
Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes


Abstract
1. Introduction
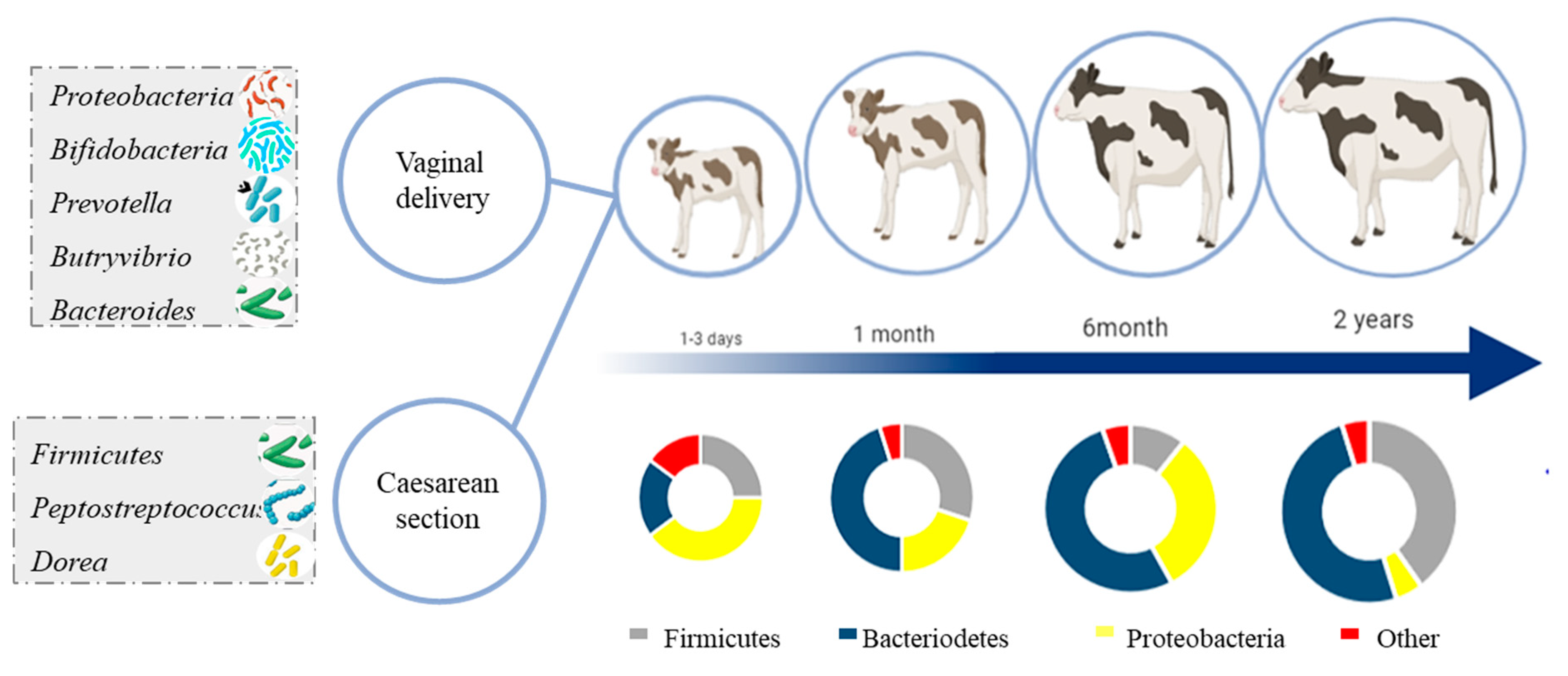
2. Development and Succession of Cattle Microbiome with Age
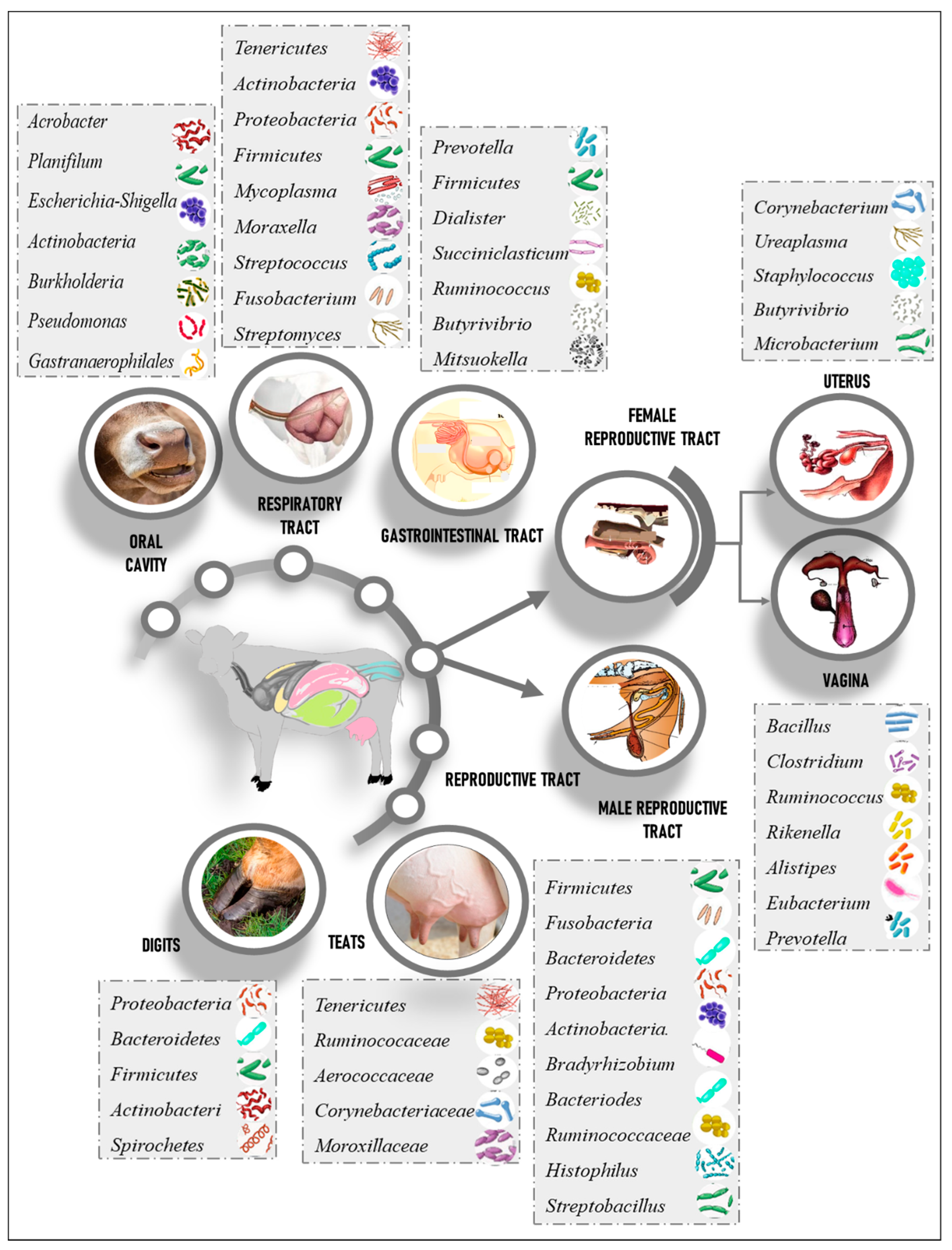
3. Healthy Cattle Microbiome Composition
3.1. Respiratory Tract Microbiome
3.2. Gastrointestinal Microbiome
3.3. Rumen Microbiome
3.4. Small Intestine
3.5. Large Intestine
3.6. Reproductive Tract Microbiome
3.7. Female Reproductive Tract
3.8. Bull Reproductive Tract
3.9. Skin Microbiome

4. Cattle Diseases Associated with Microbiome Dysbiosis
4.1. Bovine Respiratory Disease BRD
4.2. Bovine Digital Dermatitis
4.3. Mastitis
4.4. Johne’s Disease
4.5. Uterine Diseases
4.6. Metabolic Disorders
4.6.1. Ruminal Acidosis
4.6.2. Ketosis

| Disease | Significant Changes in the Microbiome | Reference |
|---|---|---|
| Bovine respiratory disease | ↑ Mycoplama bovis ↑ Mannheimia haemolytica ↑ Pasteurella multocida ↑ Mycoplasma | [67,68] |
| Mastitis | ↑ Staphylococcus aureus ↑ Streptococcus agalactiae ↑ Escherichia coli ↑ Klebsiella ↑ Streptococcus dysgalactia, ↑ Corynebacterium bovis | [85,86,88] |
| Johnes Disease | ↑ Arthrobacter ↑ Bacillus ↑ Enterococcus ↑ Camobacterium ↑ Desemzia ↑ Trichococcus ↑ Planococcaceae ↓ Paraprevotellaceae ↓ Faecalibacterium ↓ Akkermansia | [19,105] |
| Metritis and endometritis | ↑ Trueperella pyogenes ↑ Escherichia coli ↑ Porphyromonas levii ↑ Fusobacterium necrophorum ↑ Prevotella melaninogenica ↑ Bacteroides spp | [20,111,114,116,117] |
| Bovine digital dermatitis | ↑ Treponema spp. ↑ Fusobacterium spp. ↑ Mycoplasma spp. ↑ Porphyromonas spp. ↑ Prevotella spp. ↑ Corynebacterium spp. ↑ Tissierella spp. | [59] |
| Rumen Acidosis | ↓ Bacteroidetes ↓ Fibrobacter succinogenes ↓ Ruminococcus albus ↓ Ruminococcus bicirculans ↓ Butyrivibrio fibrisolvens ↑ Prevotella bryantii ↑ Selenomonas ruminantium ↓ Streptococcus ↑ Lactobacillus ↑ Succiniclasticum ↑ Clostridium | [31,127,128] |
| Ketosis | ↑ Lachnosparaceae ↑ A. lipolytica, ↑ P. bryantii ↑ M. elsdenii ↓ Ruminococcaceae ↓ Methanobrevibacter ↓ Erysipelotrichaceae ↓ Atopobium ↓ F. succinogenes ↓ Butyrivibrio proteoclasticus ↓ Euryarchaeota | [27,133] |
5. Non-Antibiotics Microbial Therapy
5.1. Probiotics
5.2. Phage Therapy
5.3. Prebiotics
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Hara, E.; Neves, A.L.; Song, Y.; Guan, L.L. The Role of the Gut Microbiome in Cattle Production and Health: Driver or Passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef] [PubMed]
- Cammack, K.M.; Austin, K.J.; Lamberson, W.R.; Conant, G.C.; Cunningham, H.C. RUMINNAT NUTRITION SYMPOSIUM: Tiny but mighty: The role of the rumen microbes in livestock production. J. Anim. Sci. 2018, 96, 752–770. [Google Scholar] [CrossRef] [PubMed]
- Firkins, J.; Yu, Z. Ruminant nutrition symposium: How to use data on the rumen microbiome to improve our understanding of ruminant nutrition. J. Anim. Sci. 2015, 93, 1450–1470. [Google Scholar] [CrossRef] [PubMed]
- Bergman, E. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.; Calsamiglia, S.; Stern, M. Nitrogen metabolism in the rumen. J. Dairy Sci. 2005, 88, E9–E21. [Google Scholar] [CrossRef]
- Van den Broek, M.F.; De Boeck, I.; Kiekens, F.; Boudewyns, A.; Vanderveken, O.M.; Lebeer, S. Translating recent microbiome insights in otitis media into probiotic strategies. Clin. Microbiol. Rev. 2019, 32, e00010–e00018. [Google Scholar] [CrossRef]
- Karstrup, C.C.; Klitgaard, K.; Jensen, T.K.; Agerholm, J.S.; Pedersen, H.G. Presence of bacteria in the endometrium and placentomes of pregnant cows. Theriogenology 2017, 99, 41–47. [Google Scholar] [CrossRef]
- Moore, S.; Ericsson, A.; Poock, S.; Melendez, P.; Lucy, M. Hot topic: 16S rRNA gene sequencing reveals the microbiome of the virgin and pregnant bovine uterus. J. Dairy Sci. 2017, 100, 4953–4960. [Google Scholar] [CrossRef]
- Krishnan, D.; Al-Harbi, H.; Gibson, J.; Olchowy, T.; Alawneh, J. On the use of probiotics to improve dairy cattle health and productivity. Microbiol. Aust. 2020, 41, 86–90. [Google Scholar] [CrossRef]
- Laguardia-Nascimento, M.; Branco, K.M.G.R.; Gasparini, M.R.; Giannattasio-Ferraz, S.; Leite, L.R.; Araujo, F.M.G.; Salim, A.C.d.M.; Nicoli, J.R.; De Oliveira, G.C.; Barbosa-Stancioli, E.F. Vaginal microbiome characterization of Nellore cattle using metagenomic analysis. PLoS ONE 2015, 10, e0143294. [Google Scholar] [CrossRef]
- Alexander, T. 145 The role of the bovine respiratory microbiota in health and disease. J. Anim. Sci. 2018, 96, 341. [Google Scholar] [CrossRef]
- Andersen, T.O. Understanding Rumen Function Using (Meta) Genome-Guided Metaproteomics. Master’s Thesis, Norwegian University of Life Sciences, Ås, Norway, 2019. [Google Scholar]
