
PGT-SR: A Comprehensive Overview and a Requiem for the Interchromosomal Effect



Abstract
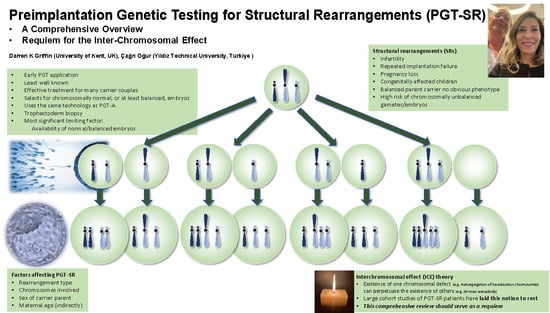
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
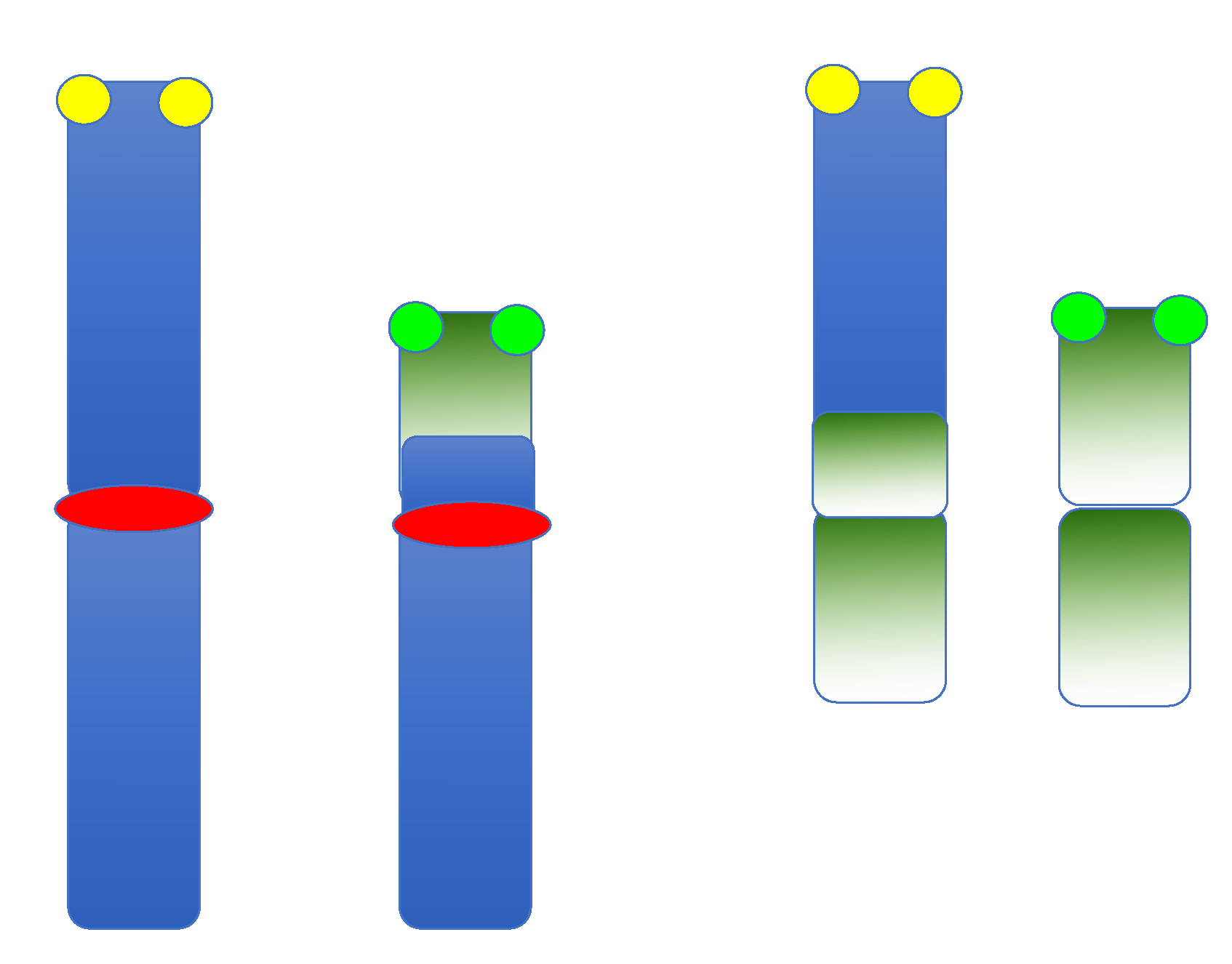
2. Balanced SRs in Overview
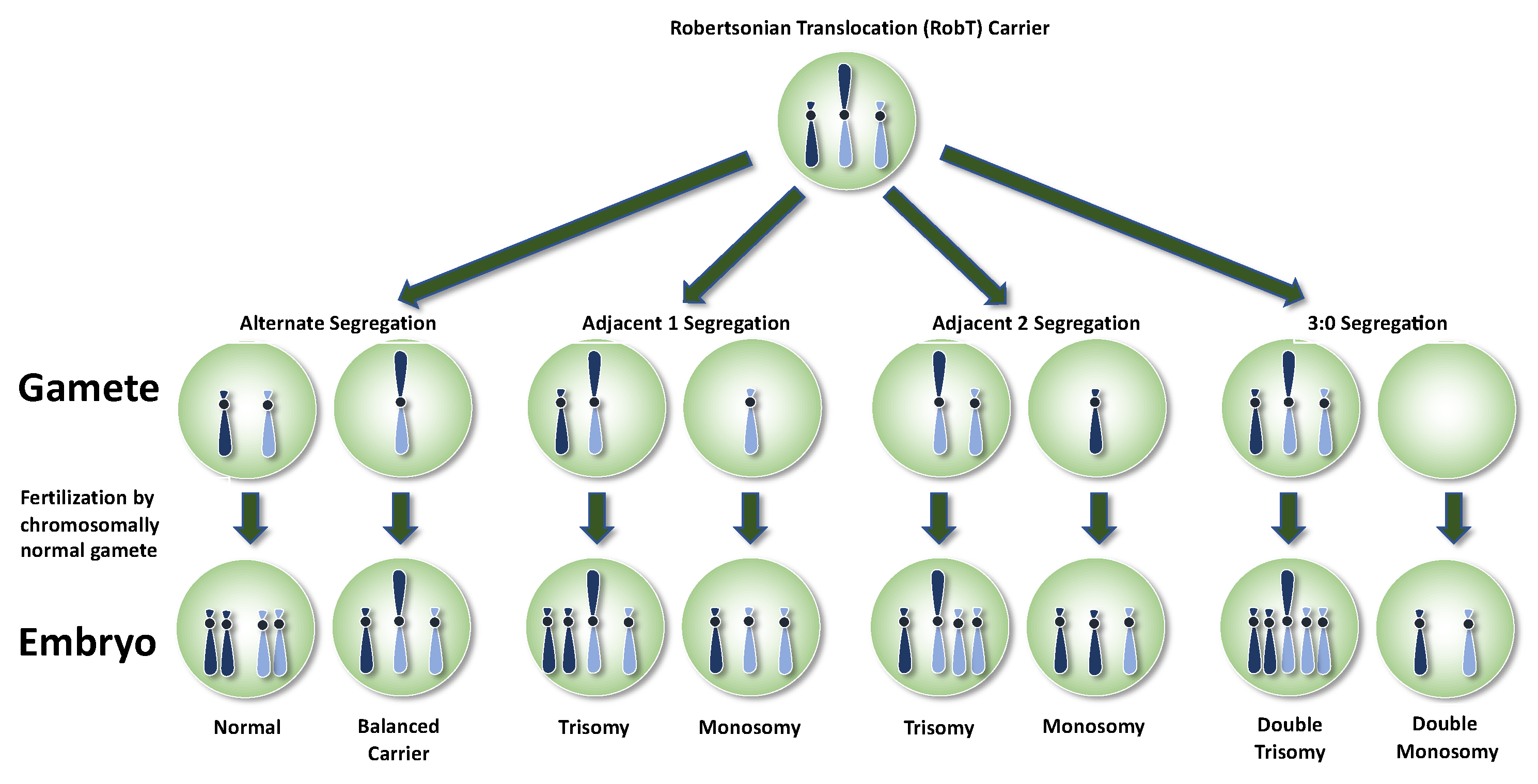
3. Robertsonian Translocations
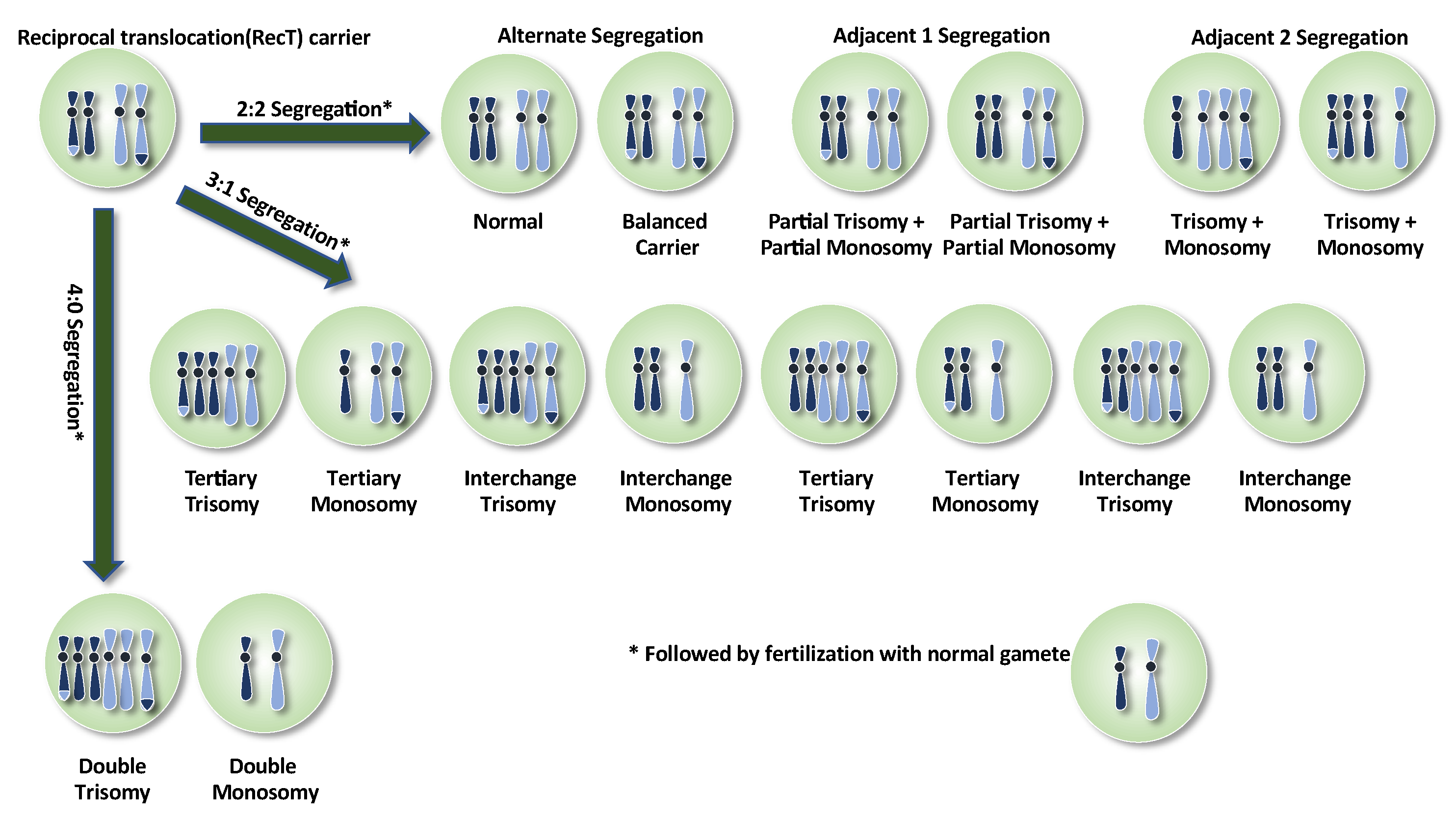
4. Reciprocal Translocations
5. Inversions
6. Insertional Translocations and Complex Chromosomal Rearrangements
7. Practicing PGT-SR
8. Invasive Sampling Methods for PGT-SR
9. Minimally Invasive Sampling Methods for PGT-SR
10. FISH for PGT-SR
11. STR-Typing for PGT-SR
12. Comparative Genomic Hybridization (CGH) and Array Comparative Genomic Hybridization (aCGH)
13. Karyomapping and SNP Arrays for PGT-SR
14. Next-Generation Sequencing (NGS) for PGT-SR
15. Towards Universal PGT
16. Should We Deselect or De-Prioritize Carrier Embryos?
17. Developmental Characteristics of the Embryo and PGT-SR
18. Ovarian Response and Its Relevance to PGT-SR
19. Meiotic Segregation Patterns and the Number of Chromosomally Balanced Embryos Available for PGT-SR
20. The Time of Biopsy and Maternal Age: No Effect on Chromosome Segregation, but Relevant to PGT-SR Nonetheless
21. Aneuploidy and Chromosomal Mosaicism
22. Requiem for the Interchromosomal Effect (ICE)
23. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Weckselblatt, B.; Hermetz, K.E.; Rudd, M.K. Unbalanced Translocations Arise from Diverse Mutational Mechanisms Including Chromothripsis. Genome Res. 2015, 25, 937–947. [Google Scholar] [CrossRef]
- De Braekeleer, M.; Dao, T.N. Cytogenetic Studies in Couples Experiencing Repeated Pregnancy Losses. Hum. Reprod. 1990, 5, 519–528. [Google Scholar] [CrossRef] [PubMed]
- De Braekeleer, M.; Dao, T.N. Cytogenetic Studies in Male Infertility: A Review. Hum. Reprod. 1991, 6, 245–250. [Google Scholar] [CrossRef]
- Shah, K.; Sivapalan, G.; Gibbons, N.; Tempest, H.; Griffin, D.K. The Genetic Basis of Infertility. Reproduction 2003, 126, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, J. Autosomal Disorders. Pediatrics 1963, 32, 326–337. [Google Scholar] [CrossRef]
- Griffin, D.K. Fluorescent Molecular Cytogenetics: Preimplantation Diagnosis, Colorectal Cancer and Mapping Chromosome 9. Ph.D. Thesis, University College London, London, UK, 1992. [Google Scholar]
- Harper, J.C.; Wilton, L.; Traeger-Synodinos, J.; Goossens, V.; Moutou, C.; SenGupta, S.B.; Pehlivan Budak, T.; Renwick, P.; De Rycke, M.; Geraedts, J.P.M.; et al. The ESHRE PGD Consortium: 10 Years of Data Collection. Hum. Reprod. Update 2012, 18, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Ogur, C. Chromosomal Analysis in IVF: Just How Useful Is It? Reproduction 2018, 156, F29–F50. [Google Scholar] [CrossRef] [PubMed]
- Ogur, C.; Griffin, D.K. Preimplantation Genetic Testing for Structural Rearrangements. In Preimplantation Genetic Testing. Recent Advances in Reproductive Medicine; Griffin, D.K., Harton, G.L., Eds.; CRC Press: Boca Raton, UK, 2020; pp. 49–76. [Google Scholar] [CrossRef]
- Lieber, M.R. The Mechanism of Double-Strand DNA Break Repair by the Nonhomologous DNA End-Joining Pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef]
- Hurles, M.E.; Lupski, J.R. Recombination hotspots in nonallelic homologous recombination. In Genomic Disorders; Lupski, J.R., Stankiewicz, P., Eds.; Humana Press: Totowa, NJ, USA, 2006; pp. 341–355. [Google Scholar] [CrossRef]
- Ou, Z.Z.; Kochmar, S.; Yatsenko, S.A.; Woerner, A.C.; Acquaro, R.; Ortiz, D.; Surti, U.; Hu, J. Partial 5p Deletion and Partial 5q Duplication in a Patient with Multiple Congenital Anomalies: A Two-Step Mechanism in Chromosomal Rearrangement Mediated by Non-Allelic Homologous Recombination. Cytogenet. Genome Res. 2018, 156, 65–70. [Google Scholar] [CrossRef]
- Pellestor, F.; Anahory, T.; Lefort, G.; Puechberty, J.; Liehr, T.; Hedon, B.; Sarda, P. Complex Chromosomal Rearrangements: Origin and Meiotic Behavior. Hum. Reprod. Update 2011, 17, 476–494. [Google Scholar] [CrossRef]
- Lee, J.A.; Carvalho, C.M.B.; Lupski, J.R. A DNA Replication Mechanism for Generating Nonrecurrent Rearrangements Associated with Genomic Disorders. Cell 2007, 131, 1235–1247. [Google Scholar] [CrossRef]
- Tucker, J. Low-Dose Ionizing Radiation and Chromosome Translocations: A Review of the Major Considerations for Human Biological Dosimetry. Mutat. Res./Rev. Mutat. Res. 2008, 659, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Hook, E.B.; Schreinemachers, D.M.; Willey, A.M.; Cross, P.K. Inherited Structural Cytogenetic Abnormalities Detected Incidentally in Fetuses Diagnosed Prenatally: Frequency, Parental-Age Associations, Sex-Ratio Trends, and Comparisons with Rates of Mutants. Am. J. Hum. Genet. 1984, 36, 422–443. [Google Scholar] [PubMed]
- Beyazyurek, C.; Ekmekci, C.G.; Sağlam, Y.; Cinar, C.; Kahraman, S. Preimplantation Genetic Diagnosis (PGD) for Extremes—Successful Birth after PGD for a Consanguineous Couple Carrying an Identical Balanced Reciprocal Translocation. Fertil. Steril. 2010, 93, e1–e2413. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, P.A.; Browne, C.; Gregson, N.; Joyce, C.; White, H. Estimates of the Frequency of Chromosome Abnormalities Detectable in Unselected Newborns Using Moderate Levels of Banding. J. Med. Genet. 1992, 29, 103–108. [Google Scholar] [CrossRef]
