

Factors Affecting Antibody-Mediated Immune Response and Cellular-Mediated Immune Response in Weaned Brahman Calves


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
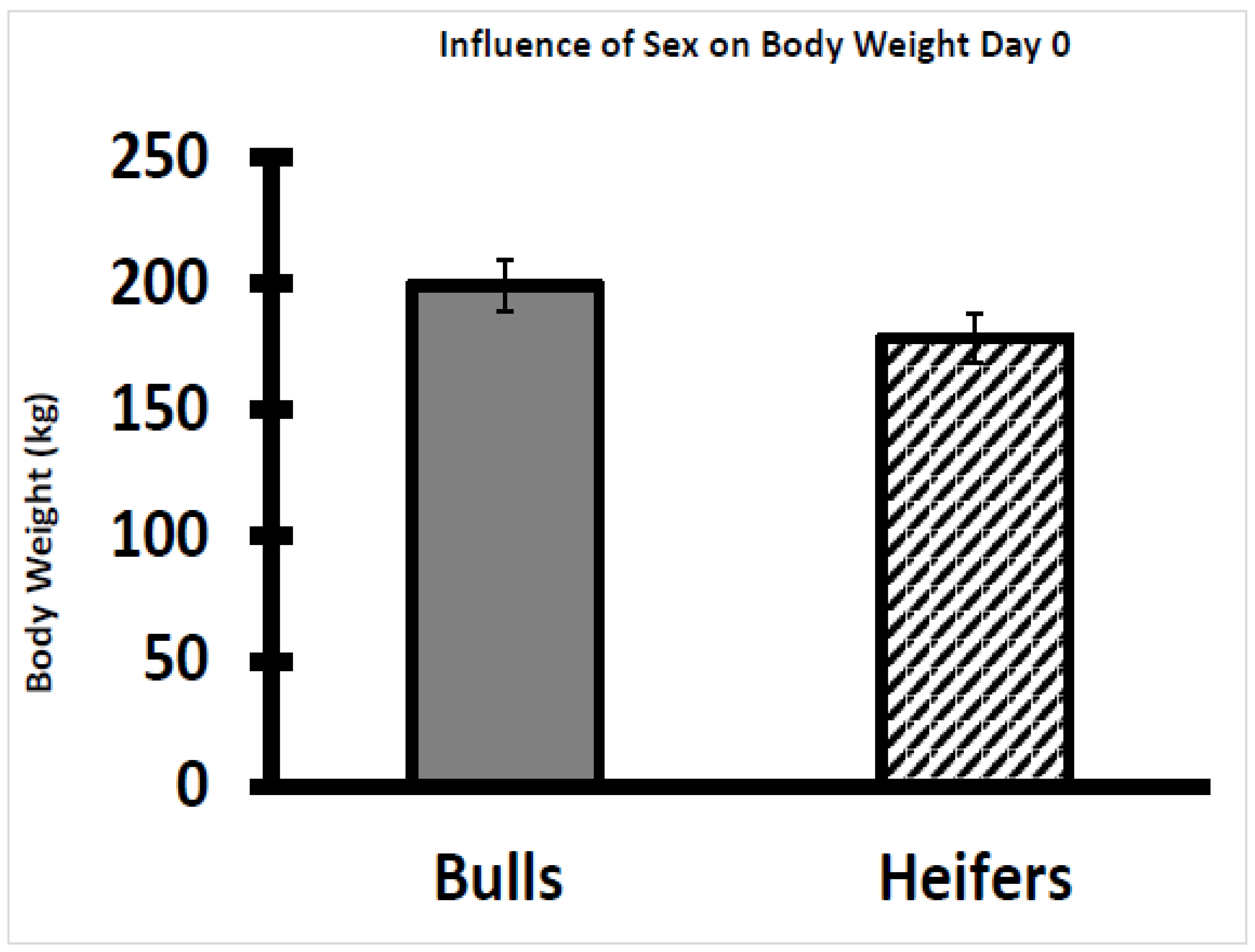
2.1. Experimental Design
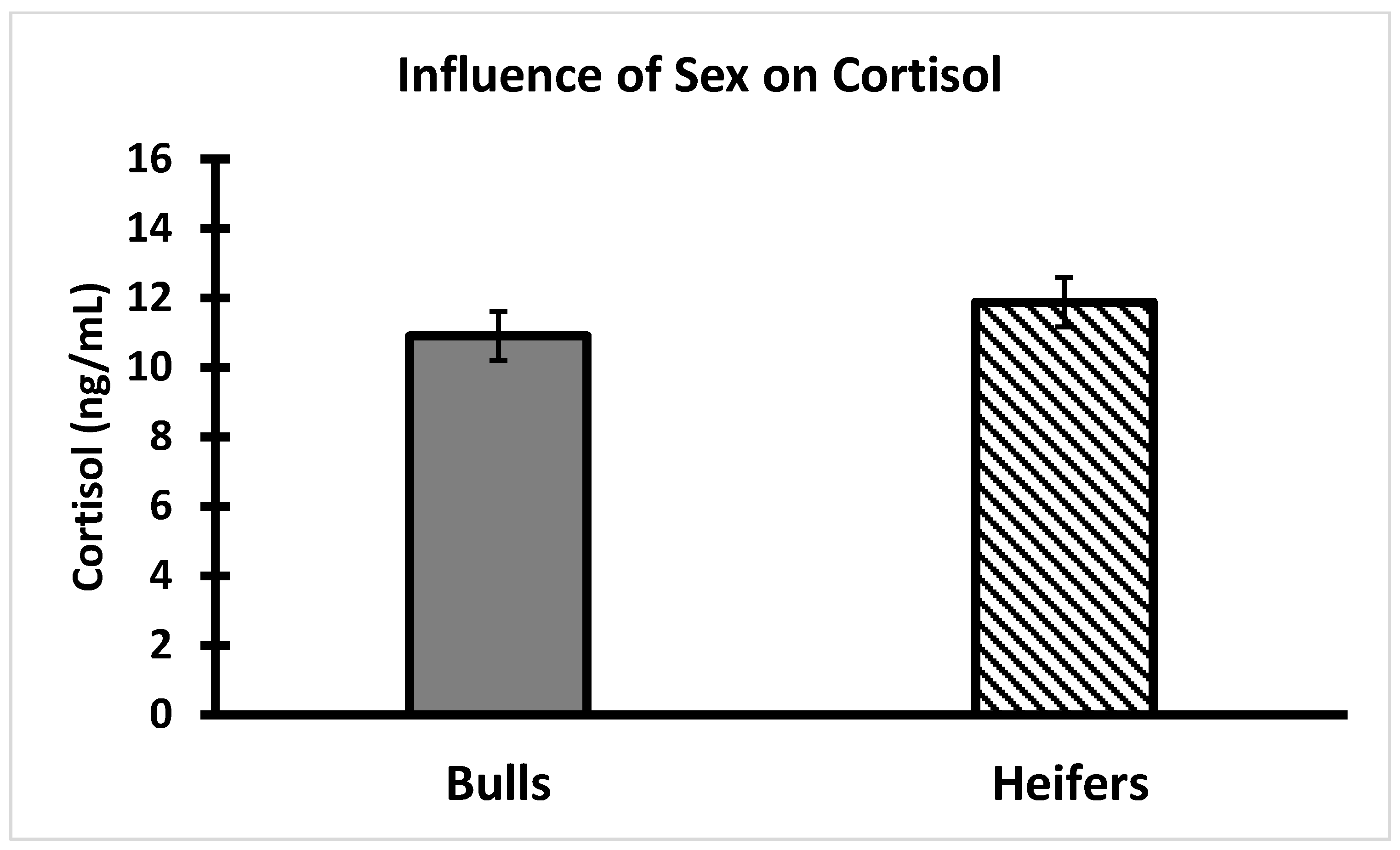
2.2. Cortisol
2.3. Evaluation of AMIR
2.4. Evaluation of CMIR
2.5. Statistical Analysis
3. Results and Discussion
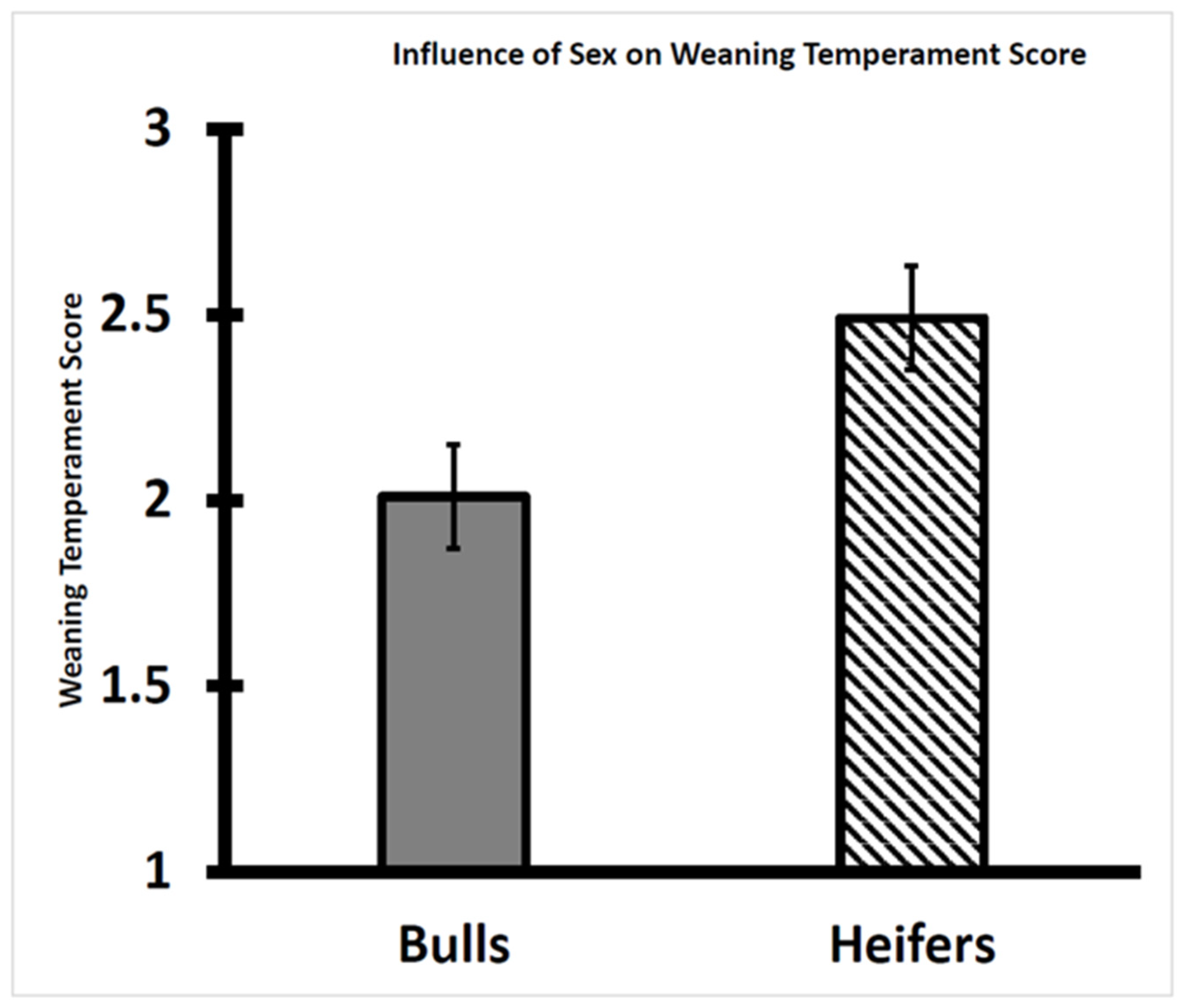
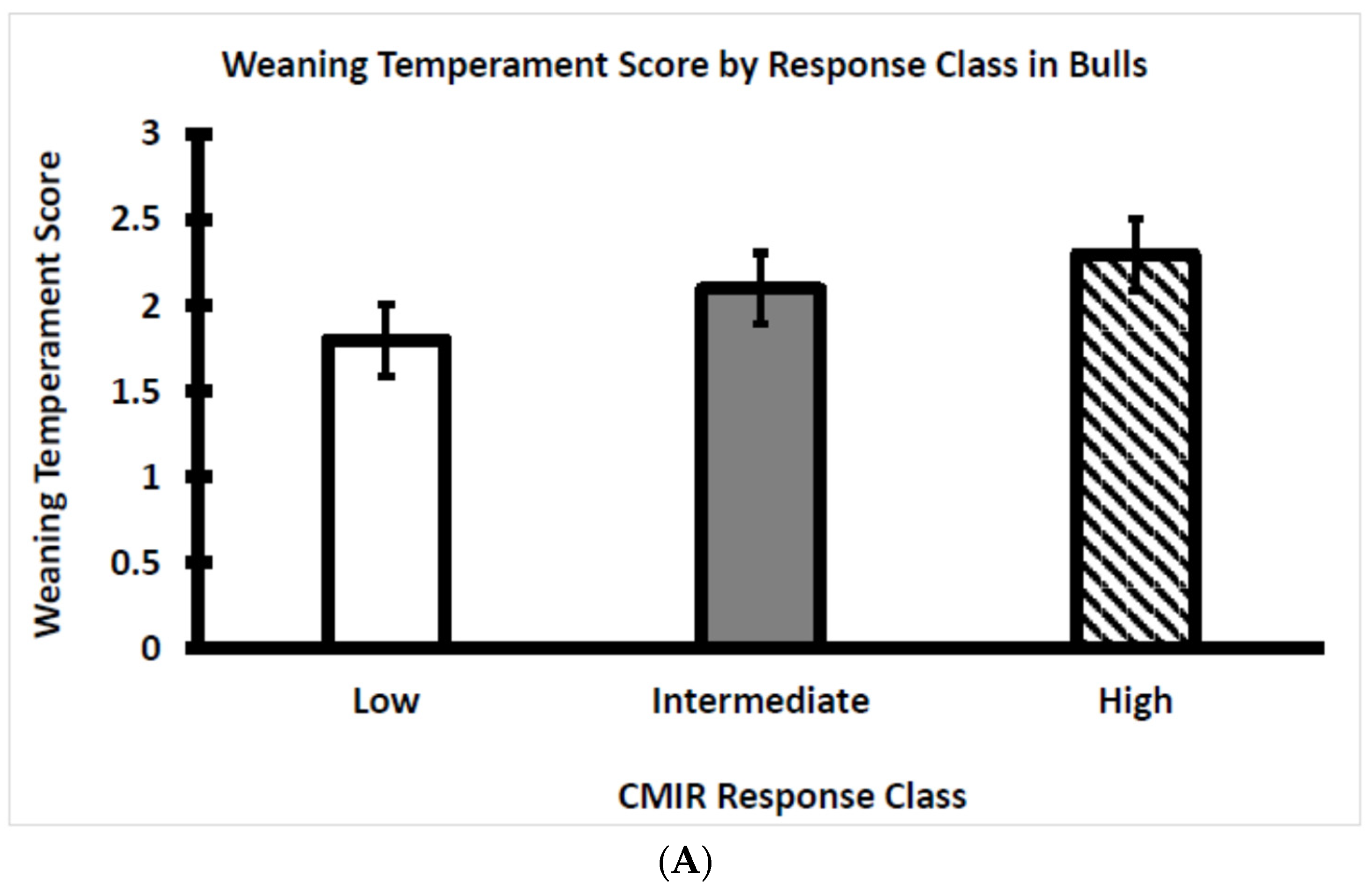
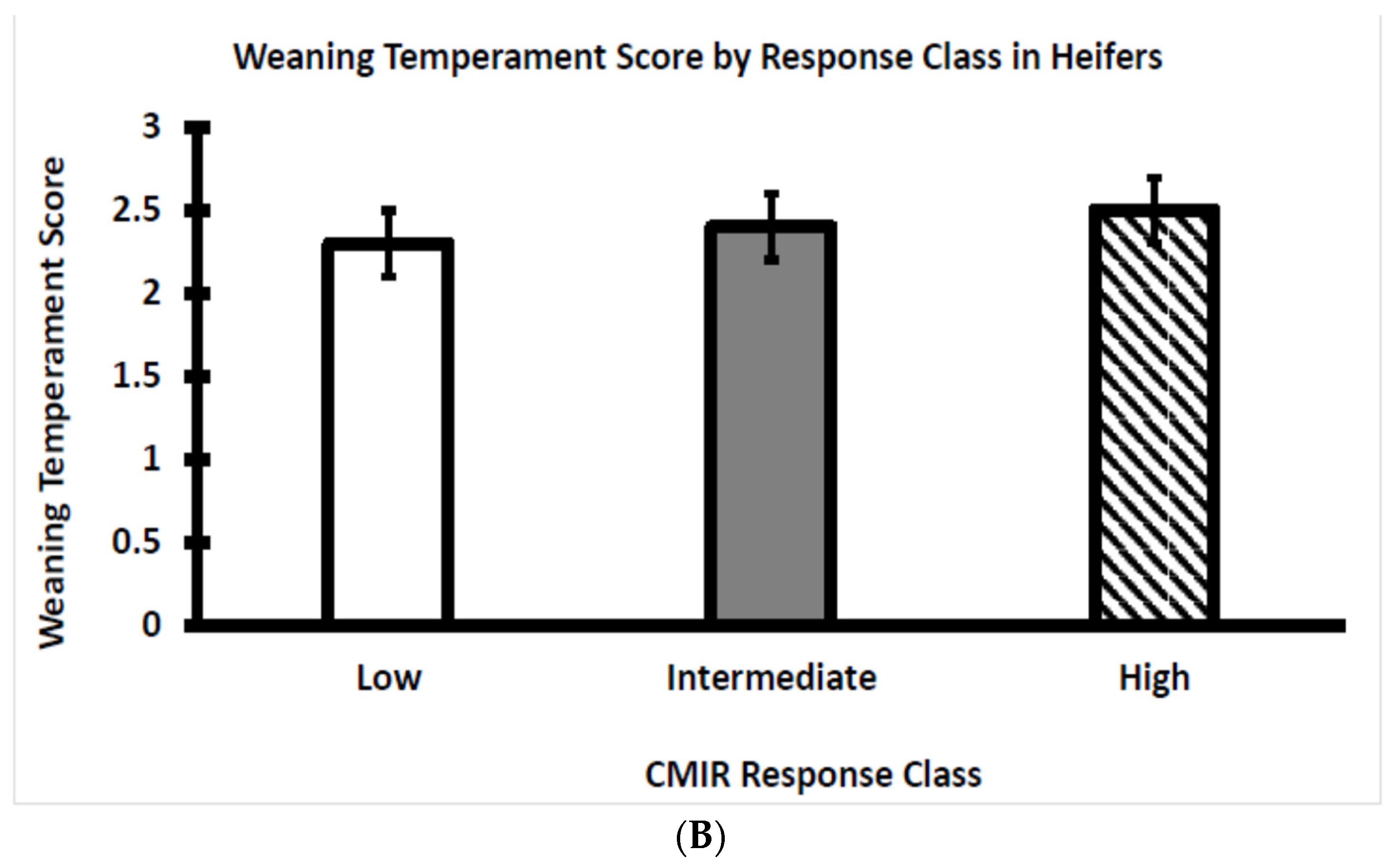
3.1. Temperament Score at Weaning Differed between Bull and Heifer Calves
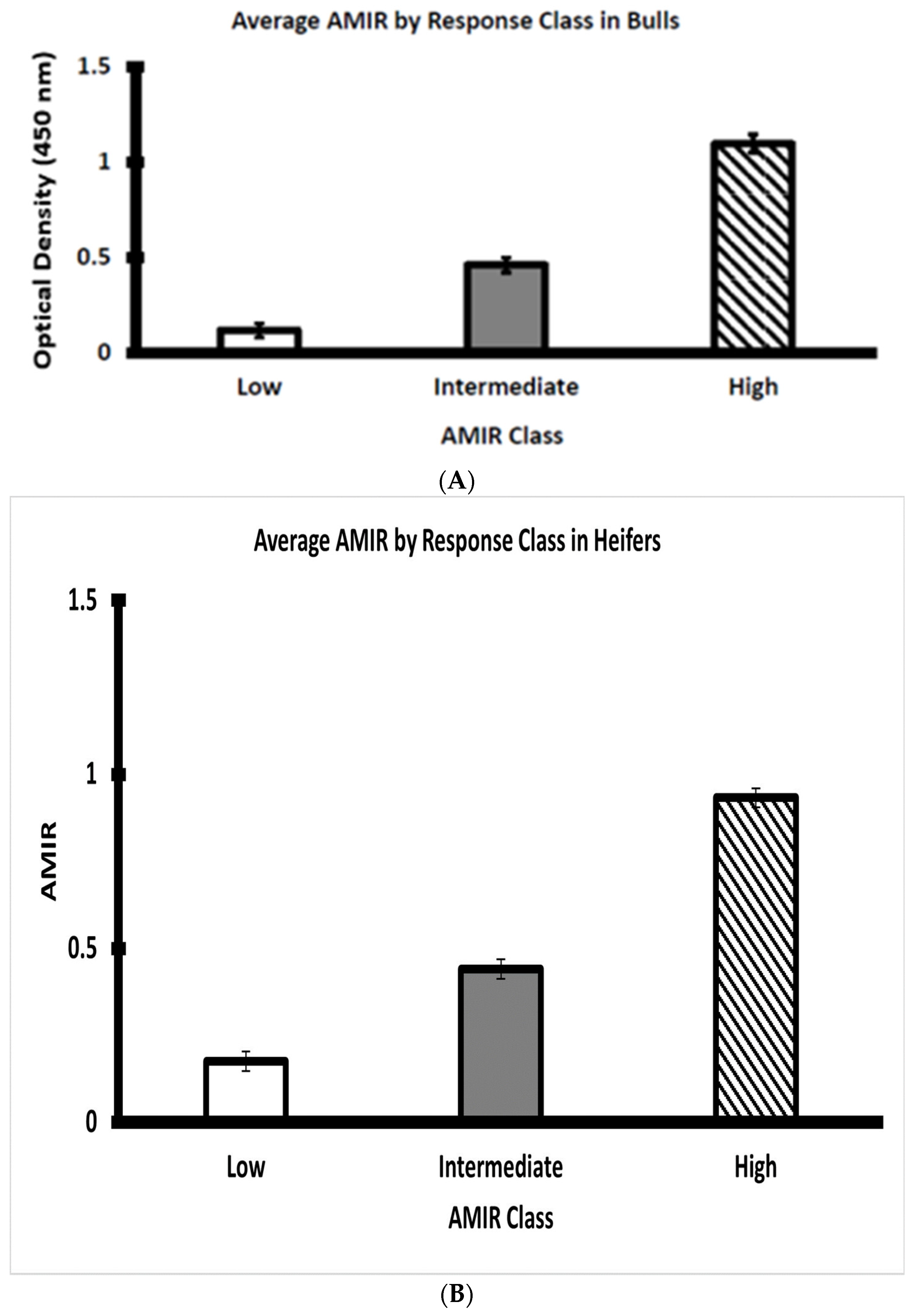
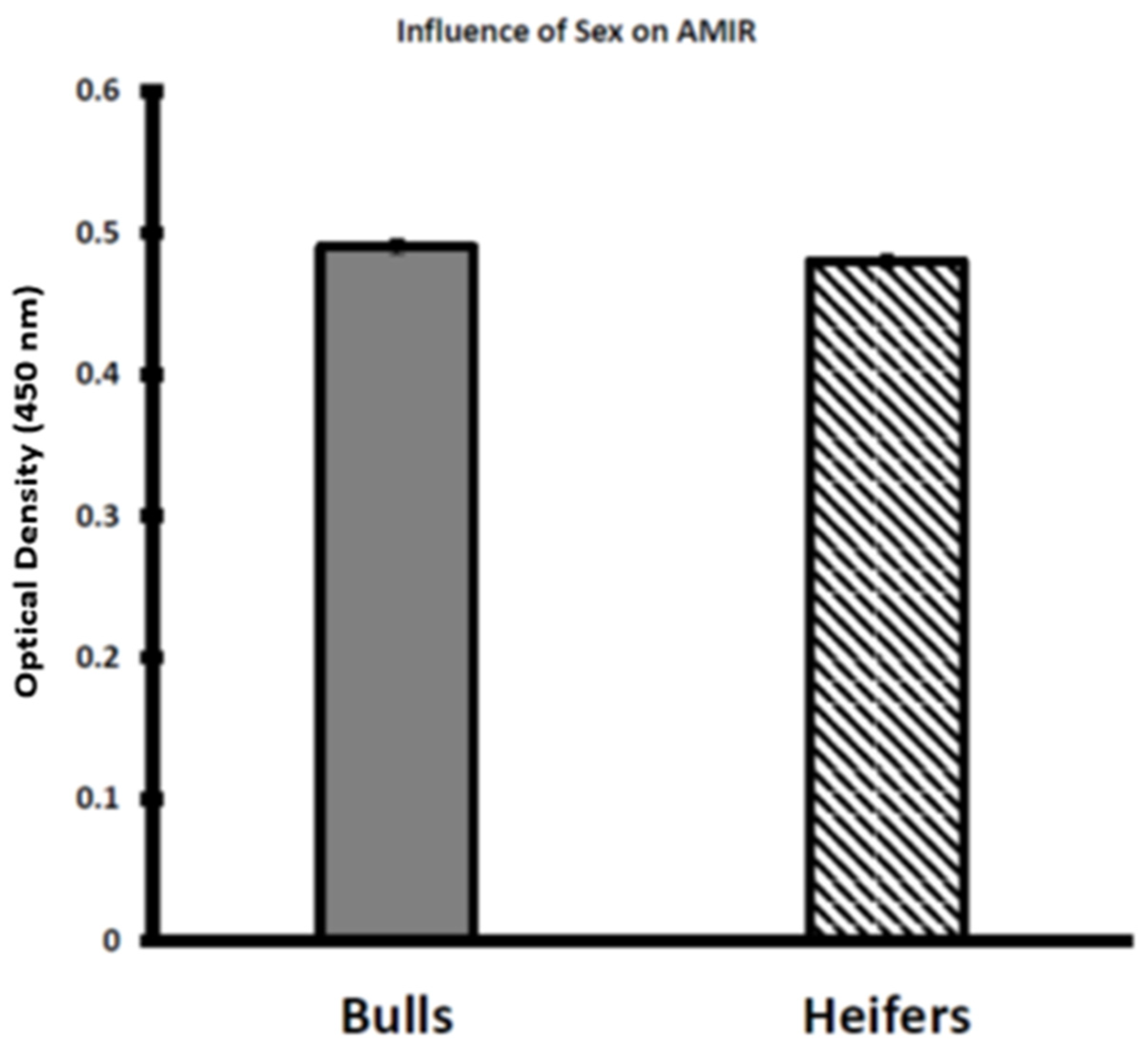
3.2. Assessment of AMIR in Bull and Heifer Calves
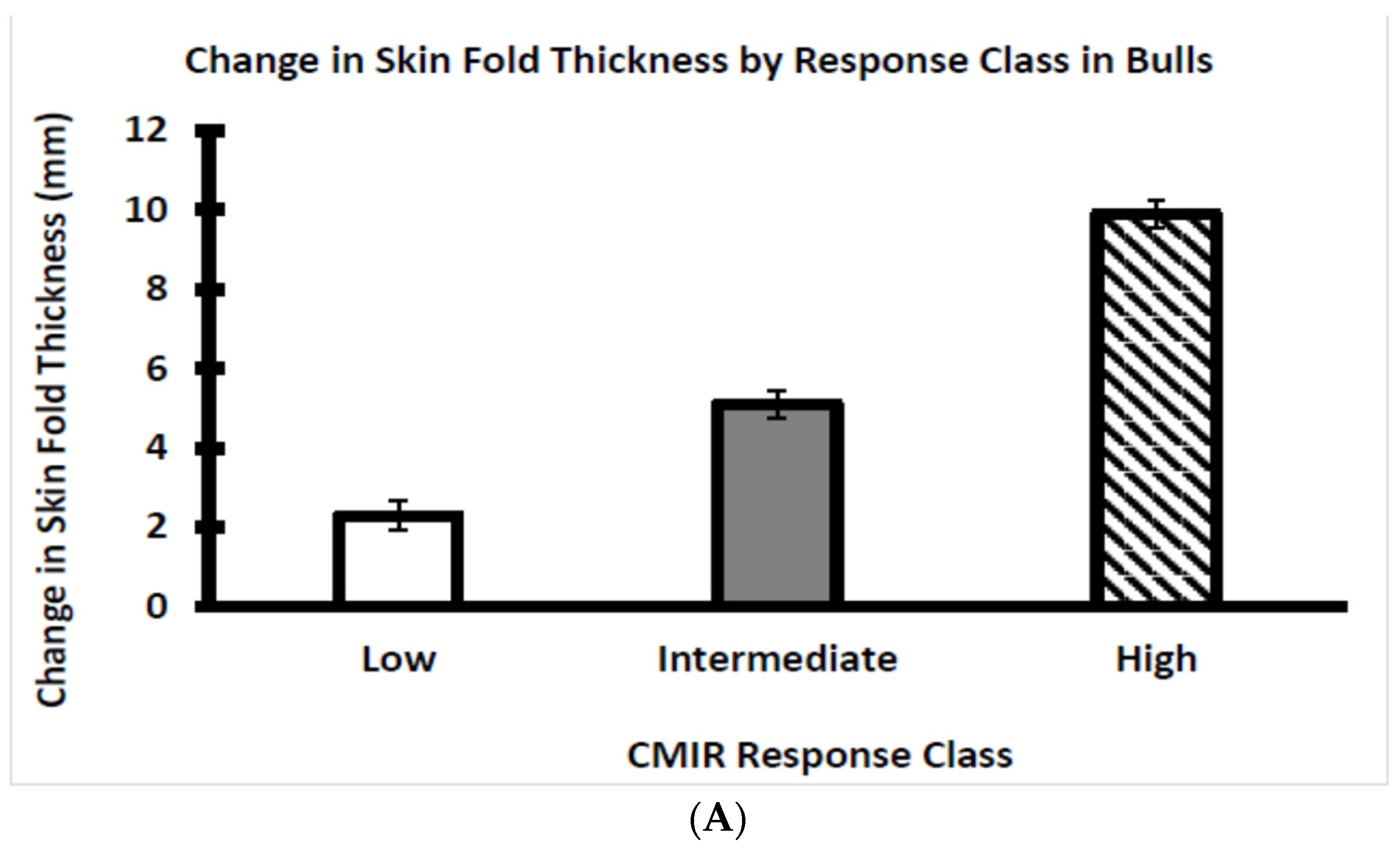
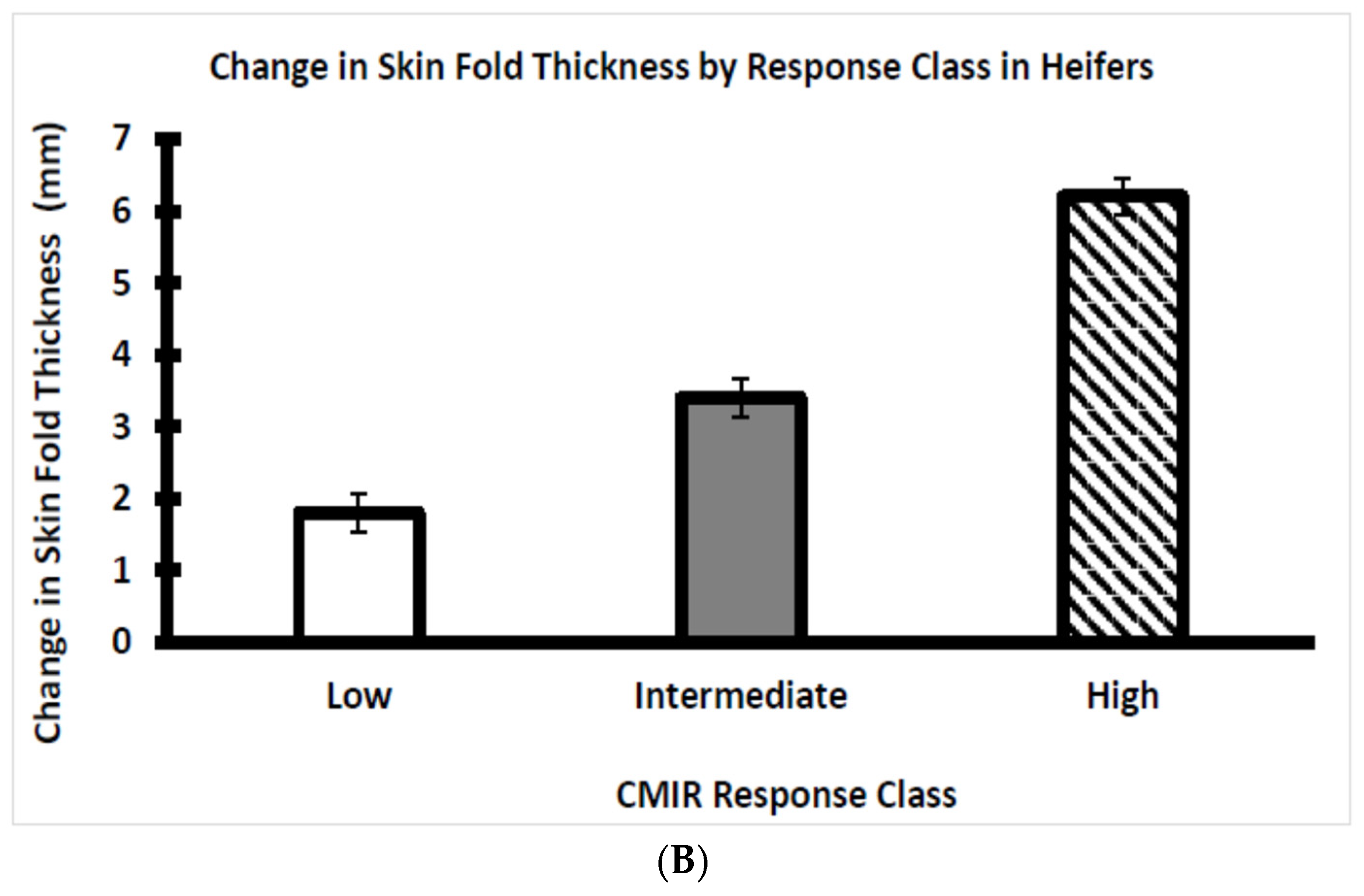
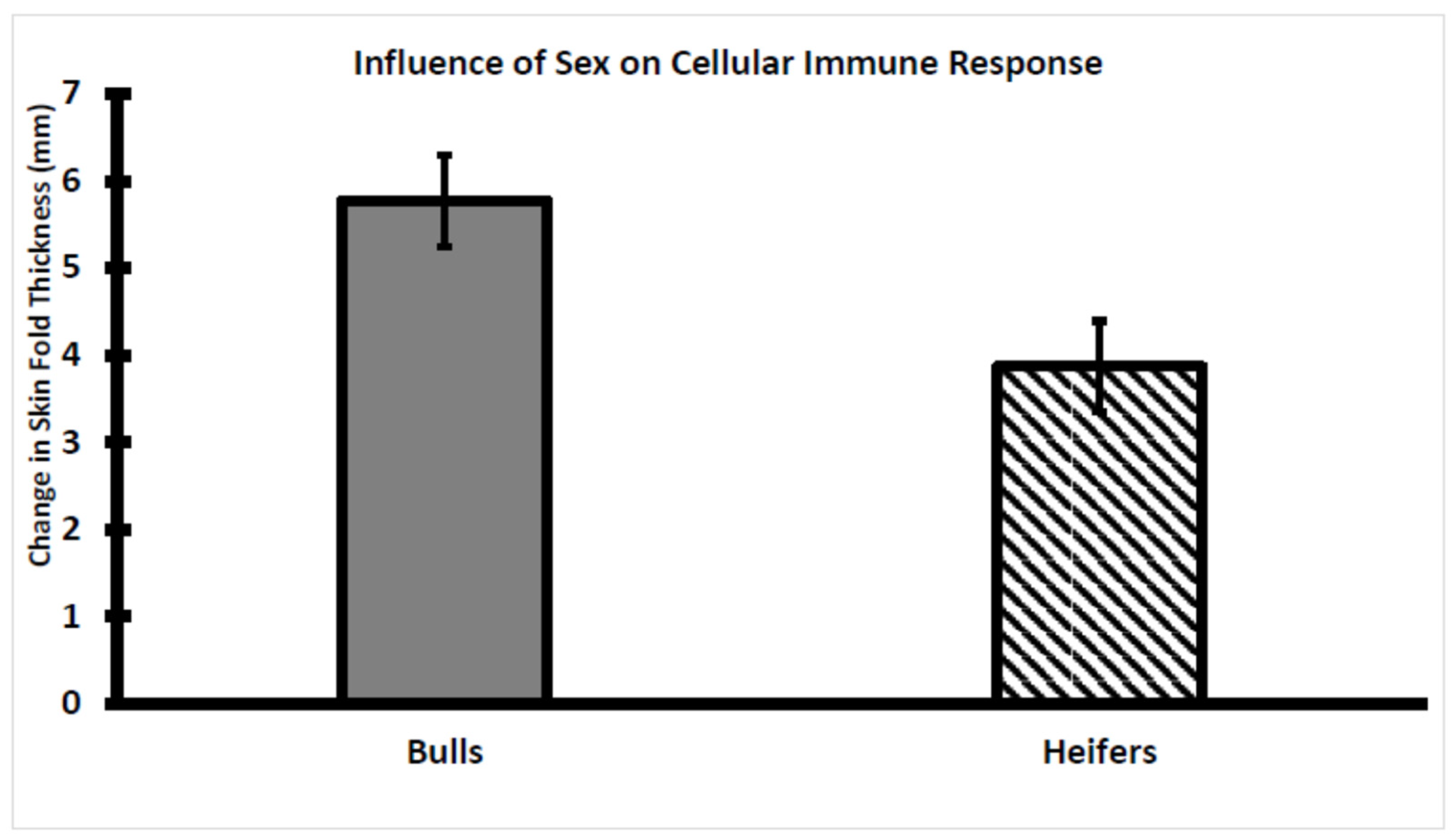
3.3. Assessment of CMIR in Bull and Heifer Calves
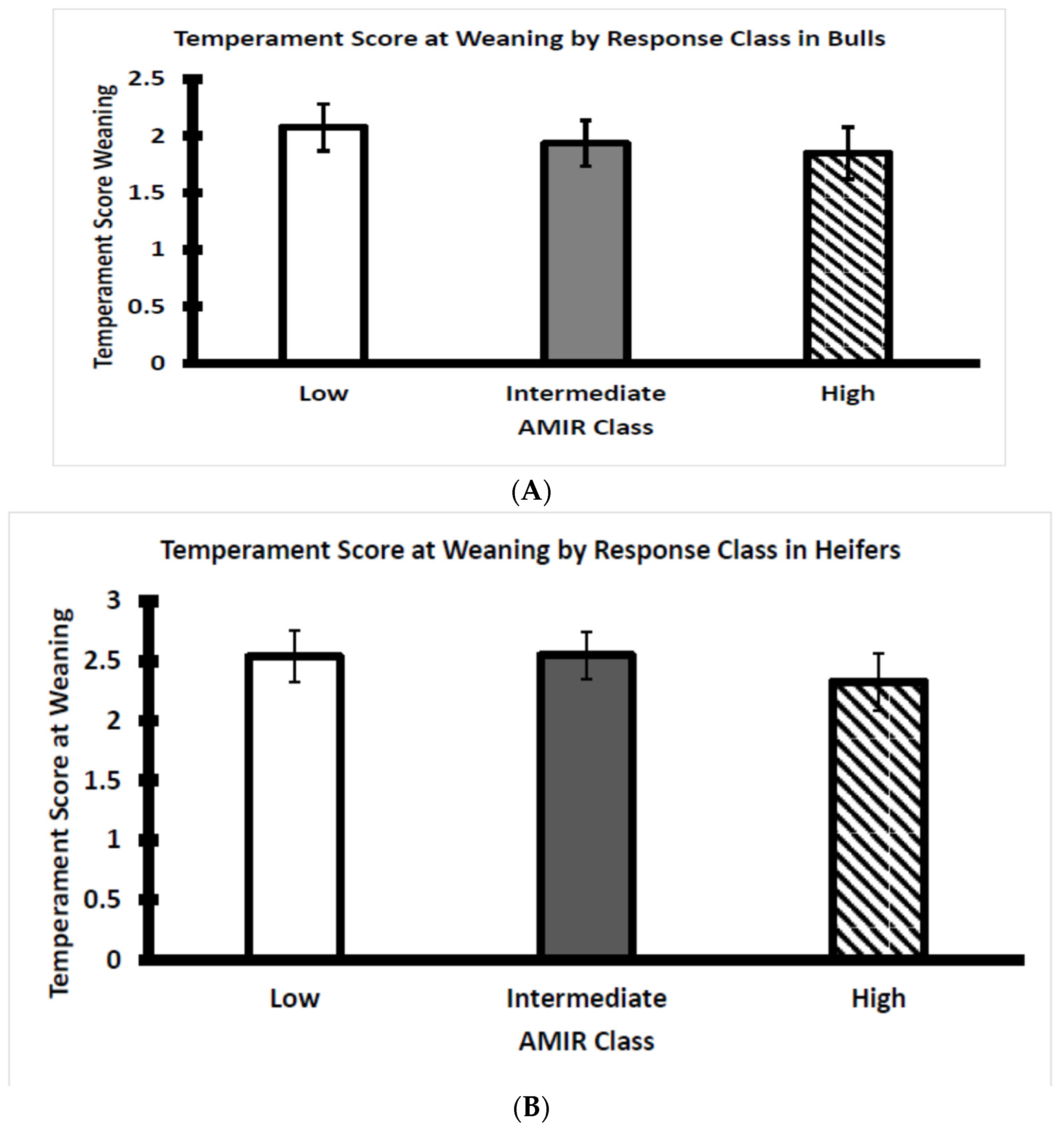
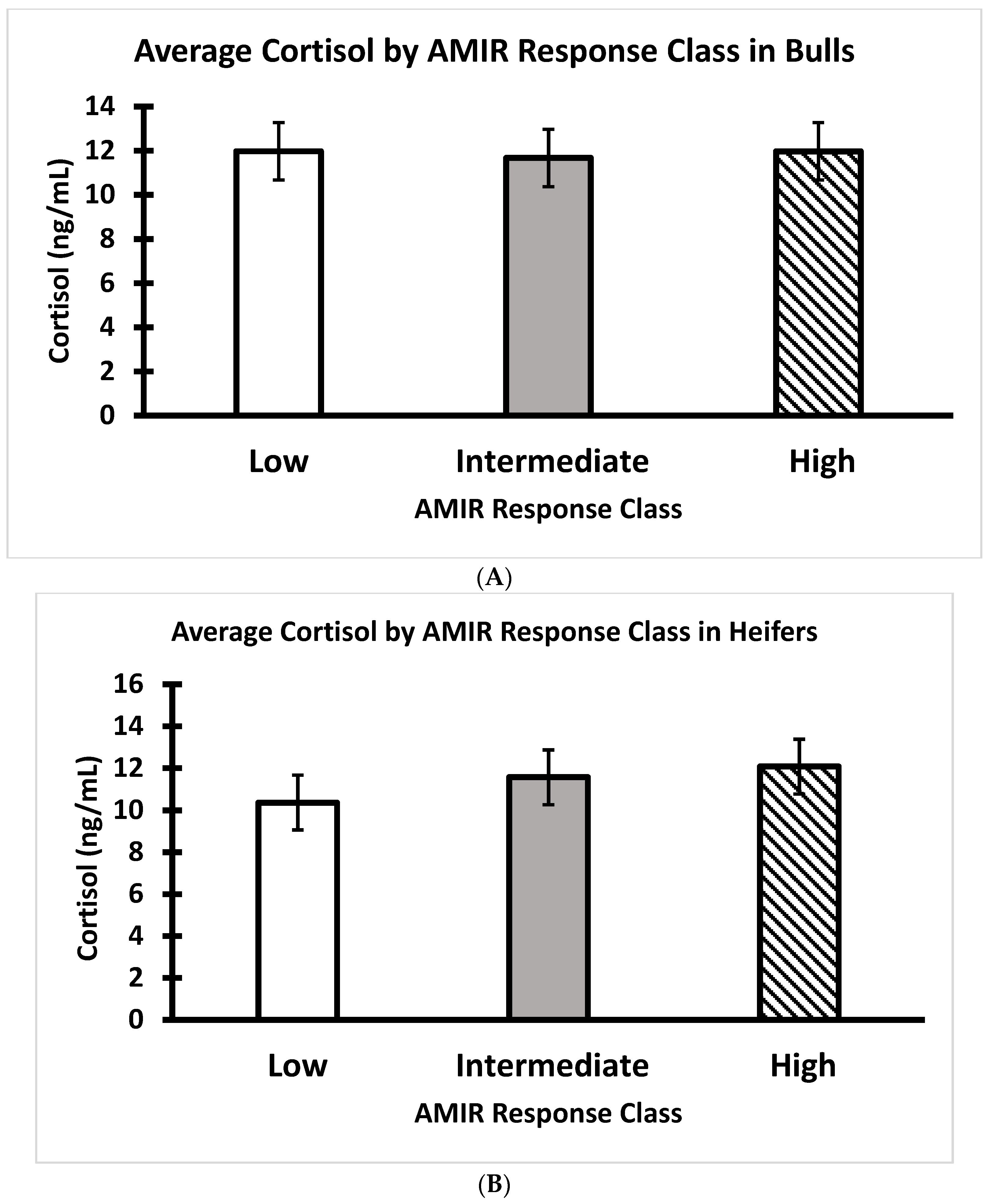
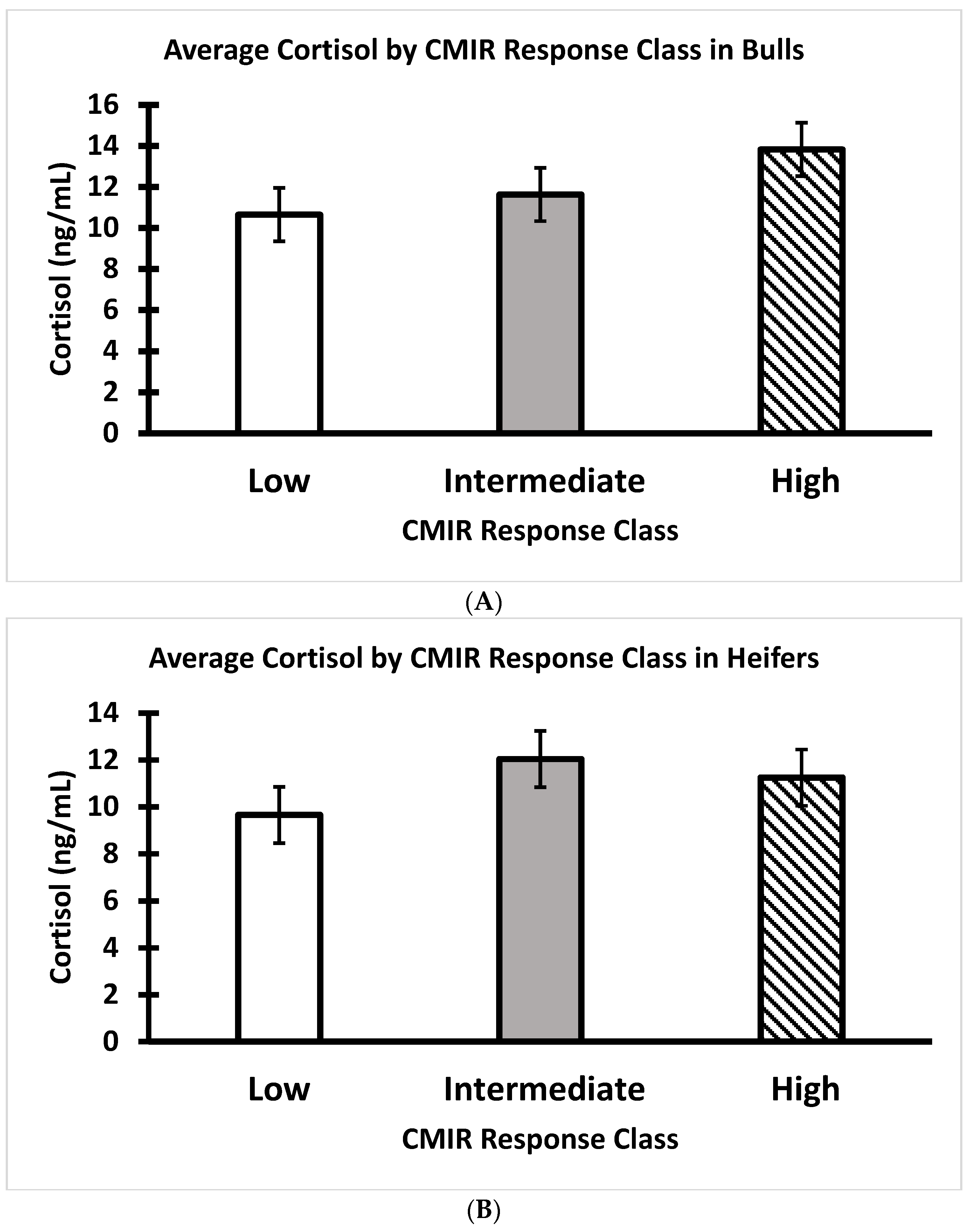
3.4. AMIR and CMIR in Relation to Body Condition Score, Cortisol, and Temperament
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Emmam, M.; Livernois, A.; Paibomesai, M.; Atalla, H.; Mallard, B. Genetic and epigenetic regulation of immune response and resistance to infectious diseases in domestic ruminants. Vet. Clin. Food Anim. 2019, 35, 405–429. [Google Scholar] [CrossRef]
- Tizard, I.R. Veterinary Immunology: An Introduction, 10th ed.; Elsevier/Saunders: Philadelphia, PA, USA, 2016; ISBN 9780323523493. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Basic Immunology: Functions and Disorders of the Immune System, 6th ed.; Elsevier/Saunders: Philadelphia, PA, USA, 2019; ISBN 9788131259573. [Google Scholar]