- Clemmons, B.A.; Reese, S.T.; Dantas, F.G.; Franco, G.A.; Smith, T.P.; Adeyosoye, O.I.; Pohler, K.G.; Myer, P.R. Vaginal and uterine bacterial communities in postpartum lactating cows. Front. Microbiol. 2017, 8, 1047. [Google Scholar] [CrossRef] [PubMed]
- Manus, M.B.; Yu, J.J.; Park, L.P.; Mueller, O.; Windsor, S.C.; Horvath, J.E.; Nunn, C.L. Environmental influences on the skin microbiome of humans and cattle in rural Madagascar. Evol. Med. Public Health 2017, 1, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome community structure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The evolution of the host microbiome as an ecosystem on a leash. Nature 2017, 548, 43–51. [Google Scholar] [CrossRef]
- Furman, O.; Shenhav, L.; Sasson, G.; Kokou, F.; Honig, H.; Jacoby, S.; Hertz, T.; Cordero, O.X.; Halperin, E.; Mizrahi, I. Stochasticity constrained by deterministic effects of diet and age drive rumen microbiome assembly dynamics. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Wilkins, L.J.; Monga, M.; Miller, A.W. Defining dysbiosis for a cluster of chronic diseases. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Derakhshani, H.; De Buck, J.; Mortier, R.; Barkema, H.W.; Krause, D.O.; Khafipour, E. The features of fecal and ileal mucosa-associated microbiota in dairy calves during early infection with Mycobacterium avium subspecies paratuberculosis. Front. Microbiol. 2016, 7, 426. [Google Scholar] [CrossRef]
- Jeon, S.J.; Cunha, F.; Ma, X.; Martinez, N.; Vieira-Neto, A.; Daetz, R.; Bicalho, R.C.; Lima, S.; Santos, J.E.; Jeong, K.C. Uterine microbiota and immune parameters associated with fever in dairy cows with metritis. PLoS ONE 2016, 11, e0165740. [Google Scholar] [CrossRef]
- Balato, A.; Cacciapuoti, S.; Di Caprio, R.; Marasca, C.; Masarà, A.; Raimondo, A.; Fabbrocini, G. Human microbiome: Composition and role in inflammatory skin diseases. Arch. Immunol. Et Ther. Exp. 2019, 67, 1–18. [Google Scholar] [CrossRef]
- Economou, V.; Gousia, P. Agriculture and food animals as a source of antimicrobial-resistant bacteria. Infect. Drug Resist. 2015, 8, 49. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, K.; Zhang, C.; Feng, Y.; Zhang, X.; Wang, X.; Wu, G. Dynamics and stabilization of the rumen microbiome in yearling Tibetan sheep. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Clemmons, B.A.; Voy, B.H.; Myer, P.R. Altering the gut microbiome of cattle: Considerations of host-microbiome interactions for persistent microbiome manipulation. Microb. Ecol. 2019, 77, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Cholewińska, P.; Czyż, K.; Nowakowski, P.; Wyrostek, A. The microbiome of the digestive system of ruminants–A review. Anim. Health Res. Rev. 2020, 21, 3–14. [Google Scholar] [CrossRef]
- Greenhalgh, K.; Meyer, K.M.; Aagaard, K.M.; Wilmes, P. The human gut microbiome in health: Establishment and resilience of microbiota over a lifetime. Environ. Microbiol. 2016, 18, 2103–2116. [Google Scholar] [CrossRef]
- Cunningham, H.C.; Austin, K.J.; Cammack, K.M. Influence of maternal factors on the rumen microbiome and subsequent host performance. Transl. Anim. Sci. 2018, 2, S101–S105. [Google Scholar] [CrossRef]
- Govil, K.; Yadav, D.; Patil, A.; Nayak, S.; Baghel, R.; Yadav, P.; Malapure, C.; Thakur, D. Feeding management for early rumen development in calves. J. Entomol. Zool. Stud. 2017, 5, 1132–1139. [Google Scholar]
- McCann, J.C.; Wickersham, T.A.; Loor, J.J. High-throughput methods redefine the rumen microbiome and its relationship with nutrition and metabolism. Bioinform. Biol. Insights 2014, 8, BBI-S15389. [Google Scholar] [CrossRef]
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Ruiz, D.R.; Abecia, L.; Newbold, C.J. Manipulating rumen microbiome and fermentation through interventions during early life: A review. Front. Microbiol. 2015, 6, 1133. [Google Scholar] [CrossRef] [PubMed]
- Rey, M.; Enjalbert, F.; Monteils, V. Establishment of ruminal enzyme activities and fermentation capacity in dairy calves from birth through weaning. J. Dairy Sci. 2012, 95, 1500–1512. [Google Scholar] [CrossRef] [PubMed]
- McMullen, C.; Alexander, T.W.; Léguillette, R.; Workentine, M.; Timsit, E. Topography of the respiratory tract bacterial microbiota in cattle. Microbiome 2020, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Timsit, E.; McMullen, C.; Amat, S.; Alexander, T.W. Respiratory bacterial microbiota in cattle: From development to modulation to enhance respiratory health. Veterinary Clinics. Food Anim. Pract. 2020, 36, 297–320. [Google Scholar] [CrossRef]
- Borsanelli, A.; Ramos, T.; Gaetti-Jardim, E.; Schweitzer, C.; Dutra, I. Treponema species in the subgingival microflora of ovine periodontitis. Vet. Rec. 2017, 180, 150. [Google Scholar] [CrossRef] [PubMed]
- Holcombe, L.J.; Patel, N.; Colyer, A.; Deusch, O.; O’Flynn, C.; Harris, S. Early canine plaque biofilms: Characterization of key bacterial interactions involved in initial colonization of enamel. PLoS ONE 2014, 9, e113744. [Google Scholar] [CrossRef]
