPHPLC analysis of pigments. Chromatographia 2001, 53, 385–391. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.; Lee, S.; Jin, E. Development of a Chlorella vulgaris mutant by chemical mutagenesis as a producer for natural violaxanthin. Algal Res. 2020, 46, 101790. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; He, M.; Gong, Y.; Huang, J. Induced high-yield production of zeaxanthin, lutein, and β-carotene by a mutant of Chlorella zofingiensis. J. Agricult. Food Chem. 2018, 66, 891–897. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Producer | Amount Added (mg L−1) | Final Concentration in the Media (µM) |
|---|---|---|---|
| NH4H2PO4 | Yates, Orica Australia Pty Ltd. | 17.25 | 150 |
| KNO3 | Biolab (Aust) Ltd. | 164 | 1624 |
| MgSO4.7H2O | Chem Supply | 82 | 333 |
| Sea salt | Aquasonic | 25 | 431 |
| RapiSol Mi6 EDTA Chelated Micro Nutrient | Agspec Australia Pty Ltd. | 4 | N/A |
| RapiSol Iron EDTA Chelated Micro Nutrient | Agspec Australia Pty Ltd. | 4 | N/A |
| Strain | Location | Fluctuating Environment | Genbank Accession |
|---|---|---|---|
| Monoraphidium convolutum GW5 | Woods Grains Pond, Goondiwindi 28°32′03″ S 150°19′39″ E | Eutrophic freshwater | KT893828 |
| Scenedesmus sp. GW63 | Woods Grains Pond, Goondiwindi 28°31′37″ S 150°17′29″ E | Eutrophic freshwater | KT893831 |
| Ankistrodesmus sp. UQL1_13 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893832 |
| Desmodesmus sp. UQL1_19 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893836 |
| Monoraphidium sp. UQL1_20 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893837 |
| Desmodesmus sp. UQL1_26 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic Ffeshwater | KT893841 |
| Scenedesmus dimorphus NT8C | Douglas Daly Research Farm, Winellie 13°49′59″ S 131°11′12″ E | High temperature freshwater | KF286273 |
| Ochromonas sp. PH1_1 | Algae Energy Farm open pond, Pinjarra Hills 27°32′21″ S 152°55′24″ E | Eutrophic freshwater | KT804909 |
| Spumella sp. GW31 | Golf Course Pond, Goondiwindi 28°31′45″ S 150°18′27″ E | Eutrophic freshwater | KT893827 |
| Pseudodidymocystis planctonica GW61 | Woods Grains Pond, Goondiwindi 28°31′37″ S 150°17′29″ E | Eutrophic freshwater | KT893829 |
| Scenedesmus sp. GW62 | Woods Grains Pond, Goondiwindi 28°31′37″ S 150°17′29″ E | Eutrophic freshwater | KT893830 |
| Pseudomuriella sp. UQL1_15 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893833 |
| Kirchneriella sp. UQL1_16 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893834 |
