
Effects of Short- and Long-Distance Road Transport and Temperament on the Magnitude of β-Endorphin Response in Limousine Bulls


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Temperament Classification
2.3. Sampling
- (1)
- At the growing–finishing farm and immediately before loading, thus representing baseline conditions (10.00–11.00).
- (2)
- At the Soual collection center, after a short-distance road transport of 55 km lasting approximately 1 h, within a restore time of 12 h (16.00–17.00).
- (3)
- Upon arrival in Sicily, after prolonged transport covering 2.200 km and 1 h after unloading (15.00–16.00).
- (4)
- At the finishing farm, 15 days after transport (15.00–16.00).
- (5)
- All bulls were individually weighed after the first and last blood sampling using a mechanical cattle scale (0.5–1000 kg) in base conditions and 15 days later, respectively.
2.4. Statistical Analysis
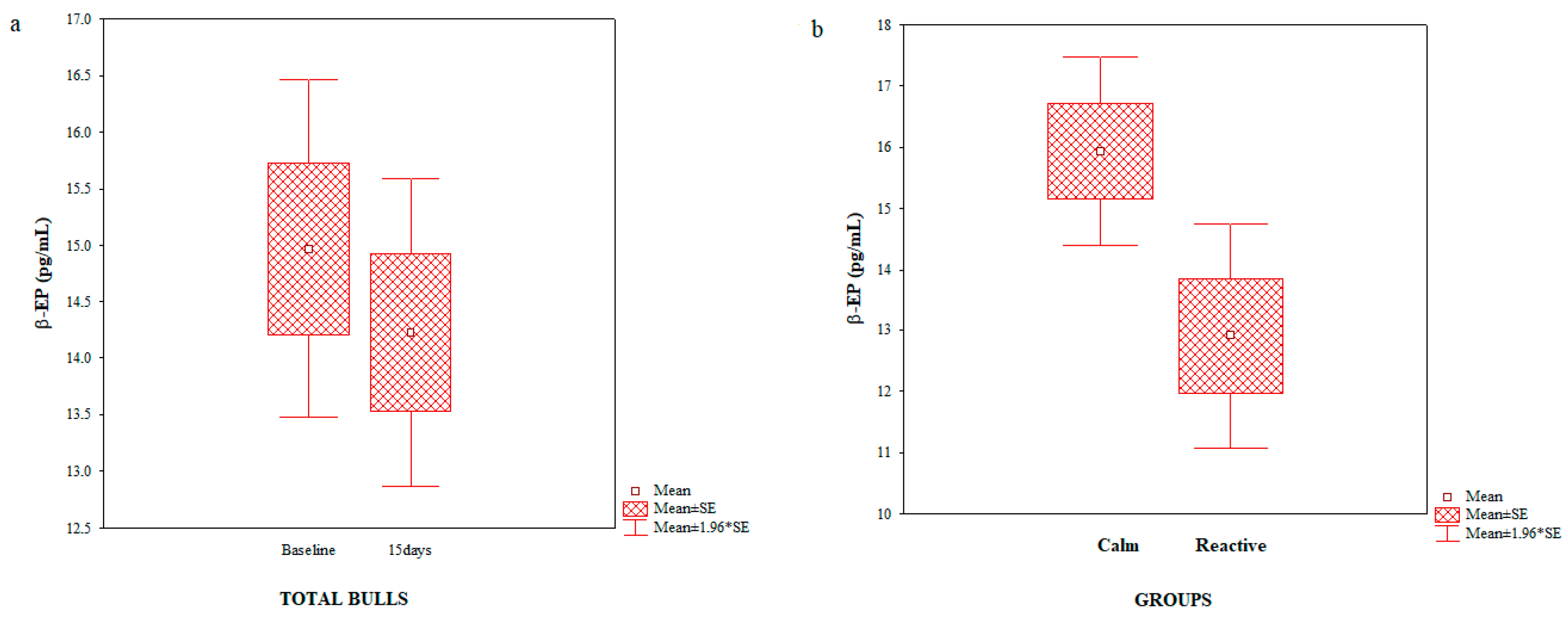
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bravo, V.; Gallo, C.; Acosta-Jamett, G. Effects of short transport and prolonged fasting in beef calves. Animals 2018, 8, 170. [Google Scholar] [CrossRef]
- Tarrant, V.; Grandin, T. Cattle transport. In Livestock Handling and Transport; Grandin, T., Ed.; CABI Publishing: Wallingford, UK; New York, NY, USA, 2000; pp. 109–126. [Google Scholar]
- Broom, D.M. Causes of poor welfare in large animal during transport. Vet. Res. Commun. 2003, 27 (Suppl. S1), 515–518. [Google Scholar]
- Hartung, J. Effects of transport on health of farm animals. Vet. Res. Commun. 2003, 27 (Suppl. S1), 525–527. [Google Scholar] [PubMed]
- Averós, X.; Martin, S.; Riu, M.; Serratosa, J.; Gosálvez, L.F. Stress response of extensively reared young bulls being transported to growing-finishing farms under Spanish summer commercial conditions. Livest. Sci. 2008, 119, 174–182. [Google Scholar]
- Ashenafi, D.; Yidersal, E.; Hussen, E.; Solomon, T.; Desiye, M. The Effect of long distance transportation stress on Cattle: A Review. Biomed. J. Sci. Tech. Res. 2018, 3, 3304–3308. [Google Scholar] [CrossRef]
- Earley, B.; Murray, M.; Prendiville, D.J. Effect of road transport for up to 24 hours followed by twenty-four hour recovery on live weight and physiological responses of bulls. BMC Vet. Res. 2010, 6, 8. [Google Scholar]
- Fazio, E.; Medica, P.; Cravana, C.; Ferlazzo, A. Release of β-endorphin, adrenocorticotropic hormone and cortisol in response to machine milking of dairy cows. Vet. World 2015, 8, 284–289. [Google Scholar]
- Odore, R.; D’ Angelo, A.; Badino, P.; Bellino, C.; Pagliasso, S.; Re, G. Road transportation effects blood hormone levels and lymphocyte glucocorticoid and β-adrenergic receptor concentrations in calves. Vet. J. 2004, 168, 297–303. [Google Scholar] [CrossRef]