- Sturgeon, A.; Pinder, S.; Costa, M.; Weese, J. Characterization of the oral microbiota of healthy cats using next-generation sequencing. Vet. J. 2014, 201, 223–229. [Google Scholar] [CrossRef]
- Borsanelli, A.C.; Lappin, D.F.; Viora, L.; Bennett, D.; Dutra, I.S.; Brandt, B.W.; Riggio, M.P. Microbiomes associated with bovine periodontitis and oral health. Vet. Microbiol. 2018, 218, 1–6. [Google Scholar] [CrossRef]
- Membrive, C.M.B. Anatomy and Physiology of the Rumen. In Rumenology; Millen, D.D., Arrigoni, M.B., Pacheco, R.D.L., Eds.; Springer: Basel, Switzerland, 2016; pp. 1–38. [Google Scholar]
- Petri, R.M.; Neubauer, V.; Humer, E.; Kröger, I.; Reisinger, N.; Zebeli, Q. Feed additives differentially impact the epimural microbiota and host epithelial gene expression of the bovine rumen fed diets rich in concentrates. Front. Microbiol. 2020, 11, 119. [Google Scholar] [CrossRef]
- Myer, P.; Freetly, H.; Wells, J.; Smith, T.; Kuehn, L. Analysis of the gut bacterial communities in beef cattle and their association with feed intake, growth, and efficiency. J. Anim. Sci. 2017, 95, 3215–3224. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Zhang, M.; Liu, J.; Zhu, W. Characterising the bacterial microbiota across the gastrointestinal tracts of dairy cattle: Membership and potential function. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Harfoot, C. Anatomy, physiology and microbiology of the ruminant digestive tract. In Lipid Metabolism in Ruminant Animals; Christie , W.W., Ed.; Pergamon Press: Oxford, UK, 1981; pp. 1–19. [Google Scholar]
- Myer, P.R.; Wells, J.E.; Smith, T.P.; Kuehn, L.A.; Freetly, H.C. Microbial community profiles of the colon from steers differing in feed efficiency. Springerplus 2015, 4, 454. [Google Scholar] [CrossRef]
- Durso, L.M.; Miller, D.N.; Schmidt, T.B.; Callaway, T. Tracking bacteria through the entire gastrointestinal tract of a beef steer. Agric. Environ. Lett. 2017, 2, 1–5. [Google Scholar] [CrossRef]
- Schmidt, C. Out of your skin. Nat. Biotechnol. 2020, 38, 392–397. Available online: https://www.nature.com/articles/s41587-020-0473-8 (accessed on 5 June 2020). [CrossRef] [PubMed]
- McClure, M.W. The Vaginal Microbiome Related to Reproductive Traits in Beef Heifers. Master’s Thesis, University of Arkansas, Fayetteville, NC, USA, 2018. [Google Scholar]
- Nesengani, L.T.; Wang, J.; Yang, Y.; Yang, L.; Lu, W. Unravelling vaginal microbial genetic diversity and abundance between Holstein and Fleckvieh cattle. RSC Adv. 2017, 7, 56137–56143. [Google Scholar] [CrossRef]
- Giannattasio-Ferraz, S.; Laguardia-Nascimento, M.; Gasparini, M.R.; Leite, L.R.; Araujo, F.M.G.; de Matos Salim, A.C.; de Oliveira, A.P.; Nicoli, J.R.; de Oliveira, G.C.; da Fonseca, F.G. A common vaginal microbiota composition among breeds of Bos taurus indicus (Gyr and Nellore). Braz. J. Microbiol. 2019, 50, 1115–1124. [Google Scholar] [CrossRef]
- Ewies, A.A.; Khan, Z.R. Cattle uterus: A novel animal laboratory model for advanced hysteroscopic surgery training. Obstet. Gynecol. Int. 2015, 2015, 967693. [Google Scholar] [CrossRef]
- Wolfe, D. Abnormalities of the bull–occurrence, diagnosis and treatment of abnormalities of the bull, including structural soundness. Animal 2018, 12, s148–s157. [Google Scholar] [CrossRef]
- Comizzoli, P.; Power, M. Reproductive microbiomes in wild animal species: A new dimension in conservation biology. In Reproductive Sciences in Animal Conservation; Comizzoli, P., Brown, J., Holt, W., Eds.; Springer: Cham, Switzerland, 2019; pp. 225–240. [Google Scholar]
- Wickware, C.L.; Johnson, T.A.; Koziol, J.H. Composition and diversity of the preputial microbiota in healthy bulls. Theriogenology 2020, 145, 231–237. [Google Scholar] [CrossRef]
- Frétin, M.; Martin, B.; Rifa, E.; Isabelle, V.-M.; Pomiès, D.; Ferlay, A.; Montel, M.-C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Verdier-Metz, I.; Gagne, G.; Bornes, S.; Monsallier, F.; Veisseire, P.; Delbès-Paus, C.; Montel, M.-C. Cow teat skin, a potential source of diverse microbial populations for cheese production. Appl. Environ. Microbiol. 2012, 78, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.R. The cutaneous ecosystem: The roles of the skin microbiome in health and its association with inflammatory skin conditions in humans and animals. Adv. Vet. Dermatol. 2017, 8, 71–83. [Google Scholar]
- Ariza, J.M.; Dopfer, D.; Anklam, K.; Labrut, S.; Oberle, K.; Bareille, N.; Relun, A.; Guatteo, R. Do footbath disinfectants modify the dynamics of the skin microbiota in dairy cattle affected by digital dermatitis? bioRxiv. 2019. Available online: https://www.biorxiv.org/content/10.1101/2019.12.20.882787v1.abstract (accessed on 1 May 2020).