| Kirchneriella sp. UQL1_18 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893835 |
| Desmodesmus sp. UQL1_22 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893838 |
| Protodesmus sp. UQL1_23 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893839 |
| Desmodesmus sp. UQL1_24 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893840 |
| Nitzschia palea UQL1_31 | UQ Lake, St Lucia 27°29′55″ S 153°0′58″ E | Eutrophic freshwater | KT893842 |
| Chaetoceros calcitrans BR1_3 | Brisbane River, Brisbane 27°29′30″ S 153°0′47″ E | Eutrophic tidal brackish | KT893843 |
| Achnanthes sp. BR1_5 | Brisbane River, Brisbane 27°29′30″ S 153°0′47″ E | Eutrophic tidal brackish | KT893844 |
| Thalassiosira sp. NB1_1 | Nudgee Beach, Brisbane 27°21′3″ S 153°6′23″ E | Tidal marine | KT893845 |
| Navicula sp. NB1_2 | Nudgee Beach, Brisbane 27°21′3″ S 153°6′23″ E | Tidal marine | KT893846 |
| Chaetoceros sp. NB1_9 | Nudgee Beach, Brisbane 27°21′3″ S 153°6′23″ E | Tidal marine | KT893847 |
| Navicula sp. NB1_13 | Nudgee Beach, Brisbane 27°21′3″ S 153°6′23″ E | Tidal marine | KT893848 |
| Amphora sp. NC1_1 | Nudgee Creek, Nudgee Beach 27°21′1″ S 153°6′17″ E | Tidal eutrophic marine | KT893849 |
| Cylindrotheca sp. NC1_4 | Nudgee Creek, Nudgee Beach 27°21′1″ S 153°6′17″ E | Tidal eutrophic marine | KT893850 |
| Strain | Average Growth Rate | Cell Densitymax (×106 cells mL−1) | Doubling Time (days) | Biomass Productivity (g L−1 day−1) | ||
|---|---|---|---|---|---|---|
| Biomass Productivity LL | Biomass Productivity AVE | Biomass Productivity ND | ||||
| S. dimorphus NT8C | 0.44 a | 22.54 | 1.59 a | 0.17 a | 0.24 a | 0.37 a |
| M. convolutum GW5 | 0.38 a | 176.50 | 1.81 a | 0.10 b | 0.21 a,b | 0.35 a,b,* |
| Scenedesmus sp. GW63 | 0.42 a | 33.67 | 1.64 a | 0.14 a | 0.27 a | 0.44 a |
| Ankistrodesmus sp. UQL1_13 | 0.60 b | 47.08 | 1.15 b | 0.09 b | 0.22 a | 0.37 a |
| Desmodesmus sp. UQL1_19 | 0.48 a,b | 26.71 | 1.44 a,b | 0.10 b | 0.19 b | 0.30 b |
| Monoraphidium sp. UQL1_20 | 0.45 a | 17.29 | 1.54 a | 0.09 b | 0.16 b | 0.25 b |
| Desmodesmus sp. UQL1_26 | 0.28 c | 14.96 | 2.45 c | 0.10 b | 0.19 b | 0.31 b |
| Fatty Acid | S. dimorphus NT8C | M. convolutum GW5 | Scenedesmus sp. GW63 | Ankistrodesmus sp. UQL1_13 | Desmodesmus sp. UQL1_19 | Monoraphidium sp. UQL1_20 | Desmodesmus sp. UQL1_26 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C14:0 | 0.3 | ± | 0.0 | 0.4 | ± | 0.0 | 0.2 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 |
| C14:1 | 0.3 | ± | 0.0 | 0.4 | ± | 0.0 | 0.2 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 |