- Tadich, N.; Gallo, H.; Bustamante, H.; Schwerter, M.; Van Schaik, G. Effects of transport and lairage time on some blood constituents of Friesian-Cross steers in Chile. Livest. Prod. Sci. 2005, 93, 223–233. [Google Scholar] [CrossRef]
- Parker, A.J.; Hamlin, G.P.; Coleman, C.J.; Fitzpattrick, L.A. Quantitative analysis of acid-base balance in Bosindi cussteers subjected to transportation of long duration. J. Anim. Sci. 2003, 81, 1434–1439. [Google Scholar]
- Eniolorunda, O.O.; Fashina, O.E.; Aro, O.O. Adaptive physiological response to load time stress during transportation of cattle In Nigeria. Arch. Zootec. 2009, 58, 223–230. [Google Scholar]
- Burdick, N.C.; Randel, R.D.; Caroll, J.A.; Welsh, T.H., Jr. Interactions between temperament, stress and immune function in cattle. Int. J. Zool. 2011, 95, 197–201. [Google Scholar] [CrossRef]
- Cooper, C.; Evans, A.C.O.; Cook, S.; Rawlings, N.C. Cortisol, progesterone and B-endorphin response to stress in calves. Can. J. Anim. Sci. 1995, 75, 197–201. [Google Scholar] [CrossRef]
- Benamar, K.; Yondorf, M.; Barreto, V.T.; Geller, E.B.; Adler, M.W. Deletion of mu-opioid receptor in mice alters the development of acute neuroinflammation. J. Pharmacol. Exp. Ther. 2007, 323, 990–994. [Google Scholar] [PubMed]
- Osawa, T.; Nakao, T.; Moriyoshi, M.; Nakada, K. Plasma beta endorphin around parturition and its relationship to cortisol level and resumption of pituitary and ovarian functions in dairy cows. Anim. Reprod. Sci. 1998, 52, 27–38. [Google Scholar] [PubMed]
- Shaw, F.D.; Tume, R.K. Beta-endorphin and cortisol concentrations in plasma of cattle. Aust. Vet. J. 1990, 67, 423–424. [Google Scholar]
- Bodnar, R.J. Endogenous opioid modulation of food intake and body weight: Implications for opioid influences upon motivation and addiction. Peptides 2019, 116, 42–62. [Google Scholar]
- Souza, G.F.P.; Solon, C.; Nascimento, L.F.; De-Lima-Junior, J.C.; Nogueira, G.; Moura, R.; Rocha, G.Z.; Fioravante, M.; Bobbo, V.; Morari, J.; et al. Defective regulation of POMC precedes hypothalamic in diet-induced obesity. Sci. Rep. 2016, 6, 29290. [Google Scholar] [CrossRef]
- Aksoy, G.; Süzergöz, F.; Polat, P.F. Investigation of serum beta-endorphin levels in cows slaughtered with halal slaughter method. Firat Univ. Vet. J. Health Sci. 2018, 32, 165–167. [Google Scholar]
- Franceschini, R.; Venturini, P.L.; Cataldi, A.; Barreca, T.; Ragni, N.; Rolandi, E. Plasma beta-endorphin concentrations during suckling in lactating women. Br. J. Obstet. Gynaecol. 1989, 96, 711–713. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Schams, D.; Blum, J.W. Milk removal in familiar and unfamiliar surroundings: Concentrations of oxytocin, prolactin, cortisol and β-endorphin. J. Dairy Res. 1993, 60, 449–456. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Pfeilsticker, H.U.; Blum, J.W. Milk yield, oxytocin and beta-endorphin gradually normalize during repeated milking in unfamiliar surroundings. J. Dairy Res. 1996, 63, 191–200. [Google Scholar] [CrossRef]