- Scriven, P.N.; Handyside, A.H.; Ogilvie, C.M. Chromosome Translocations: Segregation Modes and Strategies for Preimplantation Genetic Diagnosis. Prenat. Diagn. 1998, 18, 1437–1449. [Google Scholar] [CrossRef]
- Martin, R.H. A Detailed Method for Obtaining Preparations of Human Sperm Chromosomes. Cytogenet. Cell Genet. 1983, 35, 252–256. [Google Scholar] [CrossRef]
- Lamotte, A.; Martinez, G.; Devillard, F.; Hograindleur, J.-P.; Satre, V.; Coutton, C.; Harbuz, R.; Amblard, F.; Lespinasse, J.; Benchaib, M.; et al. Is Sperm FISH Analysis Still Useful for Robertsonian Translocations? Meiotic Analysis for 23 Patients and Review of the Literature. Basic Clin. Androl. 2018, 28, 5. [Google Scholar] [CrossRef]
- Patassini, C.; Garolla, A.; Bottacin, A.; Menegazzo, M.; Speltra, E.; Foresta, C.; Ferlin, A. Molecular Karyotyping of Human Single Sperm by Array-Comparative Genomic Hybridization. PLoS ONE 2013, 8, e60922. [Google Scholar] [CrossRef]
- Martin, R.H. Cytogenetic Determinants of Male Fertility. Hum. Reprod. Update 2008, 14, 379–390. [Google Scholar] [CrossRef]
- Goldman, A.S.; Hultén, M.A. Analysis of Chiasma Frequency and First Meiotic Segregation in a Human Male Reciprocal Translocation Heterozygote, t(1;11)(P36.3;Q13.1), Using Fluorescence in Situ Hybridisation. Cytogenet. Cell Genet. 1993, 63, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Escudero, T.; Abdelhadi, I.; Sandalinas, M.; Munné, S. Predictive Value of Sperm Fluorescence in Situ Hybridization Analysis on the Outcome of Preimplantation Genetic Diagnosis for Translocations. Fertil. Steril. 2003, 79, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Pettenati, M.J.; Rao, P.N.; Phelan, M.C.; Grass, F.; Rao, K.W.; Cosper, P.; Carroll, A.J.; Elder, F.; Smith, J.L.; Higgins, M.D. Paracentric Inversions in Humans: A Review of 446 Paracentric Inversions with Presentation of 120 New Cases. Am. J. Med. Genet. 1995, 55, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Jaarola, M.; Martin, R.H.; Ashley, T. Direct Evidence for Suppression of Recombination within Two Pericentric Inversions in Humans: A New Sperm-FISH Technique. Am. J. Hum. Genet. 1998, 63, 218–224. [Google Scholar] [CrossRef]
- Martin, R.H. Sperm Chromosome Analysis in a Man Heterozygous for a Paracentric Inversion of Chromosome 14 (Q24.1q32.1). Am. J. Hum. Genet. 1999, 64, 1480–1484. [Google Scholar] [CrossRef][Green Version]
- Bhatt, S.; Moradkhani, K.; Mrasek, K.; Puechberty, J.; Manvelyan, M.; Hunstig, F.; Lefort, G.; Weise, A.; Lespinasse, J.; Sarda, P.; et al. Breakpoint Mapping and Complete Analysis of Meiotic Segregation Patterns in Three Men Heterozygous for Paracentric Inversions. Eur. J. Hum. Genet. 2009, 17, 44–50. [Google Scholar] [CrossRef][Green Version]
- Yapan, C.; Beyazyurek, C.; Ekmekci, C.; Kahraman, S. The Largest Paracentric Inversion, the Highest Rate of Recombinant Spermatozoa. Case Report: 46,XY, Inv(2)(Q21.2q37.3) and Literature Review. Balk. J. Med. Genet. 2014, 17, 55–62. [Google Scholar] [CrossRef]
- Morel, F.; Laudier, B.; Guérif, F.; Couet, M.L.; Royère, D.; Roux, C.; Bresson, J.L.; Amice, V.; De Braekeleer, M.; Douet-Guilbert, N. Meiotic Segregation Analysis in Spermatozoa of Pericentric Inversion Carriers Using Fluorescence In-Situ Hybridization. Hum. Reprod. 2007, 22, 136–141. [Google Scholar] [CrossRef]
- Abdi, A. Prevalence of Chromosome Inversions (Pericentric and Paracentric) in Patients with Recurrent Abortions. SJRM 2017, 2, 45–50. [Google Scholar] [CrossRef]
- Anton, E.; Blanco, J.; Egozcue, J.; Vidal, F. Sperm Studies in Heterozygote Inversion Carriers: A Review. Cytogenet. Genome Res. 2005, 111, 297–304. [Google Scholar] [CrossRef]
- Salaun, G.; Tchirkov, A.; Francannet, C.; Pons, H.; Brugnon, F.; Pebrel-Richard, C.; Gouas, L.; Eymard-Pierre, E.; Vago, P.; Goumy, C. Sperm Meiotic Segregation of a Balanced Interchromosomal Reciprocal Insertion Resulting in Recurrent Spontaneous Miscarriage. Reprod. BioMed. Online 2018, 37, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Melotte, C.; Debrock, S.; D’Hooghe, T.; Fryns, J.P.; Vermeesch, J.R. Preimplantation Genetic Diagnosis for an Insertional Translocation Carrier. Hum. Reprod. 2004, 19, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- Xanthopoulou, L.; Mantzouratou, A.; Mania, A.; Cawood, S.; Doshi, A.; Ranieri, D.M.; Delhanty, J.D. Male and Female Meiotic Behaviour of an Intrachromosomal Insertion Determined by Preimplantation Genetic Diagnosis. Mol. Cytogenet. 2010, 3, 2. [Google Scholar] [CrossRef] [PubMed]
- Scriven, P.N.; Bint, S.M.; Davies, A.F.; Ogilvie, C.M. Meiotic Outcomes of Three-Way Translocations Ascertained in Cleavage-Stage Embryos: Refinement of Reproductive Risks and Implications for PGD. Eur. J. Hum. Genet. 2014, 22, 748–753. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pujol, A.; Durban, M.; Benet, J.; Boiso, I.; Calafell, J.M.; Egozcue, J.; Navarro, J. Multiple Aneuploidies in the Oocytes of Balanced Translocation Carriers: A Preimplantation Genetic Diagnosis Study Using First Polar Body. Reproduction 2003, 126, 701–711. [Google Scholar] [CrossRef]