- Krull, A.C.; Shearer, J.K.; Gorden, P.J.; Cooper, V.L.; Phillips, G.J.; Plummer, P.J. Deep sequencing analysis reveals temporal microbiota changes associated with development of bovine digital dermatitis. Infect. Immun. 2014, 82, 3359–3373. [Google Scholar] [CrossRef] [PubMed]
- Mamuad, L.L.; Seo, B.J.; Al Faruk, M.S.; Espiritu, H.M.; Jin, S.J.; Kim, W.-I.; Lee, S.-S.; Cho, Y.-I. Treponema spp., the dominant pathogen in the lesion of bovine digital dermatitis and its characterization in dairy cattle. Vet. Microbiol. 2020, 245, 108696. [Google Scholar] [CrossRef]
- Holman, D.B.; Gzyl, K.E. A meta-analysis of the bovine gastrointestinal tract microbiota. FEMS Microbiol. Ecol. 2019, 95, fiz072. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.H.; Agnol, A.M.D.; Fritzen, J.T.; Lorenzetti, E.; Alfieri, A.A.; Alfieri, A.F. Microbial diversity involved in the etiology of a bovine respiratory disease outbreak in a dairy calf rearing unit. Comp. Immunol. Microbiol. Infect. Dis. 2020, 71, 101494. [Google Scholar] [CrossRef]
- Pansri, P.; Katholm, J.; Krogh, K.; Aagaard, A.; Schmidt, L.; Kudirkiene, E.; Larsen, L.E.; Olsen, J. Evaluation of novel multiplex qPCR assays for diagnosis of pathogens associated with the bovine respiratory disease complex. Vet. J. 2020, 256, 105425. [Google Scholar] [CrossRef]
- Johnson, K.K.; Pendell, D.L. Market impacts of reducing the prevalence of bovine respiratory disease in United States beef cattle feedlots. Front. Vet. Sci. 2017, 4, 189. [Google Scholar] [CrossRef]
- Lima, S.F.; Teixeira, A.G.V.; Higgins, C.H.; Lima, F.S.; Bicalho, R.C. The upper respiratory tract microbiome and its potential role in bovine respiratory disease and otitis media. Sci. Rep. 2016, 6, 29050. [Google Scholar] [CrossRef]
- Gaeta, N.C.; Lima, S.F.; Teixeira, A.G.; Ganda, E.K.; Oikonomou, G.; Gregory, L.; Bicalho, R.C. Deciphering upper respiratory tract microbiota complexity in healthy calves and calves that develop respiratory disease using shotgun metagenomics. J. Dairy Sci. 2017, 100, 1445–1458. [Google Scholar] [CrossRef]
- Timsit, E.; Workentine, M.; van der Meer, F.; Alexander, T. Distinct bacterial metacommunities inhabit the upper and lower respiratory tracts of healthy feedlot cattle and those diagnosed with bronchopneumonia. Vet. Microbiol. 2018, 221, 105–113. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, B.A.F.D.; Gaeta, N.C.; Ribeiro, B.L.M.; Alemán, M.A.R.; Marques, L.M.; Timenetsky, J.; Melville, P.A.; Marques, J.A.; Marvulle, V.; Gregory, L. Determination of bacterial aetiologic factor on tracheobronchial lavage in relation to clinical signs of bovine respiratory disease. J. Med. Microbiol. 2016, 65, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Staton, G.; Carter, S.; Ainsworth, S.; Mullin, J.; Smith, R.; Evans, N. Putative β-barrel outer membrane proteins of the bovine digital dermatitis-associated treponemes: Identification, functional characterization, and immunogenicity. Infect. Immun. 2020, 88, e00050-20. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.A.; O’Connell, N.E. Digital dermatitis in dairy cows: A review of risk factors and potential sources of between-animal variation in susceptibility. Animals 2015, 5, 512–535. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, A.; Evans, N. Infection reservoirs and transmission of digital dermatitis in the dairy herd. Livestock 2020, 25, 118–124. [Google Scholar] [CrossRef]
- Nielsen, M.W.; Strube, M.L.; Isbrand, A.; Al-Medrasi, W.D.; Boye, M.; Jensen, T.K.; Klitgaard, K. Potential bacterial core species associated with digital dermatitis in cattle herds identified by molecular profiling of interdigital skin samples. Vet. Microbiol. 2016, 186, 139–149. [Google Scholar] [CrossRef]
- Zinicola, M.; Lima, F.; Lima, S.; Machado, V.; Gomez, M.; Döpfer, D.; Guard, C.; Bicalho, R. Altered microbiomes in bovine digital dermatitis lesions, and the gut as a pathogen reservoir. PLoS ONE 2015, 10, 1–10. [Google Scholar] [CrossRef]
- Klitgaard, K.; Bretó, A.F.; Boye, M.; Jensen, T.K. Targeting the treponemal microbiome of digital dermatitis infections by high-resolution phylogenetic analyses and comparison with fluorescent in situ hybridization. J. Clin. Microbiol. 2013, 51, 2212–2219. [Google Scholar] [CrossRef]
- Krull, A.C.; Cooper, V.L.; Coatney, J.W.; Shearer, J.K.; Gorden, P.J.; Plummer, P.J. A highly effective protocol for the rapid and consistent induction of digital dermatitis in Holstein calves. PLoS ONE 2016, 11, 1–16. [Google Scholar] [CrossRef]
- Vermeersch, A.-S.; Opsomer, G. Digital dermatitis in cattle Part I: Factors contributing to the development of digital dermatitis. Vlaams Diergeneeskd. Tijdschr. 2019, 88, 247–257. [Google Scholar] [CrossRef]
- Knappe-Poindecker, M.; Gilhuus, M.; Jensen, T.K.; Klitgaard, K.; Larssen, R.; Fjeldaas, T. Interdigital dermatitis, heel horn erosion, and digital dermatitis in 14 Norwegian dairy herds. J. Dairy Sci. 2013, 96, 7617–7629. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.; Capion, N.; Klitgaard, K.; Rogdo, T.; Fjeldaas, T.; Boye, M.; Jensen, T.K. Bovine digital dermatitis: Possible pathogenic consortium consisting of Dichelobacter nodosus and multiple Treponema species. Vet. Microbiol. 2012, 160, 151–161. [Google Scholar] [CrossRef]
- Alsaaod, M.; Locher, I.; Jores, J.; Grimm, P.; Brodard, I.; Steiner, A.; Kuhnert, P. Detection of specific Treponema species and Dichelobacter nodosus from digital dermatitis (Mortellaro’s disease) lesions in Swiss cattle. Schweiz. Arch. Tierheilkd. 2019, 161, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, N.M.; Mahmmod, Y.S.; Refaai, W.; Awad, A. Molecular detection of Treponema species organisms in foremilk and udder cleft skin of dairy cows with digital dermatitis. Trop. Anim. Health Prod. 2020, 52, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Abebe, R.; Hatiya, H.; Abera, M.; Megersa, B.; Asmare, K. Bovine mastitis: Prevalence, risk factors and isolation of Staphylococcus aureus in dairy herds at Hawassa milk shed, South Ethiopia. BMC Vet. Res. 2016, 12, 270. [Google Scholar] [CrossRef]
- Hoque, M.N.; Istiaq, A.; Clement, R.A.; Gibson, K.M.; Saha, O.; Islam, O.K.; Abir, R.A.; Sultana, M.; Siddiki, A.; Crandall, K.A. Insights Into the Resistome of Bovine Clinical Mastitis Microbiome, a Key Factor in Disease Complication. Front. Microbiol. 2020, 11, 860. [Google Scholar] [CrossRef]
