
The Dynamics of Chromatin Accessibility Prompted by Butyrate-Induced Chromatin Modification in Bovine Cells

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. MDBK Cells and Butyrate Treatment
2.2. ATAC-Seq Library Preparation and Sequencing
2.3. ATAC-Seq Data Processing and Mapping
2.4. ATAC-Seq Peak Calling and Quality Check
2.5. Identification of Differentially Accessible Regions (DARs)
2.6. Annotation of DARs
2.7. Gene Ontology and Pathway Analysis of DARs
2.8. Motif Enrichment Analysis
2.9. Gene Co-Expression Network Analysis
3. Results
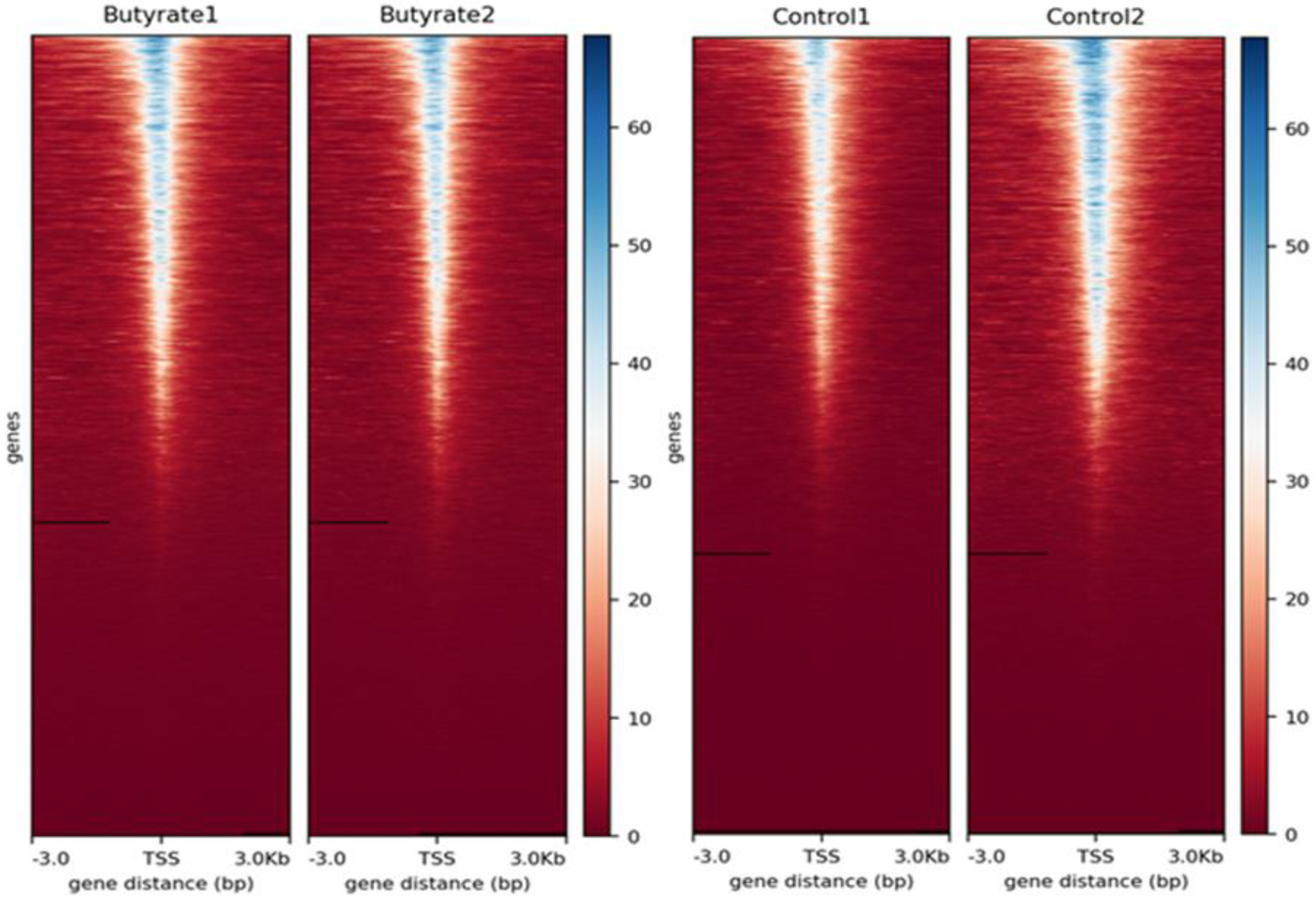
3.1. Read Quality, Alignment, and Fragment Size Distribution
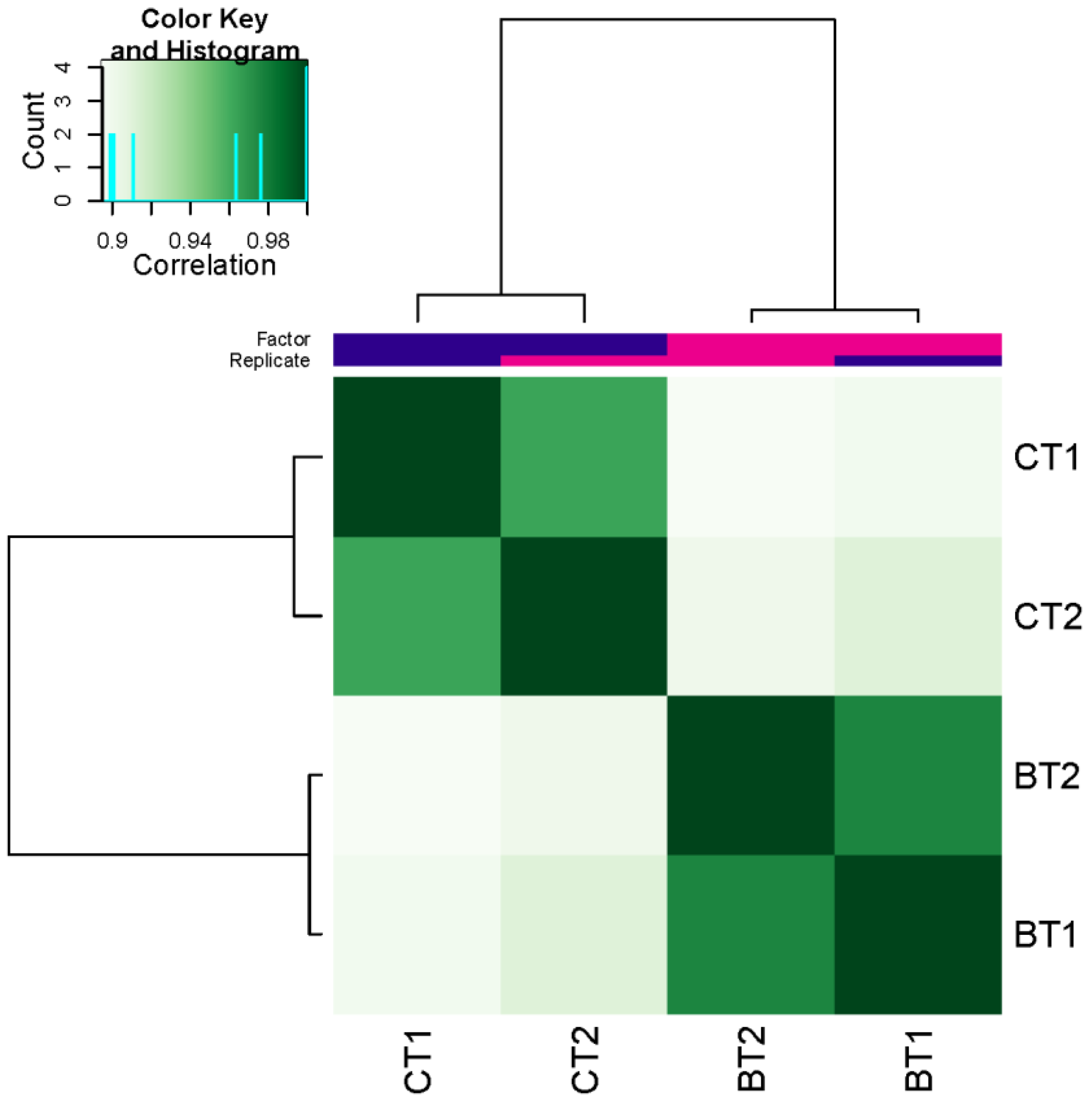
3.2. Identification of Accessible Chromatin Regions
3.3. Differentially Accessible Regions (DARs)
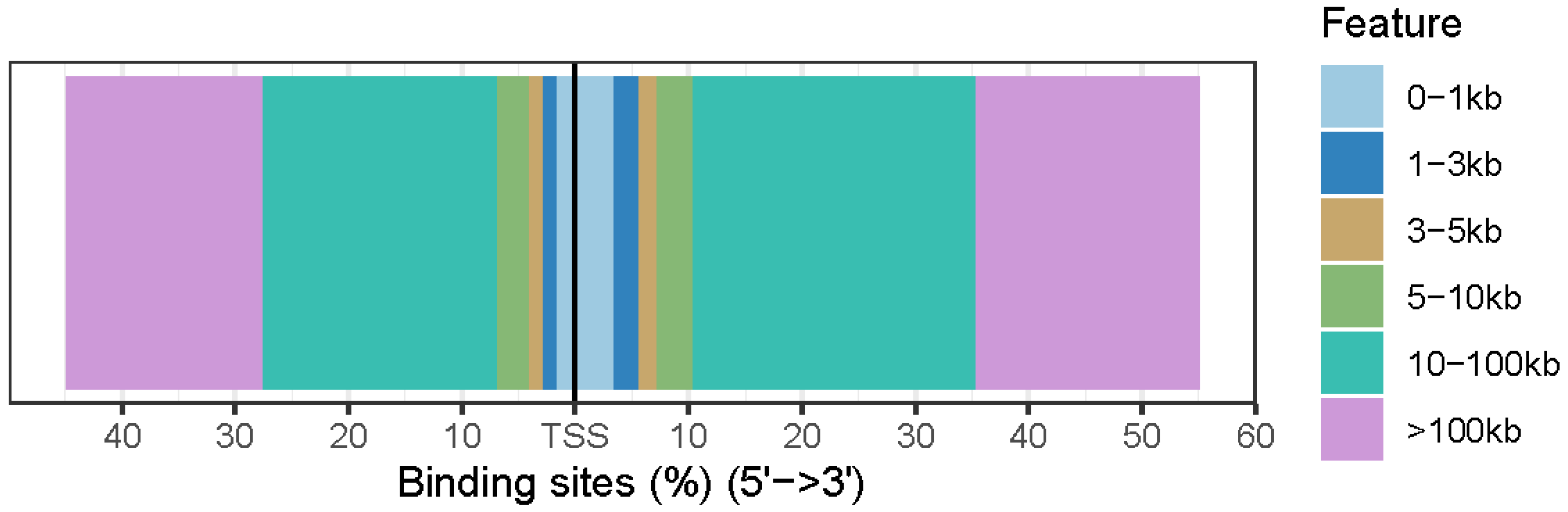
3.4. Annotation of Differentially Accessible Regions
3.5. Gene Ontology Enrichment Analysis of Differentially Accessible Regions
3.6. Pathway Analysis of Differentially Accessible Regions
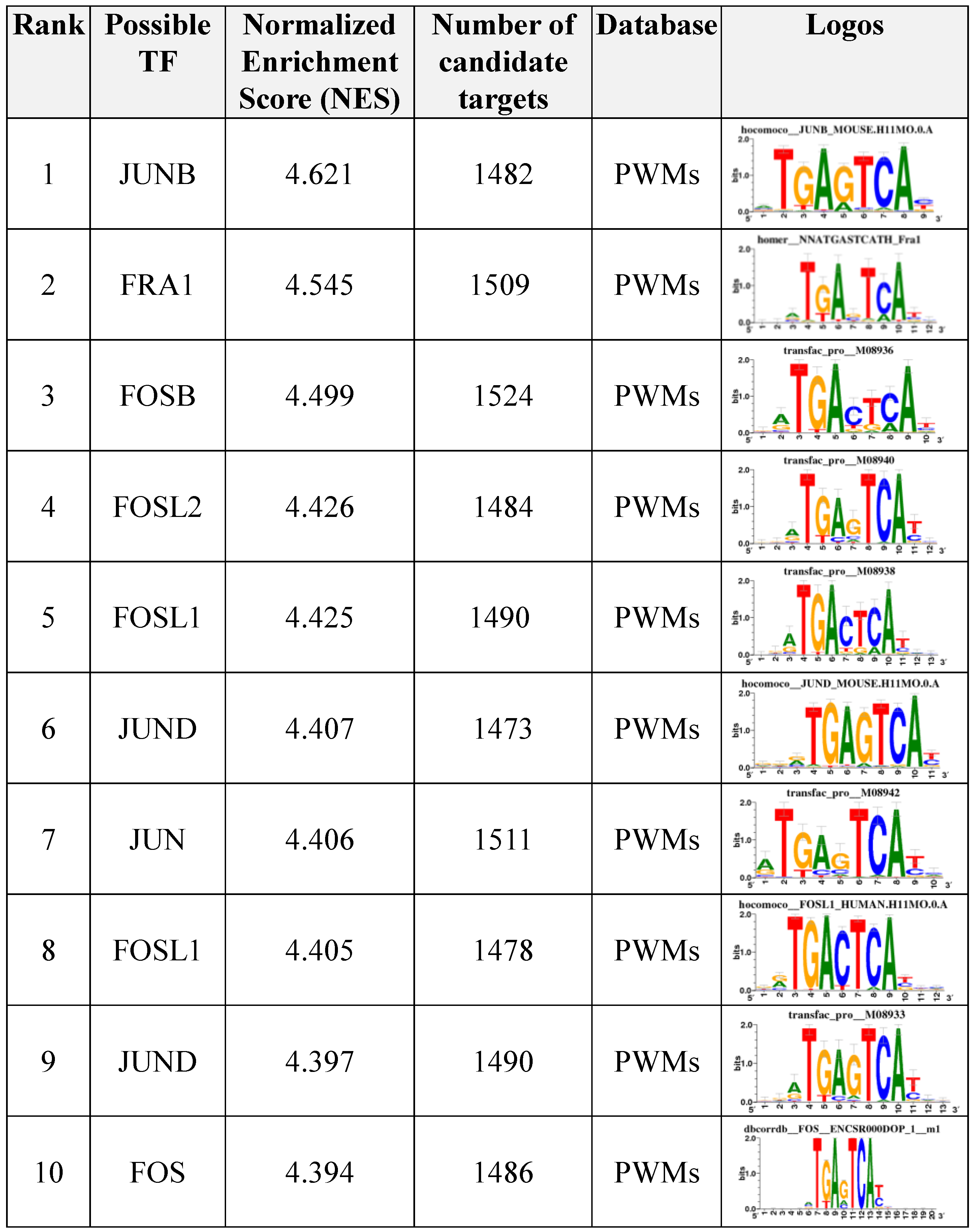
3.7. Motif Enrichment Analysis of Differentially Accessible Regions
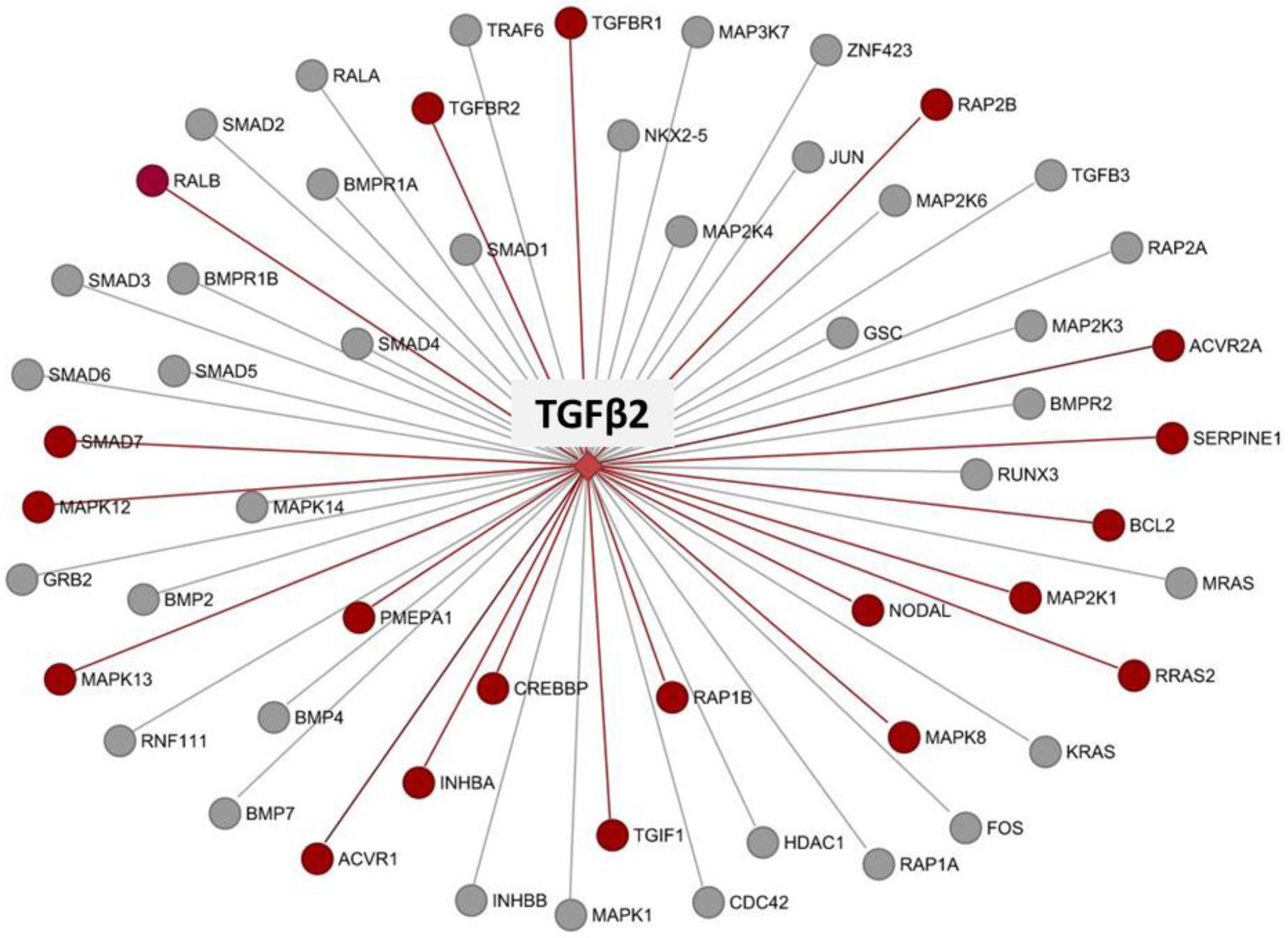
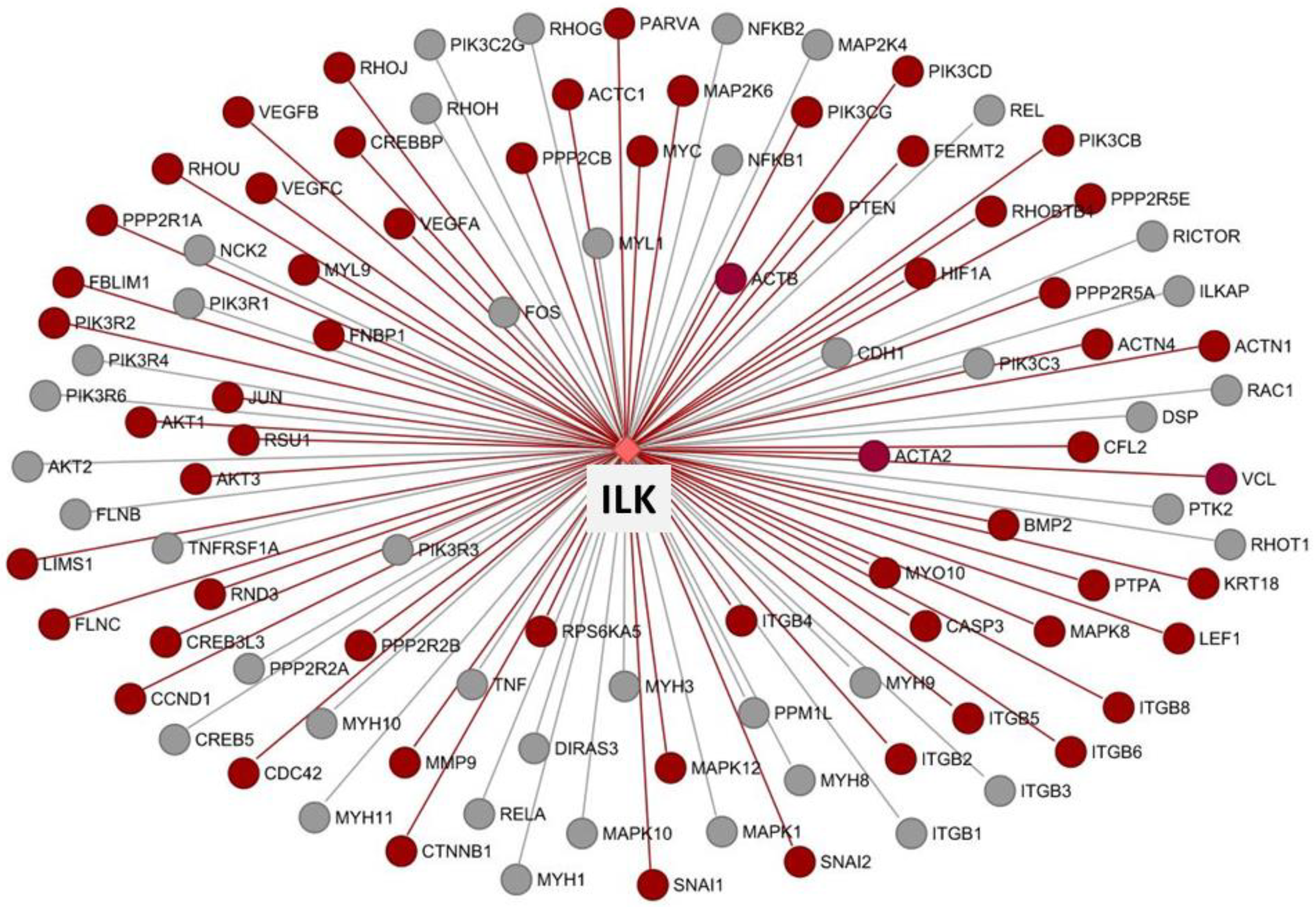
3.8. Co-Expression and Network Visualization of Critical Pathways for Rumen Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATAC-seq | assay for transposase-accessible chromatin sequencing |
| AUC | Area Under the Curve |
| BAM | binary alignment map |
| BP | biological process |
| BT | butyrate |
| CC | cellular component |
| ChIP | chromatin immunoprecipitation |
| CT | control |
| DARs | differentially accessible regions |
| DHS | DNase I hypersensitive sites |
| ENCODE | The Encyclopedia of DNA Elements |
| FAIRE | formaldehyde-assisted isolation of regulatory elements |
| FDR | false discovery rate |
| FRiP | fraction of reads in peaks |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| HDAC | histone deacetylases |
| ILK | integrin-linked kinase |
| IPA | ingenuity pathway analysis |
| MDBK | Madin–Darby bovine kidney |
| MF | molecular function |
| PWM | position weight matrices |
| REPC | rumen epithelial primary cells |
| SCFAs | short-chain fatty acids |
| TF | transcription factor |
| TOM | topological overlap matrix |
| TSS | transcription starting site |
References