- Aleri, J.W.; Hine, B.C.; Pyman, M.F.; Mansell, P.D.; Wales, W.J.; Mallard, B.; Fisher, A.D. Assessing adaptive immune response phenotypes in Australian Holstein-Friesian heifers in a pasture-based production system. J. Anim. Sci. 2015, 93, 3713–3721. [Google Scholar]
- Thompson-Crispi, K.A.; Miglior, F.; Mallard, B.A. Incidence rates of clinical mastitis among Canadian Holsteins classified as high, average, or low immune responders. Clin. Vaccine Immunol. 2013, 20, 106–112. [Google Scholar] [CrossRef]
- Hines, M.T.; Ii, H.C.S.; Bayly, W.M.; Leroux, A.J. Exercise and immunity: A review with emphasis on the horse. J. Veter Intern. Med. 1996, 10, 280–289. [Google Scholar] [CrossRef]
- Aleri, J.; Hine, B.; Pyman, M.; Mansell, P.; Wales, W.; Mallard, B.; Stevenson, M.; Fisher, A. Associations between immune competence, stress responsiveness, and production in Holstein-Friesian and Holstein-Friesian × Jersey heifers reared in a pasture-based production system in Australia. J. Dairy Sci. 2019, 102, 3282–3294. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, G.; Dodt, R.; Wythes, J. Cattle temperaments in extensive beef herds in northern Queensland: Factors affecting temperament. Aust. J. Exp. Agric. 1988, 28, 683–687. [Google Scholar] [CrossRef]
- Fell, L.R.; Colditz, I.G.; Walker, K.H.; Watson, D.L. Associations between temperament, performance and immune function in cattle entering a commercial feedlot. Aust. J. Exp. Agric. 1999, 39, 795–802. [Google Scholar] [CrossRef]
- Texas Cooperative Extension. Value Added Calf (VAC)–Vaccination Programs. 2005. Available online: https:animalscience.tamu.edu/wp-content/uploads/sites/14/2012/04/beef-vac-vaccination.pdf (accessed on 28 June 2023).