- Molina Gomes, D.; Hammoud, I.; Bailly, M.; Bergere, M.; Wainer, R.; Selva, J.; Vialard, F. Preconceptional Diagnosis for Robertsonian Translocation as an Alternative to Preimplantation Genetic Diagnosis in Two Situations: A Pilot Study. J. Assist. Reprod. Genet. 2009, 26, 113–117. [Google Scholar] [CrossRef][Green Version]
- Scott, R.T.; Upham, K.M.; Forman, E.J.; Zhao, T.; Treff, N.R. Cleavage-Stage Biopsy Significantly Impairs Human Embryonic Implantation Potential While Blastocyst Biopsy Does Not: A Randomized and Paired Clinical Trial. Fertil. Steril. 2013, 100, 624–630. [Google Scholar] [CrossRef]
- Chatzimeletiou, K.; Petrogiannis, N.; Sioga, A.; Emmanouil-Nikoloussi, E.N.; Panagiotidis, Y.; Prapa, M.; Patrikiou, A.; Filippa, M.; Zervakakou, G.; Papanikolaou, K.; et al. The human embryo following biopsy on day 5 versus day 3: Viability, ultrastructure and spindle/chromosome configurations. Reprod. Biomed. Online 2022, 45, 219–233. [Google Scholar] [CrossRef]
- Magli, M.C.; Pomante, A.; Cafueri, G.; Valerio, M.; Crippa, A.; Ferraretti, A.P.; Gianaroli, L. Preimplantation Genetic Testing: Polar Bodies, Blastomeres, Trophectoderm Cells, or Blastocoelic Fluid? Fertil. Steril. 2016, 105, 676–683.e5. [Google Scholar] [CrossRef]
- Liu, W.; Liu, J.; Du, H.; Ling, J.; Sun, X.; Chen, D. Non-Invasive Pre-Implantation Aneuploidy Screening and Diagnosis of Beta Thalassemia IVSII654 Mutation Using Spent Embryo Culture Medium. Ann. Med. 2017, 49, 319–328. [Google Scholar] [CrossRef]
- Xu, J.; Fang, R.; Chen, L.; Chen, D.; Xiao, J.-P.; Yang, W.; Wang, H.; Song, X.; Ma, T.; Bo, S.; et al. Noninvasive Chromosome Screening of Human Embryos by Genome Sequencing of Embryo Culture Medium for in Vitro Fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, 11907–11912. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Shi, B.; Sagnelli, M.; Yang, D.; Yao, Y.; Li, W.; Shao, L.; Lu, S.; Li, D.; Wang, X. Minimally Invasive Preimplantation Genetic Testing Using Blastocyst Culture Medium. Hum. Reprod. 2019, 34, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Handyside, A.H.; Penketh, R.J.; Winston, R.M.; Delhanty, J.D. Fluorescent In-Situ Hybridization to Interphase Nuclei of Human Preimplantation Embryos with X and Y Chromosome Specific Probes. Hum. Reprod. 1991, 6, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Schrurs, B.M.; Winston, R.M.; Handyside, A.H. Preimplantation Diagnosis of Aneuploidy Using Fluorescent In-Situ Hybridization: Evaluation Using a Chromosome 18-Specific Probe. Hum. Reprod. 1993, 8, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Cassel, M.J.; Munné, S.; Fung, J.; Weier, H.U. Carrier-Specific Breakpoint-Spanning DNA Probes: An Approach to Preimplantation Genetic Diagnosis in Interphase Cells. Hum. Reprod. 1997, 12, 2019–2027. [Google Scholar] [CrossRef]
- Munne, S.; Sandalinas, M.; Escudero, T.; Fung, J.; Gianaroli, L.; Cohen, J. Outcome of Preimplantation Genetic Diagnosis of Translocations. Fertil. Steril. 2000, 73, 1209–1218. [Google Scholar] [CrossRef]
- Munné, S.; Escudero, T.; Fischer, J.; Chen, S.; Hill, J.; Stelling, J.R.; Anna, E. Negligible Interchromosomal Effect in Embryos of Robertsonian Translocation Carriers. Reprod. BioMed. Online 2005, 10, 363–369. [Google Scholar] [CrossRef]
- Velilla, E.; Escudero, T.; Munné, S. Blastomere Fixation Techniques and Risk of Misdiagnosis for Preimplantation Genetic Diagnosis of Aneuploidy. Reprod. BioMed. Online 2002, 4, 210–217. [Google Scholar] [CrossRef]
- Ghevaria, H.; SenGupta, S.; Shmitova, N.; Serhal, P.; Delhanty, J. The Origin and Significance of Additional Aneuploidy Events in Couples Undergoing Preimplantation Genetic Diagnosis for Translocations by Array Comparative Genomic Hybridization. Reprod. BioMed. Online 2016, 32, 178–189. [Google Scholar] [CrossRef][Green Version]
- Verlinsky, Y.; Cieslak, J.; Evsikov, S.; Galat, V.; Kuliev, A. Nuclear Transfer for Full Karyotyping and Preimplantation Diagnosis for Translocations. Reprod. BioMed. Online 2002, 5, 300–305. [Google Scholar] [CrossRef]
- Kuliev, A.; Janzen, J.C.; Zlatopolsky, Z.; Kirillova, I.; Ilkevitch, Y.; Verlinsky, Y. Conversion and Non-Conversion Approach to Preimplantation Diagnosis for Chromosomal Rearrangements in 475 Cycles. Reprod. BioMed. Online 2010, 21, 93–99. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lim, C.K.; Cho, J.W.; Song, I.O.; Kang, I.S.; Yoon, Y.-D.; Jun, J.H. Estimation of Chromosomal Imbalances in Preimplantation Embryos from Preimplantation Genetic Diagnosis Cycles of Reciprocal Translocations with or without Acrocentric Chromosomes. Fertil. Steril. 2008, 90, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.S.; Cho, J.W.; Park, S.Y.; Kim, J.Y.; Koong, M.K.; Song, I.O.; Kang, I.S.; Lim, C.K. Clinical Outcomes of Preimplantation Genetic Diagnosis (PGD) and Analysis of Meiotic Segregation Modes in Reciprocal Translocation Carriers. Am. J. Med. Genet. 2010, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Gianaroli, L. Possible Interchromosomal Effect in Embryos Generated by Gametes from Translocation Carriers. Hum. Reprod. 2002, 17, 3201–3207. [Google Scholar] [CrossRef] [PubMed]