- Andoh, A.; Tsujikawa, T.; Fujiyama, Y. Role of Dietary Fiber and Short-Chain Fatty Acids in the Colon. Curr. Pharm. Des. 2003, 9, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.W.; Li, C. Butyrate induces profound changes in gene expression related to multiple signal pathways in bovine kidney epithelial cells. BMC Genom. 2006, 7, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.J.; Li, R.W.; Wang, Y.H.; Elsasser, T.H. Pathway analysis identifies perturbation of genetic networks induced by butyrate in a bovine kidney epithelial cell line. Funct. Integr. Genom. 2007, 7, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Li, R.W.; Li, W.; Li, C.J. Transcriptome characterization by RNA-seq unravels the mechanisms of butyrate-induced epigenomic regulation in bovine cells. PLoS ONE 2012, 7, e36940. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Elsasser, T.H. Butyrate-induced apoptosis and cell cycle arrest in bovine kidney epithelial cells: Involvement of caspase and proteasome pathways. J. Anim. Sci. 2005, 83, 89–97. [Google Scholar] [CrossRef]
- Riggs, M.G.; Whittaker, R.G.; Neumann, J.R.; Ingram, V.M. n-Butyrate causes histone modification in HeLa and Friend erythroleukaemia cells. Nature 1977, 268, 462–464. [Google Scholar] [CrossRef]
- Li, C.-J.; Li, R.W.; Elsasser, T.H. MicroRNA (miRNA) Expression is Regulated by Butyrate-Induced Epigenetic Modulation of Gene Expression in Bovine Cells. Genet. Epigenet. 2010, 3, GEG.S6144. [Google Scholar] [CrossRef]
- Wolffe, A.P.; Guschin, D. Review: Chromatin Structural Features and Targets That Regulate Transcription. J. Struct. Biol. 2000, 129, 102–122. [Google Scholar] [CrossRef] [Green Version]
- Gorka, P.; Kowalski, Z.M.; Zabielski, R.; Guilloteau, P. Invited review: Use of butyrate to promote gastrointestinal tract development in calves. J. Dairy Sci. 2018, 101, 4785–4800. [Google Scholar] [CrossRef] [Green Version]
- Niwińska, B.; Hanczakowska, E.; Arciszewski, M.B.; Klebaniuk, R. Review: Exogenous butyrate: Implications for the functional development of ruminal epithelium and calf performance. Animal 2017, 11, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, A.D.; Allis, C.D.; Bernstein, E. Epigenetics: A Landscape Takes Shape. Cell 2007, 128, 635–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Li, R.W.; Gao, Y.; Baldwin, R.t.; Li, C.J. Genome-wide ChIP-seq mapping and analysis reveal butyrate-induced acetylation of H3K9 and H3K27 correlated with transcription activity in bovine cells. Funct. Integr. Genom. 2012, 12, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Xu, L.; Li, R.W.; Gao, Y.; Bickhart, D.; Liu, G.E.; Baldwin, R.; Li, C.J. A high-resolution whole-genome map of the distinctive epigenomic landscape induced by butyrate in bovine cells. Anim. Genet. 2014, 45 (Suppl. S1), 40–50. [Google Scholar] [CrossRef]