- Richeson, J.T.; Beck, P.A.; Gadberry, M.S.; Gunter, S.A.; Hess, T.W.; Hubbell, D.S.; Jones, C. Effects of on-arrival versus delayed modified live virus vaccination on health, performance, and serum infectious bovine rhinotracheitis titers of newly received beef calves. J. Anim. Sci. 2008, 86, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Galyean, M.L.; Duff, G.C.; Rivera, J.D. Galyean appreciation club review: Revisiting nutrition and health of newly received cattle—What have we learned in the last 15 years? J. Anim. Sci. 2022, 100, skac067. [Google Scholar] [CrossRef] [PubMed]
- Matty, J.M.; Reddout, C.; Adams, J.; Major, M.; Lalman, D.; Biggs, R.; Salak-Johnson, J.L.; Beck, P.A. The effects of respiratory vaccine type and timing on antibody titers, immunoglobulins, and growth performance in pre- and post-weaned beef calves. Vet. Sci. 2023, 10, 37. [Google Scholar] [CrossRef]
- FASS. Guide for Care and Use of Agricultural Animals in Research and Teaching, 3rd ed.; Federation of Animal Science Societies: Champaign, IL, USA, 2010. [Google Scholar]
- Curley, K.O., Jr.; Paschal, J.C.; Welsh, T.H., Jr.; Randel, R.D. Exit velocity as a measurement of cattle temperament is repeatable and associated with serum concentration of cortisol in Brahman bulls. J. Anim. Sci. 2006, 84, 3100–3103. [Google Scholar] [CrossRef] [PubMed]
- King, D.; Pfeiffer, C.S.; Randel, R.; Welsh, T.; Oliphint, R.; Baird, B.; Curley, K.; Vann, R.; Hale, D.; Savell, J. Influence of animal temperament and stress responsiveness on the carcass quality and beef tenderness of feedlot cattle. Meat Sci. 2006, 74, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Burrow, H.M.; Seifert, G.W.; Corbet, N.J. A new technique for measuring temperament in cattle. Proc. Aust. Soc. Anim. Prod. 1988, 17, 154–157. [Google Scholar]