- Pujol, A.; Benet, J.; Staessen, C.; Van Assche, E.; Campillo, M.; Egozcue, J.; Navarro, J. The Importance of Aneuploidy Screening in Reciprocal Translocation Carriers. Reproduction 2006, 131, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Keymolen, K.; Staessen, C.; Verpoest, W.; Liebaers, I.; Bonduelle, M. Preimplantation Genetic Diagnosis in Female and Male Carriers of Reciprocal Translocations: Clinical Outcome until Delivery of 312 Cycles. Eur. J. Hum. Genet. 2012, 20, 376–380. [Google Scholar] [CrossRef]
- Franssen, M.T.M.; Musters, A.M.; van der Veen, F.; Repping, S.; Leschot, N.J.; Bossuyt, P.M.M.; Goddijn, M.; Korevaar, J.C. Reproductive Outcome after PGD in Couples with Recurrent Miscarriage Carrying a Structural Chromosome Abnormality: A Systematic Review. Hum. Reprod. Update 2011, 17, 467–475. [Google Scholar] [CrossRef]
- Iews, M.; Tan, J.; Taskin, O.; Alfaraj, S.; AbdelHafez, F.F.; Abdellah, A.H.; Bedaiwy, M.A. Does Preimplantation Genetic Diagnosis Improve Reproductive Outcome in Couples with Recurrent Pregnancy Loss Owing to Structural Chromosomal Rearrangement? A Systematic Review. Reprod. BioMed. Online 2018, 36, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Traversa, M.V.; Carey, L.; Leigh, D. A Molecular Strategy for Routine Preimplantation Genetic Diagnosis in Both Reciprocal and Robertsonian Translocation Carriers. Mol. Hum. Reprod. 2010, 16, 329–337. [Google Scholar] [CrossRef][Green Version]
- Fiorentino, F.; Kokkali, G.; Biricik, A.; Stavrou, D.; Ismailoglu, B.; De Palma, R.; Arizzi, L.; Harton, G.; Sessa, M.; Pantos, K. Polymerase Chain Reaction-Based Detection of Chromosomal Imbalances on Embryos: The Evolution of Preimplantation Genetic Diagnosis for Chromosomal Translocations. Fertil. Steril. 2010, 94, 2001–2011.e6. [Google Scholar] [CrossRef]
- Malmgren, H.; Sahlén, S.; Inzunza, J.; Aho, M.; Rosenlund, B.; Fridström, M.; Hovatta, O.; Ahrlund-Richter, L.; Nordenskjöld, M.; Blennow, E. Single Cell CGH Analysis Reveals a High Degree of Mosaicism in Human Embryos from Patients with Balanced Structural Chromosome Aberrations. Mol. Hum. Reprod. 2002, 8, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Le Caignec, C.; Spits, C.; Sermon, K.; De Rycke, M.; Thienpont, B.; Debrock, S.; Staessen, C.; Moreau, Y.; Fryns, J.-P.; Van Steirteghem, A.; et al. Single-Cell Chromosomal Imbalances Detection by Array CGH. Nucleic Acids Res. 2006, 34, e68. [Google Scholar] [CrossRef] [PubMed]
- Kallioniemi, A.; Kallioniemi, O.P.; Sudar, D.; Rutovitz, D.; Gray, J.W.; Waldman, F.; Pinkel, D. Comparative Genomic Hybridization for Molecular Cytogenetic Analysis of Solid Tumors. Science 1992, 258, 818–821. [Google Scholar] [CrossRef] [PubMed]
- Wilton, L. Preimplantation Genetic Diagnosis and Chromosome Analysis of Blastomeres Using Comparative Genomic Hybridization. Hum. Reprod. Update 2005, 11, 33–41. [Google Scholar] [CrossRef]
- Rius, M.; Obradors, A.; Daina, G.; Ramos, L.; Pujol, A.; Martínez-Passarell, O.; Marquès, L.; Oliver-Bonet, M.; Benet, J.; Navarro, J. Detection of Unbalanced Chromosome Segregations in Preimplantation Genetic Diagnosis of Translocations by Short Comparative Genomic Hibridization. Fertil. Steril. 2011, 96, 134–142. [Google Scholar] [CrossRef]
- Hu, L.; Cheng, D.; Gong, F.; Lu, C.; Tan, Y.; Luo, K.; Wu, X.; He, W.; Xie, P.; Feng, T.; et al. Reciprocal Translocation Carrier Diagnosis in Preimplantation Human Embryos. EBioMedicine 2016, 14, 139–147. [Google Scholar] [CrossRef]
- Fiorentino, F.; Spizzichino, L.; Bono, S.; Biricik, A.; Kokkali, G.; Rienzi, L.; Ubaldi, F.M.; Iammarrone, E.; Gordon, A.; Pantos, K. PGD for Reciprocal and Robertsonian Translocations Using Array Comparative Genomic Hybridization. Hum. Reprod. 2011, 26, 1925–1935. [Google Scholar] [CrossRef]
- Ramos, L.; del Rey, J.; Daina, G.; García-Aragonés, M.; Armengol, L.; Fernandez-Encinas, A.; Parriego, M.; Boada, M.; Martinez-Passarell, O.; Martorell, M.R.; et al. Oligonucleotide Arrays vs. Metaphase-Comparative Genomic Hybridisation and BAC Arrays for Single-Cell Analysis: First Applications to Preimplantation Genetic Diagnosis for Robertsonian Translocation Carriers. PLoS ONE 2014, 9, e113223. [Google Scholar] [CrossRef]
- Christodoulou, C.; Dheedene, A.; Heindryckx, B.; van Nieuwerburgh, F.; Deforce, D.; De Sutter, P.; Menten, B.; Van den Abbeel, E. Preimplantation Genetic Diagnosis for Chromosomal Rearrangements with the Use of Array Comparative Genomic Hybridization at the Blastocyst Stage. Fertil. Steril. 2017, 107, 212–219.e3. [Google Scholar] [CrossRef][Green Version]
- Alfarawati, S.; Fragouli, E.; Colls, P.; Wells, D. First Births after Preimplantation Genetic Diagnosis of Structural Chromosome Abnormalities Using Comparative Genomic Hybridization and Microarray Analysis. Hum. Reprod. 2011, 26, 1560–1574. [Google Scholar] [CrossRef]
- Fodina, V.; Dudorova, A.; Alksere, B.; Dzalbs, A.; Vedmedovska, N.; Andersone, S.; Una, C.; Erenpreiss, J.; Dace, B. The application of PGT-A for carriers of balanced structural chromosomal rearrangements. Gynecol. Endocrinol. 2019, 35 (Suppl. S1), 18–23. [Google Scholar] [CrossRef] [PubMed]
- Handyside, A.H.; Harton, G.L.; Mariani, B.; Thornhill, A.R.; Affara, N.; Shaw, M.-A.; Griffin, D.K. Karyomapping: A Universal Method for Genome Wide Analysis of Genetic Disease Based on Mapping Crossovers between Parental Haplotypes. J. Med. Genet. 2010, 47, 651–658. [Google Scholar] [CrossRef]