- Fang, L.; Liu, S.; Liu, M.; Kang, X.; Lin, S.; Li, B.; Connor, E.E.; Baldwin, R.L.t.; Tenesa, A.; Ma, L.; et al. Functional annotation of the cattle genome through systematic discovery and characterization of chromatin states and butyrate-induced variations. BMC Biol. 2019, 17, 68. [Google Scholar] [CrossRef]
- Kang, X.; Liu, S.; Fang, L.; Lin, S.; Liu, M.; Baldwin, R.L.; Liu, G.E.; Li, C.J. Data of epigenomic profiling of histone marks and CTCF binding sites in bovine rumen epithelial primary cells before and after butyrate treatment. Data Brief 2020, 28, 104983. [Google Scholar] [CrossRef]
- Lin, S.; Fang, L.; Kang, X.; Liu, S.; Liu, M.; Connor, E.E.; Baldwin, R.L.; Liu, G.; Li, C.J. Establishment and transcriptomic analyses of a cattle rumen epithelial primary cells (REPC) culture by bulk and single-cell RNA sequencing to elucidate interactions of butyrate and rumen development. Heliyon 2020, 6, e04112. [Google Scholar] [CrossRef]
- Levine, M. Transcriptional enhancers in animal development and evolution. Curr. Biol. 2010, 20, R754–R763. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.; Cattoglio, C.; Tjian, R. Looping back to leap forward: Transcription enters a new era. Cell 2014, 157, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Buecker, C.; Wysocka, J. Enhancers as information integration hubs in development: Lessons from genomics. Trends Genet. 2012, 28, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Calo, E.; Wysocka, J. Modification of enhancer chromatin: What, how, and why? Mol. Cell 2013, 49, 825–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Powell, D.R.; Curtis, D.J.; Wong, N.C. From reads to insight: A hitchhiker’s guide to ATAC-seq data analysis. Genome Biol. 2020, 21, 22. [Google Scholar] [CrossRef] [PubMed]
- Rosen, B.D.; Bickhart, D.M.; Schnabel, R.D.; Koren, S.; Elsik, C.G.; Tseng, E.; Rowan, T.N.; Low, W.Y.; Zimin, A.; Couldrey, C.; et al. De novo assembly of the cattle reference genome with single-molecule sequencing. Gigascience 2020, 9, giaa021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Ross-Innes, C.S.; Stark, R.; Teschendorff, A.E.; Holmes, K.A.; Ali, H.R.; Dunning, M.J.; Brown, G.D.; Gojis, O.; Ellis, I.O.; Green, A.R.; et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 2012, 481, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Shao, N.Y.; Liu, X.; Maze, I.; Feng, J.; Nestler, E.J. diffReps: Detecting differential chromatin modification sites from ChIP- seq data with biological replicates. PLoS ONE 2013, 8, e65598. [Google Scholar] [CrossRef] [Green Version]