- Hammond, A.C.; Olson, T.A.; Chase, C.C., Jr.; Bowers, E.J.; Randel, R.D.; Murphy, C.N.; Vogt, D.W.; Tewolde, A. Heat tolerance in two tropically adapted Bos taurus breeds, Senepol and Romosinuano, compared with Brahman, Angus, and Hereford cattle in Florida. J. Anim. Sci. 1996, 74, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, S.; Begley, N.; Schaeffer, L.; Burnside, E.; Mallard, B. Antibody and cell-mediated immune responses and survival between Holstein and Norwegian Red x Holstein Canadian calves. J. Dairy Sci. 2011, 94, 1576–1585. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Quinton, M.; Miglior, F.; Mallard, B.A. Genetic parameters of dairy cattle immune response traits. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Minas Gerais, Brazil, WCGALP, Belo Horizonte, Brazil, 13–18 August 2006; pp. 15–18. [Google Scholar]
- Littlejohn, B.P.; Price, D.M.; Banta, J.P.; Lewis, A.W.; Neuendorff, D.A.; Carroll, J.A.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Prenatal transportation stress alters temperament and serum cortisol concentrations in suckling Brahman calves. J. Anim. Sci. 2016, 94, 602–609. [Google Scholar] [CrossRef]
- Hermesch, D.R.; Thomson, D.U.; Loneragan, G.H.; Renter, D.R.; White, B.J. Effects of a commercially available vaccine against Salmonella enterica serotype Newport on milk production, somatic cell count and shedding of Salmonella organisms in female dairy cattle with no clinical signs of salmonellosis. Am. J. Ver. Res. 2008, 69, 1229–1234. [Google Scholar] [CrossRef]
- SAS. SAS/STAT User’s Guide, Version 9.3; SAS Institute: Cary, NC, USA, 2011. [Google Scholar]
- Voisinet, B.D.; Grandin, T.; Tatum, J.D.; O’Connor, S.F.; Struthers, J.J. Feedlot cattle with calm temperaments have higher average daily gains than cattle with excitable temperaments. J. Anim. Sci. 1997, 75, 892–896. [Google Scholar] [CrossRef]
- Shrode, R.R.; Hammack, S.P. Chute behavior of yearling beef cattle. J. Anim. Sci. 1971, 33, 193. [Google Scholar]