- Ashraf, A.; Imran, M. Diagnosis of bovine mastitis: From laboratory to farm. Trop. Anim. Health Prod. 2018, 50, 1193–1202. [Google Scholar] [CrossRef]
- Dalanezi, F.; Joaquim, S.; Guimarães, F.; Guerra, S.; Lopes, B.; Schmidt, E.; Cerri, R.; Langoni, H. Influence of pathogens causing clinical mastitis on reproductive variables of dairy cows. J. Dairy Sci. 2020, 103, 3648–3655. [Google Scholar] [CrossRef]
- Vakkamäki, J.; Taponen, S.; Heikkilä, A.-M.; Pyörälä, S. Bacteriological etiology and treatment of mastitis in Finnish dairy herds. Acta Vet. Scand. 2017, 59, 1–9. [Google Scholar] [CrossRef]
- Hoque, M.N.; Istiaq, A.; Clement, R.A.; Sultana, M.; Crandall, K.A.; Siddiki, A.Z.; Hossain, M.A. Association of milk microbiome in bovine clinical mastitis and their functional implications in cows in Bangladesh. bioRxiv 2019, 591982. [Google Scholar] [CrossRef]
- Hoque, M.N.; Istiaq, A.; Clement, R.A.; Sultana, M.; Crandall, K.A.; Siddiki, A.Z.; Hossain, M.A. Metagenomic deep sequencing reveals association of microbiome signature with functional biases in bovine mastitis. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P. Mammary microbiota of dairy ruminants: Fact or fiction? Vet. Res. 2017, 48, 25. [Google Scholar] [CrossRef] [PubMed]
- Taponen, S.; McGuinness, D.; Hiitiö, H.; Simojoki, H.; Zadoks, R.; Pyörälä, S. Bovine milk microbiome: A more complex issue than expected. Vet. Res. 2019, 50, 44. [Google Scholar] [CrossRef]
- Ganda, E.K.; Bisinotto, R.S.; Lima, S.F.; Kronauer, K.; Decter, D.H.; Oikonomou, G.; Schukken, Y.H.; Bicalho, R.C. Longitudinal metagenomic profiling of bovine milk to assess the impact of intramammary treatment using a third-generation cephalosporin. Sci. Rep. 2016, 6, 37565. [Google Scholar] [CrossRef]
- Derakhshani, H.; Fehr, K.B.; Sepehri, S.; Francoz, D.; De Buck, J.; Barkema, H.W.; Plaizier, J.C.; Khafipour, E. Invited review: Microbiota of the bovine udder: Contributing factors and potential implications for udder health and mastitis susceptibility. J. Dairy Sci. 2018, 101, 10605–10625. [Google Scholar] [CrossRef]
- Derakhshani, H.; Plaizier, J.C.; De Buck, J.; Barkema, H.W.; Khafipour, E. Composition and co-occurrence patterns of the microbiota of different niches of the bovine mammary gland: Potential associations with mastitis susceptibility, udder inflammation, and teat-end hyperkeratosis. Anim. Microbiome 2020, 2, 1–17. [Google Scholar] [CrossRef]
- Hu, X.; Guo, J.; Mu, R.; Jiang, P.; Zhao, C.; Cao, Y.; Zhang, N.; Fu, Y. The Correlation and Mechanism between Rumen Microbiota Disturbance and Mastitis in Dairy Cows. 2020. Available online: https://www.researchsquare.com/article/rs-32289/v1 (accessed on 1 May 2020).
- Ma, C.; Sun, Z.; Zeng, B.; Huang, S.; Zhao, J.; Zhang, Y.; Su, X.; Xu, J.; Wei, H.; Zhang, H. Cow-to-mouse fecal transplantations suggest intestinal microbiome as one cause of mastitis. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef]
- Hu, X.; Guo, J.; Zhao, C.; Jiang, P.; Maimai, T.; Yanyi, L.; Cao, Y.; Fu, Y.; Zhang, N. The gut microbiota contributes to the development of Staphylococcus aureus-induced mastitis in mice. ISME J. 2020, 14, 1–14. [Google Scholar] [CrossRef]
- Hu, X.; Li, S.; Fu, Y.; Zhang, N. Targeting gut microbiota as a possible therapy for mastitis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1409–1423. [Google Scholar] [CrossRef]
- Rathnaiah, G.; Zinniel, D.K.; Bannantine, J.P.; Stabel, J.R.; Gröhn, Y.T.; Collins, M.T.; Barletta, R.G. Pathogenesis, molecular genetics, and genomics of Mycobacterium avium subsp. paratuberculosis, the etiologic agent of Johne’s disease. Front. Vet. Sci. 2017, 4, 187. [Google Scholar] [CrossRef] [PubMed]
- Shoyama, F.M.; Janetanakit, T.; Bannantine, J.P.; Barletta, R.G.; Sreevatsan, S. Elucidating the Regulon of a Fur-like Protein in Mycobacterium avium subsp. paratuberculosis (MAP). Front. Microbiol. 2020, 11, 598. [Google Scholar] [CrossRef]
- Britton, L.E.; Cassidy, J.P.; O’Donovan, J.; Gordon, S.V.; Markey, B. Potential application of emerging diagnostic techniques to the diagnosis of bovine Johne’s disease (paratuberculosis). Vet. J. 2016, 209, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Eslami, M.; Shafiei, M.; Ghasemian, A.; Valizadeh, S.; Al-Marzoqi, A.H.; Shokouhi Mostafavi, S.K.; Nojoomi, F.; Mirforughi, S.A. Mycobacterium avium paratuberculosis and Mycobacterium avium complex and related subspecies as causative agents of zoonotic and occupational diseases. J. Cell. Physiol. 2019, 234, 12415–12421. [Google Scholar] [CrossRef] [PubMed]
- Pierce, E.S. Could Mycobacterium avium subspecies paratuberculosis cause Crohn’s disease, ulcerative colitis… and colorectal cancer? Infect. Agents Cancer 2018, 13, 1–6. [Google Scholar] [CrossRef]
- Torres, J.; Mehandru, S.; Colombel, J.-F.; Peyrin-Biroulet, L. Crohn’s disease. Lancet 2017, 389, 1741–1755. [Google Scholar] [CrossRef]
- Timms, V.J.; Daskalopoulos, G.; Mitchell, H.M.; Neilan, B.A. The association of Mycobacterium avium subsp. paratuberculosis with inflammatory bowel disease. PLoS ONE 2016, 11, e0148731. [Google Scholar] [CrossRef]
- Fecteau, M.-E.; Pitta, D.W.; Vecchiarelli, B.; Indugu, N.; Kumar, S.; Gallagher, S.C.; Fyock, T.L.; Sweeney, R.W. Dysbiosis of the fecal microbiota in cattle infected with Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2016, 11, e0160353. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Cronin, J.G.; Bromfield, J.J. Tolerance and innate immunity shape the development of postpartum uterine disease and the impact of endometritis in dairy cattle. Annu. Rev. Anim. Biosci. 2019, 7, 361–384. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Owens, S.E. Postpartum uterine infection and endometritis in dairy cattle. Anim. Reprod. 2018, 14, 622–629. [Google Scholar] [CrossRef]
- Mohammed, Z.; Mann, G.; Robinson, R. Impact of endometritis on post-partum ovarian cyclicity in dairy cows. Vet. J. 2019, 248, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.; Bicalho, R. The infectious disease epidemiologic triangle of bovine uterine diseases. Anim. Reprod. 2018, 12, 450–464. [Google Scholar]
- Knudsen, L.R.V.; Karstrup, C.C.; Pedersen, H.G.; Angen, Ø.; Agerholm, J.S.; Rasmussen, E.L.; Jensen, T.K.; Klitgaard, K. An investigation of the microbiota in uterine flush samples and endometrial biopsies from dairy cows during the first 7 weeks postpartum. Theriogenology 2016, 86, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.; Cunha, F.; Daetz, R.; Figueiredo, C.; Chebel, R.; Santos, J.; Risco, C.; Jeong, K.; Machado, V.; Galvão, K. Using chitosan microparticles to treat metritis in lactating dairy cows. J. Dairy Sci. 2020, 103, 7377–7391. [Google Scholar] [CrossRef]