- Treff, N.R.; Thompson, K.; Rafizadeh, M.; Chow, M.; Morrison, L.; Tao, X.; Garnsey, H.; Reda, C.V.; Metzgar, T.L.; Neal, S.; et al. SNP Array-Based Analyses of Unbalanced Embryos as a Reference to Distinguish between Balanced Translocation Carrier and Normal Blastocysts. J. Assist. Reprod. Genet. 2016, 33, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Sundheimer, L.W.; Liu, L.; Buyalos, R.P.; Hubert, G.; Al-Safi, Z.; Shamonki, M. Diagnosis of Parental Balanced Reciprocal Translocations by Trophectoderm Biopsy and Comprehensive Chromosomal Screening. J. Assist. Reprod. Genet. 2018, 35, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lei, C.; Wu, J.; Zhou, J.; Sun, H.; Fu, J.; Sun, Y.; Sun, X.; Lu, D.; Zhang, Y. The Establishment and Application of Preimplantation Genetic Haplotyping in Embryo Diagnosis for Reciprocal and Robertsonian Translocation Carriers. BMC Med. Genom. 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Zamani Esteki, M.; Dimitriadou, E.; Mateiu, L.; Melotte, C.; Van der Aa, N.; Kumar, P.; Das, R.; Theunis, K.; Cheng, J.; Legius, E.; et al. Concurrent Whole-Genome Haplotyping and Copy-Number Profiling of Single Cells. Am. J. Hum. Genet. 2015, 96, 894–912. [Google Scholar] [CrossRef] [PubMed]
- Treff, N.R.; Northrop, L.E.; Kasabwala, K.; Su, J.; Levy, B.; Scott, R.T. Single Nucleotide Polymorphism Microarray-Based Concurrent Screening of 24-Chromosome Aneuploidy and Unbalanced Translocations in Preimplantation Human Embryos. Fertil. Steril. 2011, 95, e1–e2. [Google Scholar] [CrossRef]
- Van Uum, C.M.; Stevens, S.J.; Dreesen, J.C.; Drüsedau, M.; Smeets, H.J.; Hollanders-Crombach, B.; de Die-Smulders, C.E.; Geraedts, J.P.; Engelen, J.J.; Coonen, E. SNP Array-Based Copy Number and Genotype Analyses for Preimplantation Genetic Diagnosis of Human Unbalanced Translocations. Eur. J. Hum. Genet. 2012, 20, 938–944. [Google Scholar] [CrossRef][Green Version]
- Tan, Y.-Q.; Tan, K.; Zhang, S.-P.; Gong, F.; Cheng, D.-H.; Xiong, B.; Lu, C.-F.; Tang, X.-C.; Luo, K.-L.; Lin, G.; et al. Single-Nucleotide Polymorphism Microarray-Based Preimplantation Genetic Diagnosis Is Likely to Improve the Clinical Outcome for Translocation Carriers. Hum. Reprod. 2013, 28, 2581–2592. [Google Scholar] [CrossRef]
- Tobler, K.J.; Brezina, P.R.; Benner, A.T.; Du, L.; Xu, X.; Kearns, W.G. Two Different Microarray Technologies for Preimplantation Genetic Diagnosis and Screening, Due to Reciprocal Translocation Imbalances, Demonstrate Equivalent Euploidy and Clinical Pregnancy Rates. J. Assist. Reprod. Genet. 2014, 31, 843–850. [Google Scholar] [CrossRef]
- Xiong, B.; Tan, K.; Tan, Y.Q.; Gong, F.; Zhang, S.P.; Lu, C.F.; Luo, K.L.; Lu, G.X.; Lin, G. Using SNP Array to Identify Aneuploidy and Segmental Imbalance in Translocation Carriers. Genom. Data 2014, 2, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Idowu, D.; Merrion, K.; Wemmer, N.; Mash, J.G.; Pettersen, B.; Kijacic, D.; Lathi, R.B. Pregnancy Outcomes Following 24-Chromosome Preimplantation Genetic Diagnosis in Couples with Balanced Reciprocal or Robertsonian Translocations. Fertil. Steril. 2015, 103, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, C.; Wang, J.; Zeng, Y.; Zhou, W.; Li, R.; Zhou, C.; Deng, M.-F.; Xu, Y. Number of Blastocysts Biopsied as a Predictive Indicator to Obtain at Least One Normal/Balanced Embryo Following Preimplantation Genetic Diagnosis with Single Nucleotide Polymorphism Microarray in Translocation Cases. J. Assist. Reprod. Genet. 2017, 34, 51–59. [Google Scholar] [CrossRef][Green Version]
- Zhang, S.; Zhao, D.; Zhang, J.; Mao, Y.; Kong, L.; Zhang, Y.; Liang, B.; Sun, X.; Xu, C. BasePhasing: A highly efficient approach for preimplantation genetic haplotyping in clinical application of balanced translocation carriers. BMC Med. Genom. 2019, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- Beyer, C.E.; Willats, E. Natural Selection between Day 3 and Day 5/6 PGD Embryos in Couples with Reciprocal or Robertsonian Translocations. J. Assist. Reprod. Genet. 2017, 34, 1483–1492. [Google Scholar] [CrossRef]
- Wells, D.; Kaur, K.; Grifo, J.; Glassner, M.; Taylor, J.C.; Fragouli, E.; Munne, S. Clinical Utilisation of a Rapid Low-Pass Whole Genome Sequencing Technique for the Diagnosis of Aneuploidy in Human Embryos Prior to Implantation. J. Med. Genet. 2014, 51, 553–562. [Google Scholar] [CrossRef]
- Maxwell, S.M.; Colls, P.; Hodes-Wertz, B.; McCulloh, D.H.; McCaffrey, C.; Wells, D.; Munné, S.; Grifo, J.A. Why Do Euploid Embryos Miscarry? A Case-Control Study Comparing the Rate of Aneuploidy within Presumed Euploid Embryos That Resulted in Miscarriage or Live Birth Using next-Generation Sequencing. Fertil. Steril. 2016, 106, 1414–1419.e5. [Google Scholar] [CrossRef]
- Biricik, A.; Cotroneo, E.; Minasi, M.G.; Greco, P.F.; Bono, S.; Surdo, M.; Lecciso, F.; Sessa, M.; Fiorentino, F.; Spinella, F.; et al. Cross-Validation of Next-Generation Sequencing Technologies for Diagnosis of Chromosomal Mosaicism and Segmental Aneuploidies in Preimplantation Embryos Model. Life 2021, 11, 340. [Google Scholar] [CrossRef]
- Cuman, C.; Beyer, C.E.; Brodie, D.; Fullston, T.; Lin, J.I.; Willats, E.; Zander-Fox, D.; Mullen, J. Defining the Limits of Detection for Chromosome Rearrangements in the Preimplantation Embryo Using next Generation Sequencing. Hum. Reprod. 2018, 33, 1566–1576. [Google Scholar] [CrossRef]