- Tennessen, T.; Price, M.A.; Berg, R.T. Comparative responses of bulls and steers to transportation. Can. J. Anim. Sci. 1984, 64, 333–338. [Google Scholar] [CrossRef]
- Lay, D.C., Jr.; Friend, T.H.; Grissom, K.K.; Bowers, C.L.; Mal, M.E. Effects of freeze or hot-iron branding of Angus calves on some physiological and behavioral indicators of stress. Appl. Anim. Behav. Sci. 1992, 33, 137–147. [Google Scholar] [CrossRef]
- Hoppe, S.; Brandt, H.R.; König, S.; Erhardt, G.; Gauly, M. Temperament traits of beef calves measured under field conditions and their relationships to performance. J. Anim. Sci. 2010, 88, 1982–1989. [Google Scholar] [CrossRef]
- Heriazon, A.; Yager, J.A.; Sears, W.; Mallard, B.A. Induction of delayed-type hypersensitivity and interferon-gamma to Candida albicans and anti-hen-egg white lysozyme antibody as phenotypic markers of enhanced bovine immune response. Vet. Immunol. Immunopathol. 2009, 129, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Begley, N.; Buckley, F.; Burnside, E.; Schaeffer, L.; Pierce, K.; Mallard, B. Immune responses of Holstein and Norwegian Red × Holstein calves on Canadian dairy farms. J. Dairy Sci. 2009, 92, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Aleri, J.W.; Hine, B.C.; Pyman, M.F.; Mansell, P.D.; Wales, W.J.; Mallard, B.; Fisher, A.D. An assessment of immune and stress responsiveness in Holstein Friesian cows selected for high and low feed conversion efficiency. Anim. Prod. Sci. 2016, 57, 244–251. [Google Scholar] [CrossRef]
- Browning, R., Jr.; Leite-Browning, M.L.; Neuendorff, D.A.; Randel, R.D. Preweaning growth of Angus-(Bos taurus), Brahman-(Bos indicus), and Tuli-(Sanga) sired calves and reproductive performance of their Brahman dams. J. Anim. Sci. 1995, 73, 2558–2563. [Google Scholar] [CrossRef]