| C15:0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 |
| C16:0 | 20.4 | ± | 0.3 | 21.9 | ± | 0.4 | 17.9 | ± | 0.4 | 25.7 | ± | 0.3 | 25.7 | ± | 0.1 | 18.8 | ± | 0.4 | 21.4 | ± | 0.5 |
| C16:1(n-9) | 1.1 | ± | 0.0 | 0.5 | ± | 0.0 | 1.2 | ± | 0.0 | 0.7 | ± | 0.0 | 0.6 | ± | 0.0 | 0.7 | ± | 0.1 | 1.4 | ± | 0.1 |
| C16:1(n-7) | 0.2 | ± | 0.0 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.2 | ± | 0.0 | 0.4 | ± | 0.1 | 0.5 | ± | 0.1 | 0.3 | ± | 0.0 |
| C16:2(n-6) | 0.8 | ± | 0.0 | 0.6 | ± | 0.0 | 0.7 | ± | 0.0 | 1.0 | ± | 0.0 | 0.6 | ± | 0.0 | 0.7 | ± | 0.0 | 1.1 | ± | 0.0 |
| C16:3(n-3) | 4.2 | ± | 0.1 | 1.3 | ± | 0.0 | 3.2 | ± | 0.2 | 2.3 | ± | 0.1 | 3.6 | ± | 0.1 | 3.1 | ± | 0.0 | 5.2 | ± | 0.2 |
| C16:4(n-3) | 5.6 | ± | 0.3 | 6.4 | ± | 0.3 | 2.7 | ± | 0.2 | 4.1 | ± | 0.2 | 3.7 | ± | 0.1 | 5.6 | ± | 0.5 | 3.2 | ± | 0.1 |
| C17:0 | 0.2 | ± | 0.0 | 1.3 | ± | 0.1 | 0.1 | ± | 0.0 | 1.8 | ± | 0.0 | 0.4 | ± | 0.0 | 0.4 | ± | 0.0 | 0.3 | ± | 0.0 |
| C17:1(n-7) | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.2 | ± | 0.0 | 0.1 | ± | 0.0 | 0.3 | ± | 0.0 | 0.1 | ± | 0.0 | 0.2 | ± | 0.0 |
| C18:0 | 2.5 | ± | 0.2 | 1.6 | ± | 0.1 | 2.2 | ± | 0.2 | 2.7 | ± | 0.0 | 3.5 | ± | 0.3 | 2.2 | ± | 0.2 | 2.6 | ± | 0.2 |
| C18:1(n-9) | 27.9 | ± | 1.3 | 28.5 | ± | 1.1 | 30.2 | ± | 1.0 | 26.4 | ± | 0.2 | 28.4 | ± | 0.3 | 32.7 | ± | 2.2 | 30.6 | ± | 0.4 |
| C18:2(n-6) | 9.0 | ± | 0.4 | 7.0 | ± | 0.3 | 10.1 | ± | 0.3 | 8.9 | ± | 0.2 | 6.8 | ± | 0.1 | 7.3 | ± | 0.2 | 10.2 | ± | 0.1 |
| C18:3(n-6) | 0.6 | ± | 0.0 | 0.2 | ± | 0.0 | 0.1 | ± | 0.0 | 0.2 | ± | 0.0 | 1.1 | ± | 0.0 | 0.5 | ± | 0.1 | 1.4 | ± | 0.1 |
| C18:3(n-3) | 19.3 | ± | 0.4 | 16.5 | ± | 0.6 | 20.0 | ± | 0.8 | 14.6 | ± | 0.3 | 14.9 | ± | 0.2 | 15.1 | ± | 0.5 | 11.4 | ± | 0.0 |
| C18:4(n-3) | 2.2 | ± | 0.1 | 3.4 | ± | 0.2 | 2.5 | ± | 0.2 | 3.4 | ± | 0.1 | 2.2 | ± | 0.0 | 3.6 | ± | 0.3 | 2.2 | ± | 0.1 |
| C20:0 | 0.1 | ± | 0.0 | 0.2 | ± | 0.0 | 0.2 | ± | 0.0 | 0.2 | ± | 0.0 | 0.2 | ± | 0.0 | 0.1 | ± | 0.0 | 0.2 | ± | 0.0 |
| C20:1(n-9) | 0.3 | ± | 0.0 | 0.8 | ± | 0.0 | 2.1 | ± | 0.2 | 0.7 | ± | 0.0 | 0.4 | ± | 0.0 | 1.0 | ± | 0.1 | 0.3 | ± | 0.0 |
| C20:2(n-6) | – | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | – | – | – | 0.1 | ± | 0.0 | ||||||||
| C20:3(n-3) | – | – | 0.1 | ± | 0.0 | – | – | – | – | ||||||||||||
| C20:5(n-3) | – | – | – | – | – | – | – | ||||||||||||||
| C22:0 | 0.2 | ± | 0.0 | 0.9 | ± | 0.0 | 0.3 | ± | 0.0 | 0.2 | ± | 0.0 | 0.2 | ± | 0.0 | 0.3 | ± | 0.1 | 0.3 | ± | 0.0 |