- Bustamante, H.A.; Rodríguez, A.R.; Herzberg, D.E.; Werner, M.P. Stress and pain response after oligofructose induced-lameness in dairy heifers. J. Vet. Sci. 2015, 16, 405–411. [Google Scholar] [CrossRef] [PubMed]
- El-Malky, O.M.; El-Ghait, H.A.A. Peripartum beta endorphin levels in relation to labor disorders and post- calving reproductive performance of buffaloes. J. Anim. Poultry Prod. 2016, 7, 255–263. [Google Scholar] [CrossRef]
- Rodriguez, A.R.; Herzberg, D.E.; Werner, M.P.; Müller, H.Y.; Bustamante, H.A. Plasma concentration of norepinephrine, β-endorphin, and substance P in lame dairy cows. J. Vet. Res. 2018, 62, 193–197. [Google Scholar] [CrossRef]
- Scott, S.L.; Schaefer, A.L.; Jones, S.D.; Mears, G.J.; Stanley, R.W. Stress indicator and lean tissue yield in transported cattle treated with electrolyte. In Proceedings of the 39th International Congress of Meat Science and Technology, Calgary, AB, Canada, 1–6 August 1993; paper 52. p. 22. [Google Scholar]
- Fazio, E.; Medica, P.; Cravana, C.; Giacoppo, E.; Ferlazzo, A. Physiological variables of horses after road transport. Animal 2009, 3, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, S.; Brandt, H.R.; Konig, S.; Erhardt, G.; Gauly, M. Temperament traits of beef calves measured under field conditions and their relationships to performance. J. Anim. Sci. 2010, 88, 1982–1989. [Google Scholar] [CrossRef]
- Sant’Anna, A.C.; Baldi, F.; Valente, T.S.; Albuquerque, L.G.; Menezes, L.M.; Boligon, A.A.; Paranhos da Costa, M.J.R. Genetic associations between temperament and performance traits in Nellore beef cattle. J. Anim. Breed. Genet. 2014, 132, 42–50. [Google Scholar] [CrossRef]
- Marahrens, M.; von Richthofen, I.; Schmeiduch, S.; Hartung, J. Special problems of long distance road transports of cattle. Dtsch. Tierarztl. Wochenschr. 2003, 110, 120–125. [Google Scholar]
- European Commission. SCAHAW: The Welfare of Animals During Transport (Details for Horses, Pigs, Sheep and Cattle). Report of the Scientific Committee on Animal Health and Animal Welfare (SCAHAW); European Commission: Brussels, Belgium, 2002; p. 130. [Google Scholar]
- Warriss, P.D.; Brown, S.N.; Knowles, T.G.; Kestin, S.C.; Edwards, J.E.; Dolan, S.K.; Phillips, A.J. Effects on cattle of transport by road for up to 15 h. Vet. Rec. 1995, 136, 319–323. [Google Scholar] [CrossRef]
- Burrow, H.M.; Seifert, G.W.; Corbert, N.J. A new technique for measuring temperament in cattle. Proc. Australian Soc. Anim. Prod. 1988, 17, 154–157. Available online: https://api.semanticscholar.org/CorpusID:55498584 (accessed on 1 January 1998).