- Hulbert, L.E.; Carroll, J.A.; Ballou, M.A.; Burdick, N.C.; Dailey, J.W.; Caldwell, L.C.; Loyd, A.N.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Sexually dimorphic stress and pro-inflammatory cytokine responses to an intravenous corticotropin-releasing hormone challenge in Brahman cattle following transportation. Innate Immun. 2013, 19, 378–387. [Google Scholar] [CrossRef]
- Carroll, J.A.; Sanchez, N.C.B.; Hulbert, L.E.; Ballou, M.A.; Dailey, J.W.; Caldwell, L.C.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Sexually dimorphic innate immunological responses of pre-pubertal Brahman cattle following an intravenous lipopolysaccharide challenge. Vet. Immunol. Immunopathol. 2015, 166, 108–115. [Google Scholar] [CrossRef]
- Wilkie, B.; Mallard, B. Selection for high immune response: An alternative approach to animal health maintenance? Vet. Immunol. Immunopathol. 1999, 72, 231–235. [Google Scholar] [CrossRef]
- Romani, L. Innate and adaptive immunity in Candida albicans infections and saprophytism. J. Leukocyte Biol. 2000, 68, 175–179. [Google Scholar] [CrossRef]
- Chamorro, M.F.; Woolums, A.; Walz, P.H. Vaccination of calves against common respiratory viruses in the face of maternally derived antibodies (IFOMA). Anim. Health Res. Rev. 2016, 17, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.; Fulton, R.W.; Burge, L.J.; DuBois, W.R.; Payton, M. Passively transferred immunity in newborn calves, rate of antibody decay, and effect on subsequent vaccination with modified live virus vaccine. Bov. Pract. 2019, 35, 47–55. [Google Scholar] [CrossRef]
- Wagter, L.; Mallard, B.; Wilkie, B.; Leslie, K.; Boettcher, P.; Dekkers, J. A quantitative approach to classifying Holstein cows based on antibody responsiveness and its relationship to peripartum mastitis occurrence. J. Dairy Sci. 2000, 83, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Crispi, K.; Sewalem, A.; Miglior, F.; Mallard, B. Genetic parameters of adaptive immune response traits in Canadian Holsteins. J. Dairy Sci. 2012, 95, 401–409. [Google Scholar] [CrossRef]