- Adnane, M.; Kaidi, R.; Hanzen, C.; England, G.C. Risk factors of clinical and subclinical endometritis in cattle: A review. Turk. J. Vet. Anim. Sci. 2017, 41, 1–11. [Google Scholar] [CrossRef]
- Bicalho, M.; Santin, T.; Rodrigues, M.; Marques, C.; Lima, S.; Bicalho, R. Dynamics of the microbiota found in the vaginas of dairy cows during the transition period: Associations with uterine diseases and reproductive outcome. J. Dairy Sci. 2017, 100, 3043–3058. [Google Scholar] [CrossRef]
- Jeon, S.J.; Vieira-Neto, A.; Gobikrushanth, M.; Daetz, R.; Mingoti, R.D.; Parize, A.C.B.; de Freitas, S.L.; da Costa, A.N.L.; Bicalho, R.C.; Lima, S. Uterine microbiota progression from calving until establishment of metritis in dairy cows. Appl. Environ. Microbiol. 2015, 81, 6324–6332. [Google Scholar] [CrossRef]
- Miranda-CasoLuengo, R.; Lu, J.; Williams, E.J.; Miranda-CasoLuengo, A.A.; Carrington, S.D.; Evans, A.C.; Meijer, W.G. Delayed differentiation of vaginal and uterine microbiomes in dairy cows developing postpartum endometritis. PLoS ONE 2019, 14, e0200974. [Google Scholar] [CrossRef]
- Wang, M.-L.; Liu, M.-C.; Xu, J.; An, L.-G.; Wang, J.-F.; Zhu, Y.-H. Uterine microbiota of dairy cows with clinical and subclinical endometritis. Front. Microbiol. 2018, 9, 2691. [Google Scholar] [CrossRef]
- Piersanti, R.L.; Zimpel, R.; Molinari, P.C.; Dickson, M.J.; Ma, Z.; Jeong, K.C.; Santos, J.E.; Sheldon, I.M.; Bromfield, J.J. A model of clinical endometritis in Holstein heifers using pathogenic Escherichia coli and Trueperella pyogenes. J. Dairy Sci. 2019, 102, 2686–2697. [Google Scholar] [CrossRef]
- Carneiro, L.C.; Cronin, J.G.; Sheldon, I.M. Mechanisms linking bacterial infections of the bovine endometrium to disease and infertility. Reprod. Biol. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, G.; Li, X.; Guan, Y.; Wang, Y.; Yuan, X.; Sun, G.; Wang, Z.; Li, X. Inflammatory mechanism of rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef] [PubMed]
- Pechová, A.; Nečasová, A. The relationship between subclinical ketosis and ruminal dysfunction in dairy cows. Ann. Anim. Sci. 2018, 18, 955–971. [Google Scholar] [CrossRef]
- Elghandour, M.M.; Khusro, A.; Adegbeye, M.J.; Tan, Z.; Abu Hafsa, S.; Greiner, R.; Ugbogu, E.; Anele, U.Y.; Salem, A. Dynamic role of single-celled fungi in ruminal microbial ecology and activities. J. Appl. Microbiol. 2020, 128, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Abdela, N. Sub-acute ruminal acidosis (SARA) and its consequence in dairy cattle: A review of past and recent research at global prospective. Achiev. Life Sci. 2016, 10, 187–196. [Google Scholar] [CrossRef]
- Mohammed, R.; Vyas, D.; Yang, W.; Beauchemin, K. Changes in the relative population size of selected ruminal bacteria following an induced episode of acidosis in beef heifers receiving viable and non-viable active dried yeast. J. Appl. Microbiol. 2017, 122, 1483–1496. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Tun, H.; Derakhshani, H.; Moossavi, S.; Plaizier, J. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Li, S.; Tun, H.M.; Khafipour, E. Nutritional models of experimentally-induced subacute ruminal acidosis (SARA) differ in their impact on rumen and hindgut bacterial communities in dairy cows. Front. Microbiol. 2017, 7, 2128. [Google Scholar] [CrossRef]
- Ogunade, I.; Pech-Cervantes, A.; Schweickart, H. Metatranscriptomic analysis of sub-acute ruminal acidosis in beef cattle. Animals 2019, 9, 232. [Google Scholar] [CrossRef]
- Zeineldin, M.; Barakat, R.; Elolimy, A.; Salem, A.Z.; Elghandour, M.M.; Monroy, J.C. Synergetic action between the rumen microbiota and bovine health. Microb. Pathog. 2018, 124, 106–115. [Google Scholar] [CrossRef]
- Brown, D. Optimising rumen health and the effect this will have on ketosis. Livestock 2018, 23, 174–178. [Google Scholar] [CrossRef]
- Kroezen, V.; Miglior, F.; Schenkel, F.; Squires, J. Development of a genetic marker panel for ketosis in dairy cattle. J. Anim. Sci. 2016, 94, 233–234. [Google Scholar] [CrossRef]
- Rodriguez-Jimenez, S.; Haerr, K.; Trevisi, E.; Loor, J.; Cardoso, F.; Osorio, J. Prepartal standing behavior as a parameter for early detection of postpartal subclinical ketosis associated with inflammation and liver function biomarkers in peripartal dairy cows. J. Dairy Sci. 2018, 101, 8224–8235. [Google Scholar] [CrossRef] [PubMed]
- Canning, P.C.; Srinagesh, S.; Manibusan, A.; Pinkstaff, J.; Knudsen, N. Bovine fibroblast growth factor 21 and ketosis in dairy cattle. U. S. Pat. Appl. Publ. 2019, 16, 325–345. [Google Scholar]
- Huang, Y.; Li, Y.; He, B.; Hu, J.; Ali Mohsin, M.; Yu, H.; Wang, P.; Zhang, P.; Du, Y.; Huang, L. The Influence of Ketosis on the Rectal Microbiome of Chinese Holstein Cows. Pak. Vet. J. 2019, 39, 175–180. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 3320–3333. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Wan, Y.; Hu, T.; Liu, L.; Yang, S.; Gong, Z.; Zeng, Q.; Wei, Y.; Yang, W. A novel postbiotic from Lactobacillus rhamnosus GG with a beneficial effect on intestinal barrier function. Front. Microbiol. 2019, 10, 477. [Google Scholar] [CrossRef]
- Collins, F.W.; Rea, M.C.; Hill, C.; Ross, R.P. 11 Antimicrobials from Lactic. Lactic Acid Bacteria: Microbiological and Functional Aspects. In Lactic Acid Bacteria; Vinderola, G., Ouwehand, A., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 151–174. [Google Scholar]
- Anas, M.; Ahmed, K.; Mebrouk, K. Study of the antimicrobial and probiotic effect of Lactobacillus plantarum isolated from raw goat’s milk from the region of Western Algeria. Int. J. Sci. Es Basic Appl. Res. 2014, 13, 18–27. [Google Scholar]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. 2015, ME14176. [Google Scholar] [CrossRef] [PubMed]
- Suryadi, U.; Nugraheni, Y.R.; Prasetyo, A.F.; Awaludin, A. Evaluation of effects of a novel probiotic feed supplement on the quality of broiler meat. Vet. World 2019, 12, 1775. [Google Scholar] [CrossRef]