- Curley, K.O., Jr.; Paschal, J.C.; Welsh, T.H., Jr.; Randel, R.D. Exit velocity as a measurement of cattle temperament is repeatable and associated with serum concentration of cortisol in Brahman bulls. J. Anim. Sci. 2006, 84, 3100–3103. [Google Scholar] [CrossRef] [PubMed]
- Buckham Sporer, K.R.; Weber, P.S.D.; Burton, J.L.; Earley, B.; Crowe, M.A. Transportation of young beef bulls alters circulating physiological parameters that may be effective biomarkers of stress. J. Anim. Sci. 2008, 86, 1325–1334. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Willard, S.T.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Relationship between temperament and transportation with rectal temperature and serum concentrations of cortisol and epinephrine in bulls. Livest. Sci. 2010, 129, 166–172. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Randel, R.D.; Willard, S.T.; Vann, R.C.; Chase, C.C., Jr.; Lawhon, S.D.; Hulbert, L.E.; Welsh, T.H., Jr. Influence of temperament and transportation on physiological and endocrinological parameters in bulls. Livest. Sci. 2011, 139, 213–221. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Alberghina, D.; Cavaleri, S.; Ferlazzo, A. Effect of long-distance road transport on thyroid and adrenal function and haematocrit values in Limousin cattle: Influence of body weight decrease. Vet. Res. Commun. 2005, 29, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Fazio, E.; Medica, P.; Cravana, C.; Cavaleri, S.; Ferlazzo, A. Effect of temperament and prolonged transportation on endocrine and functional variables in young beef bulls. Vet. Rec. 2012, 171, 644. [Google Scholar] [CrossRef]
- Fazio, E.; Cravana, C.; Medica, P.; Ferlazzo, A. Cortisol response to road transport stress in calm and nervous stallions. J. Vet. Behav. 2013, 8, 231–237. [Google Scholar] [CrossRef]
- Hall, S.; Broom, D.; Goode, J.; Lloyd, D.; Parrott, R.; Rodway, R. Physiological responses of sheep during long road journeys involving ferry crossings. Anim. Sci. 1999, 69, 19–27. [Google Scholar] [CrossRef]
- Micera, E.; Dimatteo, S.; Grimaldi, M.; Marsico, G.; Zarilli, A. Stress indicators in steers at slaughtering. Ital. J. Anim. Sci. 2007, 6, 457–459. [Google Scholar] [CrossRef]
- Geburt, K.; Piechotta, M.; von Borstel, U.K.; Gauly, M. Influence of testosterone on the docility of German Simmental and Charolais heifers during behavior tests. Physiol. Behav. 2015, 141, 164–171. [Google Scholar] [PubMed]
- Curley, K.O., Jr.; Neuendorff, D.A.; Lewis, A.W.; Cleere, J.J.; Welsh, T.H., Jr.; Randel, R.D. Functional characteristics of the bovine hypothalamic-pituitary-adrenal axis vary with temperament. Horm. Behav. 2008, 53, 20–27. [Google Scholar] [PubMed]
- Francolino da Silva, M.K.; Landgraf, R.G.; Nichi, M.; Sant’Ana Borges, M.; Arantes Pereira, M.F.; Crespilho, M. Influence of animal temperament on body and reproductive development of beef bulls. Anim. Reprod. Sci. 2025, 274, 107788. [Google Scholar]
- Cafe, L.M.; Robinson, D.L.; Ferguson, D.M.; McIntyre, B.L.; Geesink, G.H.; Greenwood, P.L. Cattle temperament: Persistence of assessments and associations with productivity, efficiency, carcass and meat quality traits. J. Anim. Sci. 2011, 89, 1452–1465. [Google Scholar] [CrossRef]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar] [PubMed]