- Yin, X.; Tan, K.; Vajta, G.; Jiang, H.; Tan, Y.; Zhang, C.; Chen, F.; Chen, S.; Zhang, C.; Pan, X.; et al. Massively Parallel Sequencing for Chromosomal Abnormality Testing in Trophectoderm Cells of Human Blastocysts. Biol. Reprod. 2013, 88, 69. [Google Scholar] [CrossRef]
- Tan, Y.; Yin, X.; Zhang, S.; Jiang, H.; Tan, K.; Li, J.; Xiong, B.; Gong, F.; Zhang, C.; Pan, X.; et al. Clinical Outcome of Preimplantation Genetic Diagnosis and Screening Using next Generation Sequencing. Gigascience 2014, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Gui, B.; Yao, Z.; Li, Y.; Liu, D.; Liu, N.; Xia, Y.; Huang, Y.; Mei, L.; Ma, R.; Lu, S.; et al. Chromosomal Analysis of Blastocysts from Balanced Chromosomal Rearrangement Carriers. Reproduction 2016, 151, 455–464. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chow, J.F.C.; Yeung, W.S.B.; Lee, V.C.Y.; Lau, E.Y.L.; Ng, E.H.Y. Evaluation of Preimplantation Genetic Testing for Chromosomal Structural Rearrangement by a Commonly Used next Generation Sequencing Workflow. Eur. J. Obs. Gynecol. Reprod. Biol. 2018, 224, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shen, J.; Cram, D.S.; Ma, M.; Wang, H.; Zhang, W.; Fan, J.; Gao, Z.; Zhang, L.; Li, Z.; et al. Preferential Selection and Transfer of Euploid Noncarrier Embryos in Preimplantation Genetic Diagnosis Cycles for Reciprocal Translocations. Fertil. Steril. 2017, 108, 620–627.e4. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Ammae, M.; Satoh, M.; Mizuno, S.; Nakaoka, Y.; Morimoto, Y. Analysis of Clinical Outcomes and Meiotic Segregation Modes Following Preimplantation Genetic Testing for Structural Rearrangements Using aCGH/NGS in Couples with Balanced Chromosome Rearrangement. Reprod Med. Biol. 2022, 21, e12476. [Google Scholar] [CrossRef]
- Chen, S.; Yin, X.; Zhang, S.; Xia, J.; Liu, P.; Xie, P.; Yan, H.; Liang, X.; Zhang, J.; Chen, Y.; et al. Comprehensive Preimplantation Genetic Testing by Massively Parallel Sequencing. Hum. Reprod. 2021, 36, 236–247. [Google Scholar] [CrossRef]
- Boynukalin, F.K.; Gultomruk, M.; Turgut, N.E.; Rubio, C.; Rodrigo, L.; Yarkiner, Z.; Ecemis, S.; Karlikaya, G.; Findikli, N.; Bahceci, M. The Impact of Patient, Embryo, and Translocation Characteristics on the Ploidy Status of Young Couples Undergoing Preimplantation Genetic Testing for Structural Rearrangements (PGT-SR) by next Generation Sequencing (NGS). J Assist Reprod. Genet. 2021, 38, 387–396. [Google Scholar] [CrossRef]
- Walters-Sen, L.; Neitzel, D.; Wilkinson, J.; Poll, S.; Faulkner, N.; Aradhya, S. EP472: Deriving Risk Estimates for Balanced Rearrangement Carriers Utilizing PGT-SR Data. Genet. Med. 2022, 24, S299–S300. [Google Scholar] [CrossRef]
- Yuan, P.; Zheng, L.; Ou, S.; Zhao, H.; Li, R.; Luo, H.; Tan, X.; Zhang, Q.; Wang, W. Evaluation of Chromosomal Abnormalities from Preimplantation Genetic Testing to the Reproductive Outcomes: A Comparison between Three Different Structural Rearrangements Based on next-Generation Sequencing. J. Assist. Reprod. Genet. 2021, 38, 709–718. [Google Scholar] [CrossRef]
- Tong, J.; Jiang, J.; Niu, Y.; Zhang, T. Do Chromosomal Inversion Carriers Really Need Preimplantation Genetic Testing? J. Assist. Reprod. Genet. 2022, 39, 2573–2579. [Google Scholar] [CrossRef]
- Zheng, W.; Ren, B.; Mu, M.; Liu, Y.; Liu, X.; Yang, C.; Yang, S.; Yang, R.; Li, J.; Zu, R.; et al. Perinatal Outcomes of Singleton Live Births Following Preimplantation Genetic Testing for Chromosomal Structural Rearrangements in Single Frozen-Thawed Blastocyst Transfer Cycles: A Retrospective Cohort Study. Reprod. Sci. 2022, 29, 3039–3046. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.A.; Backenroth, D.; Hakam-Spector, E.; Renbaum, P.; Mann, T.; Zahdeh, F.; Segel, R.; Zeligson, S.; Eldar-Geva, T.; Ben-Ami, I.; et al. Expanded clinical validation of Haploseek for comprehensive preimplantation genetic testing. Genet. Med. Off. J. Am. Coll. Med. Genet. 2021, 23, 1334–1340. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ullmann, R.; Langnick, C.; Menzel, C.; Wotschofsky, Z.; Hu, H.; Döring, A.; Hu, Y.; Kang, H.; Tzschach, A.; et al. Breakpoint Analysis of Balanced Chromosome Rearrangements by Next-Generation Paired-End Sequencing. Eur. J. Hum. Genet. 2010, 18, 539–543. [Google Scholar] [CrossRef]
- Zhai, F.; Wang, Y.; Li, H.; Wang, Y.; Zhu, X.; Kuo, Y.; Guan, S.; Li, J.; Song, S.; He, Q.; et al. Preimplantation Genetic Testing for Structural Rearrangement Based on Low-Coverage next-Generation Sequencing Accurately Discriminates between Normal and Carrier Embryos for Patients with Translocations. Reprod. BioMed. Online 2022, 45, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Yin, B.; Zhu, Y.; Li, G.; Ye, L.; Liang, D.; Zeng, Y. First Report on an X-Linked Hypohidrotic Ectodermal Dysplasia Family with X Chromosome Inversion: Breakpoint Mapping Reveals the Pathogenic Mechanism and Preimplantation Genetics Diagnosis Achieves an Unaffected Birth. Clin. Chim. Acta 2017, 475, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Findikli, N.; Kahraman, S.; Kumtepe, Y.; Donmez, E.; Benkhalifa, M.; Biricik, A.; Sertyel, S.; Berkil, H.; Oncu, N. Assessment of DNA Fragmentation and Aneuploidy on Poor Quality Human Embryos. Reprod. Biomed. Online 2004, 8, 196–206. [Google Scholar] [CrossRef]