- Lodato, N.J.; Rampersaud, A.; Waxman, D.J. Impact of CAR Agonist Ligand TCPOBOP on Mouse Liver Chromatin Accessibility. Toxicol. Sci. 2018, 164, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; He, Q.Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, J.; Kellis, M. ChromHMM: Automating chromatin-state discovery and characterization. Nat. Methods 2012, 9, 215–216. [Google Scholar] [CrossRef] [Green Version]
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, C.; Van de Sande, B.; Potier, D.; Aerts, S. i-cisTarget: An integrative genomics method for the prediction of regulatory features and cis-regulatory modules. Nucleic Acids Res. 2012, 40, e114. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z. Using VisANT to Analyze Networks. Curr. Protoc. Bioinform. 2014, 45, 8.8.1–8.8.39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimin, A.V.; Delcher, A.L.; Florea, L.; Kelley, D.R.; Schatz, M.C.; Puiu, D.; Hanrahan, F.; Pertea, G.; Van Tassell, C.P.; Sonstegard, T.S.; et al. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biol. 2009, 10, R42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Fang, L.; Baldwin, R.L.t.; Connor, E.E.; Cole, J.B.; Van Tassell, C.P.; Ma, L.; Li, C.J.; Liu, G.E. Single-cell transcriptomic analyses of dairy cattle ruminal epithelial cells during weaning. Genomics 2021, 113, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Radjendirane, V.; Wary, K.K.; Chakrabarty, S. Transforming growth factor beta regulates cell-cell adhesion through extracellular matrix remodeling and activation of focal adhesion kinase in human colon carcinoma Moser cells. Oncogene 2004, 23, 5558–5561. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, R.; Nguyen-Tat, M.D.; Giehl, K.; Adler, G.; Wedlich, D.; Menke, A. TGFbeta-induced downregulation of E-cadherin-based cell-cell adhesion depends on PI3-kinase and PTEN. J. Cell Sci. 2005, 118, 4901–4912. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Li, S.; Docheva, D.; Grashoff, C.; Sakai, K.; Kostka, G.; Braun, A.; Pfeifer, A.; Yurchenco, P.D.; Fässler, R. Integrin-linked kinase (ILK) is required for polarizing the epiblast, cell adhesion, and controlling actin accumulation. Genes Dev. 2003, 17, 926–940. [Google Scholar] [CrossRef] [Green Version]
- Hannigan, G.E.; Leung-Hagesteijn, C.; Fitz-Gibbon, L.; Coppolino, M.G.; Radeva, G.; Filmus, J.; Bell, J.C.; Dedhar, S. Regulation of cell adhesion and anchorage-dependent growth by a new beta 1-integrin-linked protein kinase. Nature 1996, 379, 91–96. [Google Scholar] [CrossRef]