- Goto, H.; Qadis, A.Q.; Kim, Y.-H.; Ikuta, K.; Ichijo, T.; Sato, S. Effects of a bacterial probiotic on ruminal pH and volatile fatty acids during subacute ruminal acidosis (SARA) in cattle. J. Vet. Med. Sci. 2016, 16-0211. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rainard, P.; Foucras, G. A critical appraisal of probiotics for mastitis control. Front. Vet. Sci. 2018, 5, 251. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, A.; Warda, A.K.; Hill, C.; Ross, R.P. Non-antibiotic microbial solutions for bovine mastitis–live biotherapeutics, bacteriophage, and phage lysins. Crit. Rev. Microbiol. 2019, 45, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.; Berardo, N.; Giraudo, J.; Nader-Macías, M.; Bogni, C. Bovine mastitis prevention: Humoral and cellular response of dairy cows inoculated with lactic acid bacteria at the dry-off period. Benef. Microbes 2017, 8, 589–596. [Google Scholar] [CrossRef]
- Souza, R.F.S.; Rault, L.; Seyffert, N.; Azevedo, V.; Le Loir, Y.; Even, S. Lactobacillus casei BL23 modulates the innate immune response in Staphylococcus aureus-stimulated bovine mammary epithelial cells. Benef. Microbes 2018, 9, 985–995. [Google Scholar] [CrossRef]
- Wallis, J.K.; Krömker, V.; Paduch, J.-H. Biofilm formation and adhesion to bovine udder epithelium of potentially probiotic lactic acid bacteria. AIMS Microbiol. 2018, 4, 209. [Google Scholar] [CrossRef]
- Wallis, J.K.; Krömker, V.; Paduch, J.-H. Biofilm challenge: Lactic acid bacteria isolated from bovine udders versus Staphylococci. Foods 2019, 8, 79. [Google Scholar] [CrossRef]
- Armas, F.; Camperio, C.; Marianelli, C. In vitro assessment of the probiotic potential of Lactococcus lactis LMG 7930 against ruminant mastitis-causing pathogens. PLoS ONE 2017, 12, e0169543. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.S.; Frola, I.D.; Natanael, B.; Gobelli, D.; Nader-Macias, M.E.; Bogni, C.I. In vitro characterization of lactic acid bacteria isolated from bovine milk as potential probiotic strains to prevent bovine mastitis. Probiotics Antimicrob. Proteins 2019, 11, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ren, Y.; Xi, X.; Huang, W.; Zhang, H. A novel lactobacilli-based teat disinfectant for improving bacterial communities in the milks of cow teats with subclinical mastitis. Front. Microbiol. 2017, 8, 1782. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Alexander, T.W.; Holman, D.B.; Schwinghamer, T.; Timsit, E. Intranasal Bacterial Therapeutics Reduce Colonization by the Respiratory Pathogen Mannheimia haemolytica in Dairy Calves. Msystems 2020, 5, e00629-19. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Nie, Y.; Wang, M.; Xu, H.-X.; Ma, D.-L.; Liang, J.-L.; Wu, X.-L. Mono-homologous linear DNA recombination by the non-homologous end-joining pathway as a novel and simple gene inactivation method: A proof of concept study in Dietzia sp. DQ12-45-1b. bioRxiv 2018, 300723. [Google Scholar] [CrossRef]
- Click, R.E. A 60-day probiotic protocol with Dietzia subsp. C79793-74 prevents development of Johne’s disease parameters after in utero and/or neonatal MAP infection. Virulence 2011, 2, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Sabo, S.; Villalobos, E.F.; Meireles, A.C. Impact of Probiotics on Animal Health. Lactic Acid Bacteria: A Functional Approach; CRC Press: Boca Raton, FL, USA, 2020; pp. 261–290. [Google Scholar] [CrossRef]
- Appiah, M.O.; Wang, J.; Lu, W. Microflora in the Reproductive Tract of Cattle: A Review (Running Title: The Microflora and Bovine Reproductive Tract). Agriculture 2020, 10, 232. [Google Scholar] [CrossRef]
- Deng, Q.; Odhiambo, J.F.; Farooq, U.; Lam, T.; Dunn, S.M.; Ametaj, B.N. Intravaginal lactic acid bacteria modulated local and systemic immune responses and lowered the incidence of uterine infections in periparturient dairy cows. PLoS ONE 2015, 10, e0124167. [Google Scholar] [CrossRef]
- Genís, S.; Bach, À.; Fàbregas, F.; Arís, A. Potential of lactic acid bacteria at regulating Escherichia coli infection and inflammation of bovine endometrium. Theriogenology 2016, 85, 625–637. [Google Scholar] [CrossRef]
- Genís, S.; Sánchez-Chardi, A.; Bach, À.; Fàbregas, F.; Arís, A. A combination of lactic acid bacteria regulates Escherichia coli infection and inflammation of the bovine endometrium. J. Dairy Sci. 2017, 100, 479–492. [Google Scholar] [CrossRef]
- Boranbayeva, T.; Karahan, A.G.; Tulemissova, Z.; Myktybayeva, R.; Özkaya, S. Properties of a New Probiotic Candidate and Lactobacterin-TK 2 Against Diarrhea in Calves. Probiotics Antimicrob. Proteins 2020, 12, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Otsuka, M.; Nishi, K.; Nishi, Y.; Tsukano, K.; Noda, J.; Higuchi, H.; Suzuki, K. Evaluation of probiotic therapy for calf diarrhea with serum diamine oxidase activity as an indicator. Jpn. J. Vet. Res. 2019, 67, 305–311. [Google Scholar]
- Renaud, D.; Kelton, D.; Weese, J.; Noble, C.; Duffield, T. Evaluation of a multispecies probiotic as a supportive treatment for diarrhea in dairy calves: A randomized clinical trial. J. Dairy Sci. 2019, 102, 4498–4505. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.C.; Chiaraviglio, J.; Bru, E.; De Chazal, L.; Santos, V.; Nader-Macías, M. Effect of milk fermented with lactic acid bacteria on diarrheal incidence, growth performance and microbiological and blood profiles of newborn dairy calves. Probiotics Antimicrob. Proteins 2018, 10, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Bukhari, S.M.; Andleeb, S.; Ali, M.; Raza, S.; Nawaz, M.A.; Hussain, T.; Rahman, S.U.; Shah, S.S. Bacteriophages: An overview of the control strategies against multiple bacterial infections in different fields. J. Basic Microbiol. 2019, 59, 123–133. [Google Scholar] [CrossRef]
- Altamirano, F.L.G.; Barr, J.J. Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar]
- Liang, X.; Radosevich, M. Commentary: A host-produced quorum-sensing autoinducer controls a phage lysis-lysogeny decision. Front. Microbiol. 2019, 10, 1201. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Górski, A.; Międzybrodzki, R.; Borysowski, J. Phage Therapy: A Practical Approach; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Zduńczyk, S.; Janowski, T. Bacteriophages and associated endolysins in therapy and prevention of mastitis and metritis in cows: Current knowledge. Anim. Reprod. Sci. 2020, 218, 106504. [Google Scholar] [CrossRef]
- Hamza, A.; Perveen, S.; Abbas, Z.; Rehman, S.U. The lytic SA phage demonstrate bactericidal activity against mastitis causing Staphylococcus aureus. Open Life Sci. 2016, 11, 39–45. [Google Scholar] [CrossRef]
- Tahir, A.; Asif, M.; Abbas, Z.; ur Rehman, S. Three bacteriophages SA, SA2 and SNAF can control growth of milk isolated Staphylococcal species. Pak. J. Zool. 2017, 49, 425–759. [Google Scholar] [CrossRef]