- Evsikov, S.; Cieslak, J.; Verlinsky, Y. Effect of Chromosomal Translocations on the Development of Preimplantation Human Embryos in Vitro. Fertil. Steril. 2000, 74, 672–677. [Google Scholar] [CrossRef]
- Chang, E.M.; Han, J.E.; Kwak, I.P.; Lee, W.S.; Yoon, T.K.; Shim, S.H. Preimplantation Genetic Diagnosis for Couples with a Robertsonian Translocation: Practical Information for Genetic Counseling. J. Assist. Reprod. Genet. 2012, 29, 67–75. [Google Scholar] [CrossRef]
- Findikli, N.; Kahraman, S.; Kumtepe, Y.; Donmez, E.; Biricik, A.; Sertyel, S.; Berkil, H.; Melil, S. Embryo Development Characteristics in Robertsonian and Reciprocal Translocations: A Comparison of Results with Non-Translocation Cases. Reprod. BioMed. Online 2003, 7, 563–571. [Google Scholar] [CrossRef]
- Campbell, A.; Fishel, S.; Bowman, N.; Duffy, S.; Sedler, M.; Hickman, C.F.L. Modelling a Risk Classification of Aneuploidy in Human Embryos Using Non-Invasive Morphokinetics. Reprod. BioMed. Online 2013, 26, 477–485. [Google Scholar] [CrossRef]
- Meseguer, M.; Herrero, J.; Tejera, A.; Hilligsoe, K.M.; Ramsing, N.B.; Remohi, J. The Use of Morphokinetics as a Predictor of Embryo Implantation. Hum. Reprod. 2011, 26, 2658–2671. [Google Scholar] [CrossRef] [PubMed]
- Amir, H.; Barbash-Hazan, S.; Kalma, Y.; Frumkin, T.; Malcov, M.; Samara, N.; Hasson, J.; Reches, A.; Azem, F.; Ben-Yosef, D. Time-Lapse Imaging Reveals Delayed Development of Embryos Carrying Unbalanced Chromosomal Translocations. J. Assist. Reprod. Genet. 2019, 36, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lammers, J.; Reignier, A.; Splingart, C.; Moradkhani, K.; Barrière, P.; Fréour, T. Morphokinetic Parameters in Chromosomal Translocation Carriers Undergoing Preimplantation Genetic Testing. Reprod. BioMed. Online 2019, 38, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Insogna, I.G.; Lanes, A.; Dobson, L.; Ginsburg, E.S.; Racowsky, C.; Yanushpolsky, E. Blastocyst. Conversion Rate and Ploidy in Patients with Structural Rearrangements. J. Assist. Reprod. Genet. 2021, 38, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Escudero, T.; Cekleniak, N.A.; Sable, D.B.; Garrisi, M.G.; Munne, S. Patterns of Ovarian Response to Gonadotropin Stimulation in Female Carriers of Balanced Translocation. Fertil. Steril. 2005, 83, 1504–1509. [Google Scholar] [CrossRef]
- Dechanet, C.; Castelli, C.; Reyftmann, L.; Hamamah, S.; Hedon, B.; Dechaud, H.; Anahory, T. Do Female Translocations Influence the Ovarian Response Pattern to Controlled Ovarian Stimulation in Preimplantation Genetic Diagnosis? Hum. Reprod. 2011, 26, 1232–1240. [Google Scholar] [CrossRef]
- Lledo, B.; Ortiz, J.A.; Morales, R.; Ten, J.; de la Fuente, P.E.; Garcia-Ochoa, C.; Bernabeu, R. The Paternal Effect of Chromosome Translocation Carriers Observed from Meiotic Segregation in Embryos. Hum. Reprod. 2010, 25, 1843–1848. [Google Scholar] [CrossRef]
- Ye, Y.; Qian, Y.; Xu, C.; Jin, F. Meiotic Segregation Analysis of Embryos from Reciprocal Translocation Carriers in PGD Cycles. Reprod. BioMed. Online 2012, 24, 83–90. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, W.; Zhu, Y.; Chen, H.; Yan, J.; Chen, Z.-J. Effects of a Carrier’s Sex and Age on the Segregation Patterns of the Trivalent of Robertsonian Translocations. J. Assist. Reprod. Genet. 2019, 36, 1963–1969. [Google Scholar] [CrossRef]
- Song, H.; Shi, H.; Yang, E.-T.; Bu, Z.-Q.; Jin, Z.-Q.; Huo, M.-Z.; Zhang, Y. Effects of Gender of Reciprocal Chromosomal Translocation on Blastocyst Formation and Pregnancy Outcome in Preimplantation Genetic Testing. Front. Endocrinol. 2021, 12, 704299. [Google Scholar] [CrossRef]
- LeMaire-Adkins, R.; Radke, K.; Hunt, P.A. Lack of Checkpoint Control at the Metaphase/Anaphase Transition: A Mechanism of Meiotic Nondisjunction in Mammalian Females. J. Cell Biol. 1997, 139, 1611–1619. [Google Scholar] [CrossRef]
- Xie, P.; Hu, L.; Peng, Y.; Tan, Y.-Q.; Luo, K.; Gong, F.; Lu, G.; Lin, G. Risk Factors Affecting Alternate Segregation in Blastocysts From Preimplantation Genetic Testing Cycles of Autosomal Reciprocal Translocations. Front. Genet. 2022, 13, 880208. [Google Scholar] [CrossRef] [PubMed]
- Benkhalifa, M.; Kasakyan, S.; Clement, P.; Baldi, M.; Tachdjian, G.; Demirol, A.; Gurgan, T.; Fiorentino, F.; Mohammed, M.; Qumsiyeh, M.B. Array comparative genomic hybridization profiling of first-trimester spontaneous abortions that fail to grow in vitro. Prenat. Diagn. 2005, 25, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Franasiak, J.M.; Forman, E.J.; Hong, K.H.; Werner, M.D.; Upham, K.M.; Treff, N.R.; Scott, R.T., Jr. The nature of aneuploidy with increasing age of the female partner: A review of 15,169 consecutive trophectoderm biopsies evaluated with comprehensive chromosomal screening. Fertil. Steril. 2014, 101, 656–663.e1. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Diez, C.; FitzHarris, G. Causes and consequences of chromosome segregation error in preimplantation embryos. Reproduction 2018, 155, R63–R76. [Google Scholar] [CrossRef]