- Bell, O.; Tiwari, V.K.; Thomä, N.H.; Schübeler, D. Determinants and dynamics of genome accessibility. Nat. Rev. Genet. 2011, 12, 554–564. [Google Scholar] [CrossRef]
- Johnston, D.; Kim, J.; Taylor, J.F.; Earley, B.; McCabe, M.S.; Lemon, K.; Duffy, C.; McMenamy, M.; Cosby, S.L.; Waters, S.M. ATAC-Seq identifies regions of open chromatin in the bronchial lymph nodes of dairy calves experimentally challenged with bovine respiratory syncytial virus. BMC Genom. 2021, 22, 14. [Google Scholar] [CrossRef]
- Alexandre, P.A.; Naval-Sanchez, M.; Menzies, M.; Nguyen, L.T.; Porto-Neto, L.R.; Fortes, M.R.S.; Reverter, A. Chromatin accessibility and regulatory vocabulary across indicine cattle tissues. Genome Biol. 2021, 22, 273. [Google Scholar] [CrossRef]
- Cao, X.; Cheng, J.; Huang, Y.; Lan, X.; Lei, C.; Chen, H. Comparative Enhancer Map of Cattle Muscle Genome Annotated by ATAC-Seq. Front. Vet. Sci. 2021, 8, 1467. [Google Scholar] [CrossRef] [PubMed]
- Halstead, M.M.; Kern, C.; Saelao, P.; Wang, Y.; Chanthavixay, G.; Medrano, J.F.; Van Eenennaam, A.L.; Korf, I.; Tuggle, C.K.; Ernst, C.W.; et al. A comparative analysis of chromatin accessibility in cattle, pig, and mouse tissues. BMC Genom. 2020, 21, 698. [Google Scholar] [CrossRef] [PubMed]
- Khalili, A.A.; Ahmad, M.R. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.C.; Qi, R.Z.; Paudel, H.; Zhu, H.J. Regulation and function of protein kinases and phosphatases. Enzyme Res. 2011, 2011, 794089. [Google Scholar] [CrossRef] [Green Version]
- Rauch, J.; Volinsky, N.; Romano, D.; Kolch, W. The secret life of kinases: Functions beyond catalysis. Cell Commun. Signal 2011, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Bond, J.J.; Donaldson, A.J.; Coumans, J.V.F.; Austin, K.; Ebert, D.; Wheeler, D.; Oddy, V.H. Protein profiles of enzymatically isolated rumen epithelium in sheep fed a fibrous diet. J. Anim. Sci. Biotechnol. 2019, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.L. The proliferative actions of insulin, insulin-like growth factor-I, epidermal growth factor, butyrate and propionate on ruminal epithelial cells in vitro. Small Rumin. Res. 1999, 32, 261–268. [Google Scholar] [CrossRef]
- LeBaron, R.G.; Bezverkov, K.I.; Zimber, M.P.; Pavelec, R.; Skonier, J.; Purchio, A.F. Beta IG-H3, a novel secretory protein inducible by transforming growth factor-beta, is present in normal skin and promotes the adhesion and spreading of dermal fibroblasts in vitro. J. Invest. Dermatol. 1995, 104, 844–849. [Google Scholar] [CrossRef] [Green Version]