- Biozzi, G.; Mouton, D.; Heumann, A.M.; Bouthillier, Y.; Stiffel, C.; Mevel, J.C. Genetic analysis of antibody responsiveness to sheep erythrocytes in crosses between lines of mice selected for high or low antibody synthesis. Am. J. Immunol. 1979, 36, 427–438. [Google Scholar]
- Mouton, D.; Bouthillier, Y.; Mevel, J.; Biozzi, G. Genetic selection for antibody responsiveness in mice: Further evidence for inverse modification of macrophage catabolic activity without alteration of the expression of T-cell-mediated immunity. Eur. J. Immunol. 1984, 135, 173–186. [Google Scholar] [CrossRef]
- Sarker, N.; Tsudzuki, M.; Nishibori, M.; Yamamoto, Y. Direct and correlated response to divergent selection for serum immunoglobulin M and G levels in chickens. Poult. Sci. 1999, 78, 1–7. [Google Scholar] [CrossRef]
- Abdel-Azim, G.; Freeman, A.; Kehrli, M., Jr.; Kelm, S.; Burton, J.; Kuck, A.; Schnell, S. Genetic basis and risk factors for infectious and noninfectious diseases in US Holsteins. J. Dairy Sci. 2005, 88, 1199–1207. [Google Scholar] [CrossRef]
- Ahuja, S.K.; Manoharan, M.S.; Lee, G.C.; McKinnon, L.R.; Meunier, J.A.; Steri, M.; Harper, N.; Fiorillo, E.; Smith, A.M.; Restrepo, M.I.; et al. Immune resilience despite inflammatory stress promotes longevity and favorable health outcomes including resistance to infection. Nat. Commun. 2023, 14, 3286. [Google Scholar] [CrossRef]
- Gifford, C.A.; Holland, B.P.; Mills, R.L.; Maxwell, C.L.; Farney, J.K.; Terrill, S.J.; Step, D.L.; Richards, C.J.; Robles, L.O.B.; Krehbiel, C.R. Growth and development symposium: Impacts of inflammation on cattle growth and carcass merit. J. Anim. Sci. 2012, 90, 1438–1451. [Google Scholar] [CrossRef]