- Taylor, T.H.; Gitlin, S.A.; Patrick, J.L.; Crain, J.L.; Wilson, J.M.; Griffin, D.K. The origin, mechanisms, incidence and clinical consequences of chromosomal mosaicism in humans. Hum. Reprod. Update 2014, 20, 571–581. [Google Scholar] [CrossRef]
- Coonen, E.; Derhaag, J.G.; Dumoulin, J.C.; van Wissen, L.C.; Bras, M.; Janssen, M.; Evers, J.L.; Geraedts, J.P. Anaphase lagging mainly explains chromosomal mosaicism in human preimplantation embryos. Hum. Reprod. 2004, 19, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Lindenbaum, R.H.; Hultén, M.; McDermott, A.; Seabright, M. The Prevalence of Translocations in Parents of Children with Regular Trisomy 21: A Possible Interchromosomal Effect? J. Med. Genet. 1985, 22, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.; Vidal, F.; Benet, J.; Templado, C.; Marina, S.; Egozcue, J. XY-Trivalent Association and Synaptic Anomalies in a Male Carrier of a Robertsonian t(13;14) Translocation. Hum. Reprod. 1991, 6, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.H.; Spriggs, E.L. Sperm Chromosome Complements in a Man Heterozygous for a Reciprocal Translocation 46,XY,t(9;13)(Q21.1;Q21.2) and a Review of the Literature. Clin. Genet. 1995, 47, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Amiel, A.; Sardos-Albertini, F.; Fejgin, M.D.; Sharony, R.; Diukman, R.; Bartoov, B. Interchromosomal Effect Leading to an Increase in Aneuploidy in Sperm Nuclei in a Man Heterozygous for Pericentric Inversion (Inv 9) and C-Heterochromatin. J. Hum. Genet. 2001, 46, 245–250. [Google Scholar] [CrossRef]
- Machev, N.; Gosset, P.; Warter, S.; Treger, M.; Schillinger, M.; Viville, S. Fluorescence in Situ Hybridization Sperm Analysis of Six Translocation Carriers Provides Evidence of an Interchromosomal Effect. Fertil. Steril. 2005, 84, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Godo, A.; Blanco, J.; Vidal, F.; Anton, E. Accumulation of Numerical and Structural Chromosome Imbalances in Spermatozoa from Reciprocal Translocation Carriers. Hum. Reprod. 2013, 28, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Scriven, P.N.; Flinter, F.A.; Braude, P.R.; Ogilvie, C.M. Robertsonian Translocations--Reproductive Risks and Indications for Preimplantation Genetic Diagnosis. Hum. Reprod. 2001, 16, 2267–2273. [Google Scholar] [CrossRef] [PubMed]
- Ferfouri, F.; Boitrelle, F.; Clément, P.; Molina Gomes, D.; Selva, J.; Vialard, F. Can One Translocation Impact the Meiotic Segregation of Another Translocation? A Sperm-FISH Analysis of a 46,XY,t(1;16)(Q21;P11.2),t(8;9) (Q24.3;P24) Patient and His 46,XY,t(8;9)(Q24.3;P24) Brother and Cousin. MHR: Basic Sci. Reprod. Med. 2013, 19, 109–117. [Google Scholar] [CrossRef][Green Version]
- Young, D.; Klepacka, D.; McGarvey, M.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Infertility Patients with Chromosome Inversions Are Not Susceptible to an Inter-Chromosomal Effect. J. Assist. Reprod. Genet. 2019, 36, 509–516. [Google Scholar] [CrossRef]
- Vanneste, E.; Voet, T.; Melotte, C.; Debrock, S.; Sermon, K.; Staessen, C.; Liebaers, I.; Fryns, J.-P.; D’Hooghe, T.; Vermeesch, J.R. What next for Preimplantation Genetic Screening? High Mitotic Chromosome Instability Rate Provides the Biological Basis for the Low Success Rate. Hum. Reprod. 2009, 24, 2679–2682. [Google Scholar] [CrossRef]
- Alfarawati, S.; Fragouli, E.; Colls, P.; Wells, D. Embryos of Robertsonian Translocation Carriers Exhibit a Mitotic Interchromosomal Effect That Enhances Genetic Instability during Early Development. PLoS Genet. 2012, 8, e1003025. [Google Scholar] [CrossRef]
- Lynch, C.A.; Gordon, T.; Griffin, D.K. Retrospective analysis of 479 Pgt-Sr cycles—Analysis of chromosome information and availability of embryos for transfer. Fertil. Steril. 2021, 116, e397. [Google Scholar] [CrossRef]
- Lynch, C.A.; Whiting, O.; Sloan, J.; Cameron, E.; Xanthopoulou, L.; Gordon, T.; Griffin, D.K. The importance of retrospective data analysis in genetic counselling for Pgt-sr—maternal age is more significant than rearrangement type. Fertil. Steril. 2022, 118, e97. [Google Scholar] [CrossRef]
- Ogur, C.; Kahraman, S.; Griffin, D.K.; Cinar, C.; Tufekci, M.A.; Cetinkaya, M.; Temel, S.G.; Yilmaz, A. Preimplantation Genetic Testing for Structural Rearrangements (PGT-SR) in 300 Couples Reveals Individual Specific Risk Factors but an Inter Chromosomal Effect (ICE) Is Unlikely. Reprod. BioMed. Online 2022, S1472648322005223. [Google Scholar] [CrossRef]
- Scriven, P.N. PGT-SR: The Red-Herring and the Siren; Interchromosomal Effect and Screening for Unrelated Aneuploidy. J. Assist. Reprod. Genet. 2021, 38, 1015–1018. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Griffin, D.K.; Ogur, C. PGT-SR: A Comprehensive Overview and a Requiem for the Interchromosomal Effect. DNA 2023, 3, 41-64. https://doi.org/10.3390/dna3010004
Griffin DK, Ogur C. PGT-SR: A Comprehensive Overview and a Requiem for the Interchromosomal Effect. DNA. 2023; 3(1):41-64. https://doi.org/10.3390/dna3010004
Chicago/Turabian StyleGriffin, Darren K., and Cagri Ogur. 2023. "PGT-SR: A Comprehensive Overview and a Requiem for the Interchromosomal Effect" DNA 3, no. 1: 41-64. https://doi.org/10.3390/dna3010004
APA StyleGriffin, D. K., & Ogur, C. (2023). PGT-SR: A Comprehensive Overview and a Requiem for the Interchromosomal Effect. DNA, 3(1), 41-64. https://doi.org/10.3390/dna3010004