- Connor, E.E.; Baldwin, R.L.t.; Walker, M.P.; Ellis, S.E.; Li, C.; Kahl, S.; Chung, H.; Li, R.W. Transcriptional regulators transforming growth factor-beta1 and estrogen-related receptor-alpha identified as putative mediators of calf rumen epithelial tissue development and function during weaning. J. Dairy Sci. 2014, 97, 4193–4207. [Google Scholar] [CrossRef]
- Brey, C.W.; Nelder, M.P.; Hailemariam, T.; Gaugler, R.; Hashmi, S. Kruppel-like family of transcription factors: An emerging new frontier in fat biology. Int. J. Biol. Sci. 2009, 5, 622–636. [Google Scholar] [CrossRef]
- Swamynathan, S.K. Kruppel-like factors: Three fingers in control. Hum. Genom. 2010, 4, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, J.M.; Christy, R.J.; Yang, V.W. Identification and characterization of a gene encoding a gut-enriched Krüppel-like factor expressed during growth arrest. J. Biol. Chem. 1996, 271, 20009–20017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaleb, A.M.; Yang, V.W. Kruppel-like factor 4 (KLF4): What we currently know. Gene 2017, 611, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Subbalakshmi, A.R.; Sahoo, S.; McMullen, I.; Saxena, A.N.; Venugopal, S.K.; Somarelli, J.A.; Jolly, M.K. KLF4 Induces Mesenchymal-Epithelial Transition (MET) by Suppressing Multiple EMT-Inducing Transcription Factors. Cancers 2021, 13, 5135. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, I.; Imoto, M.; Adjei, P.N.; Gores, G.J.; Subramaniam, M.; Spelsberg, T.C.; Urrutia, R. Overexpression of the TGFbeta-regulated zinc finger encoding gene, TIEG, induces apoptosis in pancreatic epithelial cells. J. Clin. Investig. 1997, 99, 2365–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, M.; Harris, S.A.; Oursler, M.J.; Rasmussen, K.; Riggs, B.L.; Spelsberg, T.C. Identification of a novel TGF-beta-regulated gene encoding a putative zinc finger protein in human osteoblasts. Nucleic Acids Res. 1995, 23, 4907–4912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, M.; Hawse, J.R.; Rajamannan, N.M.; Ingle, J.N.; Spelsberg, T.C. Functional role of KLF10 in multiple disease processes. Biofactors 2010, 36, 8–18. [Google Scholar] [CrossRef] [Green Version]
- Ruberto, A.A.; Grechez-Cassiau, A.; Guerin, S.; Martin, L.; Revel, J.S.; Mehiri, M.; Subramaniam, M.; Delaunay, F.; Teboul, M. KLF10 integrates circadian timing and sugar signaling to coordinate hepatic metabolism. Elife 2021, 10, e65574. [Google Scholar] [CrossRef]