- Braz, K.M.G.; Monteiro, F.M.; Fernandes, L.G.; Rodrigues, N.N.; Peixoto Jr, K.C.; Green, R.E.; Cortez, A.; Crespilho, A.M. Does bull temperament impact growth performance and semen quality? Livest. Sci. 2020, 236, 104038. [Google Scholar] [CrossRef]
- Est’evez-Moreno, L.X.; Fl’orez-Díaz, H.; Duque-Muñoz, L.G.; Villarroel, M.; la Lama, G.C.M. Influence of temperament on performance and carcass quality of commercial Brahman steers in a Colombian tropical grazing system. Animals 2022, 118, 108867. [Google Scholar]
- Petherick, J.C.; Holroyd, R.G.; Doogan, V.J.; Venus, B.K. Productivity, carcass and meat quality of lot-fed Bos indicus cross steers grouped according to temperament. Aust. J. Exp. Agric. 2002, 42, 389–398. [Google Scholar]
- Odore, R.; Biasato, I.; Gardini, G.; D’Angelo, A.; Bellino, C. Effects of compost-bedded pack barn on circulating cortisol and Beta-endorphins in dairy cows: A case study. Animals 2021, 11, 3318. [Google Scholar] [CrossRef]
- Le Neindre, P.; Trillat, G.; Sapa, J.; Menissier, F.; Bonnet, J.N.; Chupin, J.M. Individual differences in docility of Limousin cattle. J. Anim. Sci. 1995, 73, 2249–2253. [Google Scholar]
- Moberg, G.P. Biological Response to Stress: Implications for Animal Welfare. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; Moberg, G.P., Mench, J.A., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 1–21. 377p. [Google Scholar]
- Galhardo, L.; Vital, J.; Oliveira, R.F. The role of predictability in the stress response of a cichlid fish. Physiol. Behav. 2011, 102, 367–372. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
| Sampling Time | 55 km | 2200 km | 15 Days |
|---|---|---|---|
| Calm | −12% | +60% | −4% |
| Reactive | +23% | +49% | −6% |
| All bulls | +6% | +54% | −5% |
| Sampling Time | Basal | Arrival | 15 Days |
|---|---|---|---|
| Calm | 530.40 ± 39.71 A | 488.20 ± 20.48 B | 533.30 ± 22.90 A |
| Reactive | 501.28 ± 28.04 AB | 447.46 ± 18.85 C | 504.38 ± 25.70 AB |
| All bulls | 513.94 ± 35.94 A | 465.17 ± 28.14 B | 516.96 ± 28.10 A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazio, E.; Cavaleri, S.; Medica, P.; Cravana, C.; La Fauci, D. Effects of Short- and Long-Distance Road Transport and Temperament on the Magnitude of β-Endorphin Response in Limousine Bulls. Ruminants 2025, 5, 11. https://doi.org/10.3390/ruminants5020011
Fazio E, Cavaleri S, Medica P, Cravana C, La Fauci D. Effects of Short- and Long-Distance Road Transport and Temperament on the Magnitude of β-Endorphin Response in Limousine Bulls. Ruminants. 2025; 5(2):11. https://doi.org/10.3390/ruminants5020011
Chicago/Turabian StyleFazio, Esterina, Salvatore Cavaleri, Pietro Medica, Cristina Cravana, and Deborah La Fauci. 2025. "Effects of Short- and Long-Distance Road Transport and Temperament on the Magnitude of β-Endorphin Response in Limousine Bulls" Ruminants 5, no. 2: 11. https://doi.org/10.3390/ruminants5020011
APA StyleFazio, E., Cavaleri, S., Medica, P., Cravana, C., & La Fauci, D. (2025). Effects of Short- and Long-Distance Road Transport and Temperament on the Magnitude of β-Endorphin Response in Limousine Bulls. Ruminants, 5(2), 11. https://doi.org/10.3390/ruminants5020011