| C22:1(n-9) | – | 0.2 | ± | 0.0 | 0.2 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | 0.1 | ± | 0.0 | ||
| C22:6(n-3) | – | – | – | – | – | – | – | ||||||||||||||
| C24:0 | 0.2 | ± | 0.0 | 1.0 | ± | 0.1 | – | 0.8 | ± | 0.1 | 0.1 | ± | 0.0 | 0.7 | ± | 0.0 | 0.1 | ± | 0.0 | ||
| Other fatty acids | 4.7 | ± | 0.5 | 6.2 | ± | 0.3 | 5.0 | ± | 0.6 | 5.2 | ± | 0.6 | 6.1 | ± | 0.4 | 5.8 | ± | 0.9 | 6.8 | ± | 0.7 |
| Total SFA | 23.8 | ± | 0.4 | 27.4 | ± | 0.5 | 21.0 | ± | 0.6 | 31.8 | ± | 0.2 | 30.5 | ± | 0.3 | 22.8 | ± | 0.4 | 25.2 | ± | 0.7 |
| Total MUFA | 30.0 | ± | 1.2 | 30.9 | ± | 1.1 | 34.5 | ± | 0.9 | 28.5 | ± | 0.3 | 30.3 | ± | 0.3 | 35.4 | ± | 2.1 | 33.3 | ± | 0.3 |
| Total PUFA | 41.6 | ± | 1.1 | 35.4 | ± | 1.4 | 39.5 | ± | 1.4 | 34.5 | ± | 0.8 | 33.0 | ± | 0.2 | 36.0 | ± | 1.6 | 34.8 | ± | 0.3 |
| ω-3 PUFA | 31.2 | ± | 0.8 | 27.5 | ± | 1.1 | 28.5 | ± | 1.2 | 24.4 | ± | 0.6 | 24.4 | ± | 0.1 | 27.4 | ± | 1.3 | 22.0 | ± | 0.2 |
| ω-6 PUFA | 10.3 | ± | 0.4 | 7.9 | ± | 0.3 | 11.0 | ± | 0.3 | 10.1 | ± | 0.3 | 8.6 | ± | 0.1 | 8.6 | ± | 0.3 | 12.8 | ± | 0.2 |
| ω-6/ω-3 | 0.3 | ± | 0.0 | 0.3 | ± | 0.0 | 0.4 | ± | 0.0 | 0.4 | ± | 0.0 | 0.4 | ± | 0.0 | 0.3 | ± | 0.0 | 0.6 | ± | 0.0 |
| TFA (% DW) | 11.8 | ± | 0.3 | 13.4 | ± | 0.6 | 12.1 | ± | 1.0 | 14.9 | ± | 1.4 | 12.8 | ± | 0.9 | 15.9 | ± | 0.6 | 11.7 | ± | 0.5 |
| TFA productivity (mg L-1 day-1) | 30.8 | ± | 2.6 | 28.0 | ± | 1.1 | 32.6 | ± | 4.2 | 32.2 | ± | 3.1 | 23.6 | ± | 1.4 | 25.2 | ± | 3.4 | 22.3 | ± | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, B.; Thomas-Hall, S.R.; Schenk, P.M. Fast-Tracking Isolation, Identification and Characterization of New Microalgae for Nutraceutical and Feed Applications. Phycology 2022, 2, 86-107. https://doi.org/10.3390/phycology2010006
Bao B, Thomas-Hall SR, Schenk PM. Fast-Tracking Isolation, Identification and Characterization of New Microalgae for Nutraceutical and Feed Applications. Phycology. 2022; 2(1):86-107. https://doi.org/10.3390/phycology2010006
Chicago/Turabian StyleBao, Boer, Skye R. Thomas-Hall, and Peer M. Schenk. 2022. "Fast-Tracking Isolation, Identification and Characterization of New Microalgae for Nutraceutical and Feed Applications" Phycology 2, no. 1: 86-107. https://doi.org/10.3390/phycology2010006
APA StyleBao, B., Thomas-Hall, S. R., & Schenk, P. M. (2022). Fast-Tracking Isolation, Identification and Characterization of New Microalgae for Nutraceutical and Feed Applications. Phycology, 2(1), 86-107. https://doi.org/10.3390/phycology2010006