- Farre, M.; Kim, J.; Proskuryakova, A.A.; Zhang, Y.; Kulemzina, A.I.; Li, Q.; Zhou, Y.; Xiong, Y.; Johnson, J.L.; Perelman, P.L.; et al. Evolution of gene regulation in ruminants differs between evolutionary breakpoint regions and homologous synteny blocks. Genome Res. 2019, 29, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Garces de Los Fayos Alonso, I.; Liang, H.C.; Turner, S.D.; Lagger, S.; Merkel, O.; Kenner, L. The Role of Activator Protein-1 (AP-1) Family Members in CD30-Positive Lymphomas. Cancers 2018, 10, 93. [Google Scholar] [CrossRef] [Green Version]
- Piechaczyk, M.; Farras, R. Regulation and function of JunB in cell proliferation. Biochem. Soc. Trans. 2008, 36, 864–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, J.M.; Floyd, D.H.; Weilbaecher, K.N.; Green, P.L.; Boris-Lawrie, K. Multiple facets of junD gene expression are atypical among AP-1 family members. Oncogene 2008, 27, 4757–4767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savy, V.; Alberio, V.; Canel, N.G.; Ratner, L.D.; Gismondi, M.I.; Ferraris, S.F.; Fernandez-Martin, R.; Knott, J.G.; Bevacqua, R.J.; Salamone, D.F. CRISPR-on for activation of endogenous SMARCA4 and TFAP2C expression in bovine embryos. Reproduction 2020, 159, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Beuling, E.; Baffour-Awuah, N.Y.; Stapleton, K.A.; Aronson, B.E.; Noah, T.K.; Shroyer, N.F.; Duncan, S.A.; Fleet, J.C.; Krasinski, S.D. GATA factors regulate proliferation, differentiation, and gene expression in small intestine of mature mice. Gastroenterology 2011, 140, 1219–1229.e1-2. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | N of Reads | N of Reads Mapped | % of Mapped Reads | N of MT Reads | % of MT Reads 1 | N of Duplicate Reads | % of Duplicate Reads 1 | N of MQ < 10 Reads | % of MQ < 10 Reads 1 | N of Clean Reads 2 |
|---|---|---|---|---|---|---|---|---|---|---|
| Butyrate 1 | 105,966,982 | 103,703,996 | 97.86 | 8,407,564 | 8.11 | 6,046,975 | 5.83 | 18,580,667 | 17.92 | 71,028,846 |
| Butyrate 2 | 103,071,942 | 100,623,739 | 97.62 | 11,467,359 | 11.40 | 5,539,727 | 5.51 | 17,727,806 | 17.62 | 66,596,923 |
| Control 1 | 54,275,704 | 54,019,636 | 99.53 | 7,069,259 | 13.09 | 1,670,430 | 3.09 | 5,993,034 | 11.09 | 38,580,543 |
| Control 2 | 110,735,198 | 108,587,882 | 98.06 | 9,386,819 | 8.64 | 8,873,305 | 8.17 | 13,695,222 | 12.61 | 77,193,679 |
| Total | 374,049,826 | 366,935,253 | - | 36,331,001 | - | 22,130,437 | - | 55,996,729 | - | 253,399,991 |
| Average | 93,512,457 | 91,733,813 | 98.27 | 9,082,750 | 10.31 | 5,532,609 | 5.65 | 13,999,182 | 14.81 | 63,349,998 |
| Condition | N of Clean Reads 1 | N of Clean Reads Used for FRiP 2 | N of MACS2 Peaks 2 | N of Assigned Reads in Peaks 2 | FRiP 3 | Average Peak Length | Proportion of Peaks near TSS (±3 kb, %) |