- Cockrum, R.R.; Speidel, S.E.; Salak-Johnson, J.L.; Chase, C.C.L.; Peel, R.K.; Weaber, R.L.; Loneragan, G.H.; Wagner, J.J.; Boddhireddy, P.; Thomas, M.G.; et al. Genetic parameters estimated at receiving for circulating cortisol, immunoglobulin G, interleukin 8, and incidence of bovine respiratory disease in feedlot beef steers. J. Anim. Sci. 2016, 94, 2770–2778. [Google Scholar] [CrossRef] [PubMed]
- Husseini, N.; Beard, S.C.; Hodgins, D.C.; Barnes, C.; Chik, E.; Mallard, B.A. Immuno-phenotyping of Canadian beef cattle: Adaptation of the high immune response methodology for utilization in beef cattle. Transl. Anim. Sci. 2022, 6, txac006. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, C.L.; Riley, D.G.; Randel, R.D.; Welsh, T.H., Jr. Factors Affecting Antibody-Mediated Immune Response and Cellular-Mediated Immune Response in Weaned Brahman Calves. Ruminants 2023, 3, 385-400. https://doi.org/10.3390/ruminants3040032
Young CL, Riley DG, Randel RD, Welsh TH Jr. Factors Affecting Antibody-Mediated Immune Response and Cellular-Mediated Immune Response in Weaned Brahman Calves. Ruminants. 2023; 3(4):385-400. https://doi.org/10.3390/ruminants3040032
Chicago/Turabian StyleYoung, Christian L., David G. Riley, Ronald D. Randel, and Thomas H. Welsh, Jr. 2023. "Factors Affecting Antibody-Mediated Immune Response and Cellular-Mediated Immune Response in Weaned Brahman Calves" Ruminants 3, no. 4: 385-400. https://doi.org/10.3390/ruminants3040032
APA StyleYoung, C. L., Riley, D. G., Randel, R. D., & Welsh, T. H., Jr. (2023). Factors Affecting Antibody-Mediated Immune Response and Cellular-Mediated Immune Response in Weaned Brahman Calves. Ruminants, 3(4), 385-400. https://doi.org/10.3390/ruminants3040032