- Ganaie, M.; Qureshi, S.; Kashoo, Z.; Wani, S.; Hussain, M.; Kumar, R.; Maqbool, R.; Sikander, P.; Banday, M.; Malla, W. Isolation and characterization of two lytic bacteriophages against Staphylococcus aureus from India: Newer therapeutic agents against Bovine mastitis. Vet. Res. Commun. 2018, 42, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Titze, I.; Lehnherr, T.; Lehnherr, H.; Krömker, V. Efficacy of bacteriophages against Staphylococcus aureus isolates from bovine mastitis. Pharmaceuticals 2020, 13, 35. [Google Scholar] [CrossRef]
- Iwano, H.; Inoue, Y.; Takasago, T.; Kobayashi, H.; Furusawa, T.; Taniguchi, K.; Fujiki, J.; Yokota, H.; Usui, M.; Tanji, Y. Bacteriophage ΦSA012 has a broad host range against Staphylococcus aureus and effective lytic capacity in a mouse mastitis model. Biology 2018, 7, 8. [Google Scholar] [CrossRef]
- Geng, H.; Zou, W.; Zhang, M.; Xu, L.; Liu, F.; Li, X.; Wang, L.; Xu, Y. Evaluation of phage therapy in the treatment of Staphylococcus aureus-induced mastitis in mice. Folia Microbiol. 2020, 65, 339–351. [Google Scholar] [CrossRef]
- Porter, J.; Anderson, J.; Carter, L.; Donjacour, E.; Paros, M. In vitro evaluation of a novel bacteriophage cocktail as a preventative for bovine coliform mastitis. J. Dairy Sci. 2016, 99, 2053–2062. [Google Scholar] [CrossRef]
- Da Silva Duarte, V.; Dias, R.S.; Kropinski, A.M.; Campanaro, S.; Treu, L.; Siqueira, C.; Vieira, M.S.; da Silva Paes, I.; Santana, G.R.; Martins, F. Genomic analysis and immune response in a murine mastitis model of vB_EcoM-UFV13, a potential biocontrol agent for use in dairy cows. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Da Silva Duarte, V.; Dias, R.S.; Kropinski, A.M.; da Silva Xavier, A.; Ferro, C.G.; Vidigal, P.M.; da Silva, C.C.; de Paula, S.O. A T4virus prevents biofilm formation by Trueperella pyogenes. Vet. Microbiol. 2018, 218, 45–51. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, H.; Bao, H.; Wang, X.; Wang, R. The lytic activity of recombinant phage lysin LysKΔamidase against staphylococcal strains associated with bovine and human infections in the Jiangsu province of China. Res. Vet. Sci. 2017, 111, 113–119. [Google Scholar] [CrossRef]
- Fan, J.; Zeng, Z.; Mai, K.; Yang, Y.; Feng, J.; Bai, Y.; Sun, B.; Xie, Q.; Tong, Y.; Ma, J. Preliminary treatment of bovine mastitis caused by Staphylococcus aureus, with trx-SA1, recombinant endolysin of S. aureus bacteriophage IME-SA1. Vet. Microbiol. 2016, 191, 65–71. [Google Scholar] [CrossRef]
- Scholte, C.; Nelson, D.; Garcia, M.; Linden, S.; Elsasser, T.; Kahl, S.; Qu, Y.; Moyes, K. Recombinant bacteriophage endolysin PlyC is nontoxic and does not alter blood neutrophil oxidative response in lactating dairy cows. J. Dairy Sci. 2018, 101, 6419–6423. [Google Scholar] [CrossRef]
- Frizzo, L.S.; Signorini, M.L.; Rosmini, M.R. Probiotics and prebiotics for the health of cattle. In Probiotics and Prebiotics in Animal Health and Food Safety; Di Gioia, D., Biavati, B., Eds.; Springer: Cham, Switzerland, 2018; pp. 155–174. [Google Scholar]
- Smulski, S.; Turlewicz-Podbielska, H.; Wylandowska, A.; Włodarek, J. Non-antibiotic possibilities in prevention and treatment of calf diarrhoea. J. Vet. Res. 2020, 64, 119–126. [Google Scholar] [CrossRef]
- Singh, A.; Kerketta, S.; Yogi, R.; Kumar, A.; Ojha, L. Prebiotics: The new feed supplement for dairy calf. Int. J. Livest Res. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Grispoldi, L.; Bertero, F.; Franceschini, S.; Mastrosimone, F.; Sechi, P.; Iulietto, M.F.; Ceccarelli, M.; Cenci-Goga, B.T. Prevalence and characterisation of shigatoxigenic Escherichia coli isolated from beef cattle fed with prebiotics. Ital. J. Food Saf. 2017, 6, 4. [Google Scholar] [CrossRef]
- Al-Shawi, S.G.; Dang, D.S.; Yousif, A.Y.; Al-Younis, Z.K.; Najm, T.A.; Matarneh, S.K. The Potential Use of Probiotics to Improve Animal Health, Efficiency, and Meat Quality: A Review. Agriculture 2020, 10, 452. [Google Scholar] [CrossRef]
- Klein-Jöbstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing reveals diverse fecal microbiota in Simmental calves during early development. Front. Microbiol. 2014, 5, 622. [Google Scholar]
- Oikonomou, G.; Teixeira, A.G.V.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal microbial diversity in pre-weaned dairy calves as described by pyrosequencing of metagenomic 16S rDNA. Associations of Faecalibacterium species with health and growth. PLoS ONE 2013, 8, e63157. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Disease | Probiotic Combination | Reference |
|---|---|---|
| Rumen acidosis | L. plantarum, E.faecium, and C.butyricum | [142] |
| Saccharomycescerevisiae | [124] | |
| Mastitis | L. perolens CRL1724 and L. lactis subsp. lactis CRL1655 | [145] |
| L. casei BL23 | [146] | |
| L. rhamnosusATCC 7469,L. plantarum 2/37,L. Brevis 104/37,L. plantarum 118/37, and L. plantarum 6E | [148] | |
| Lactococcus lactis LMG 7930 | [149] | |
| L. plantarum IMAU80065 and IMAU10155 | [151] | |
| Bovine respiratory disease | L. amylovorus,L. buchneri,L. curvatus, and L. paracasei, | [36,152] |
| Dietzia spp. | [153] | |
| Dietzia subsp. C79793-74 | [154] | |
| Metritis | Lactobacillus sakei FUA 3089, Pediococcus acidilactici FUA 3138, and FUA 3140 | [157] |
| Lactobacillus rhamnosus,Pediococcus acidilactici,Lactobacillus reuteri, and Lactobacillus sakei | [158] | |
| Calf diarrhoea | Pediococcus acidilactici,Lactobacillus acidophilus,Lactobacillus casei,Enterococcus faecium, and Bifidobacterium bifidum | [162] |
| Lactobacillus murinus CRL 1695, Lactobacillus mucosae CRL 1696, Lactobacillus johnsonii CRL1693, and Lactobacillus salivarius CRL 1702 | [163] | |
| Bacillus mesentericus,Clostridium butyricum and Enterococcus faecalis | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, A.; Batool, A.; Arif, S. Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes. Ruminants 2022, 2, 134-156. https://doi.org/10.3390/ruminants2010009
Khalil A, Batool A, Arif S. Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes. Ruminants. 2022; 2(1):134-156. https://doi.org/10.3390/ruminants2010009
Chicago/Turabian StyleKhalil, Alaa, Aamina Batool, and Sania Arif. 2022. "Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes" Ruminants 2, no. 1: 134-156. https://doi.org/10.3390/ruminants2010009
APA StyleKhalil, A., Batool, A., & Arif, S. (2022). Healthy Cattle Microbiome and Dysbiosis in Diseased Phenotypes. Ruminants, 2(1), 134-156. https://doi.org/10.3390/ruminants2010009