|---|---|---|---|---|---|---|---|
| Butyrate 1 | 71,028,846 | 69,244,296 | 118,521 | 27,738,114 | 0.39 | 389 | 12.42 |
| Butyrate 2 | 66,596,923 | 64,917,875 | 113,935 | 25,700,148 | 0.39 | 379 | 12.82 |
| Control 1 | 38,580,543 | 37,715,866 | 102,063 | 18,628,115 | 0.48 | 396 | 14.22 |
| Control 2 | 77,193,679 | 75,492,156 | 135,755 | 42,954,062 | 0.56 | 380 | 12.30 |
| Total | 253,399,991 | 247,370,193 | 470,274 | 115,020,439 | - | - | - |
| Average | 63,349,998 | 61,842,548 | 117,569 | 28,755,110 | 0.45 | 386 | 12.94 |
| Butyrate × Control DARs | N of DARs | % of DARs |
|---|---|---|
| DARs initially identified (p-value < 0.05) | 95,629 | - |
| Significant DARs (FDR < 0.01 and −1 ≤ log2FC ≥ 1) | 22,746 | 23.79 |
| Significant DARs that overlapped with peaks | 21,530 | 22.51 |
| Unique significant DARs that overlapped with peaks | 21,347 | - |
| Induced DARs with log2FC ≥ 1 | 6312 | 29.57 |
| Repressed DARs with log2FC ≤ −1 | 15,035 | 70.43 |
| Feature | Number | Frequency (%) |
|---|---|---|
| Promoter (<1 kb) | 1101 | 5.16 |
| Promoter (1–2 kb) | 428 | 2.00 |
| 5’ UTR | 12 | 0.06 |
| 3’ UTR | 118 | 0.55 |
| First Exon | 1 | 0.005 |
| Other Exon | 328 | 1.54 |
| First Intron | 1645 | 7.71 |
| Other Intron | 3357 | 15.73 |
| Downstream (<1 kb) | 47 | 0.22 |
| Downstream (1–2 kb) | 42 | 0.20 |
| Downstream (2–3 kb) | 64 | 0.30 |
| Distal Intergenic | 14,204 | 66.54 |
| Total | 21,347 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boschiero, C.; Gao, Y.; Liu, M.; Baldwin, R.L., VI; Ma, L.; Li, C.-J.; Liu, G.E. The Dynamics of Chromatin Accessibility Prompted by Butyrate-Induced Chromatin Modification in Bovine Cells. Ruminants 2022, 2, 226-243. https://doi.org/10.3390/ruminants2020015
Boschiero C, Gao Y, Liu M, Baldwin RL VI, Ma L, Li C-J, Liu GE. The Dynamics of Chromatin Accessibility Prompted by Butyrate-Induced Chromatin Modification in Bovine Cells. Ruminants. 2022; 2(2):226-243. https://doi.org/10.3390/ruminants2020015
Chicago/Turabian StyleBoschiero, Clarissa, Yahui Gao, Mei Liu, Ransom L. Baldwin, VI, Li Ma, Cong-Jun Li, and George E. Liu. 2022. "The Dynamics of Chromatin Accessibility Prompted by Butyrate-Induced Chromatin Modification in Bovine Cells" Ruminants 2, no. 2: 226-243. https://doi.org/10.3390/ruminants2020015
APA StyleBoschiero, C., Gao, Y., Liu, M., Baldwin, R. L., VI, Ma, L., Li, C.-J., & Liu, G. E. (2022). The Dynamics of Chromatin Accessibility Prompted by Butyrate-Induced Chromatin Modification in Bovine Cells. Ruminants, 2(2), 226-243. https://doi.org/10.3390/ruminants2020015