
Nanotechnology-Enabled COVID-19 mRNA Vaccines



Definition
:1. Introduction
2. Mechanism of Action for COVID-19 mRNA Vaccines
3. Efficacy and Safety of COVID-19 mRNA Vaccines
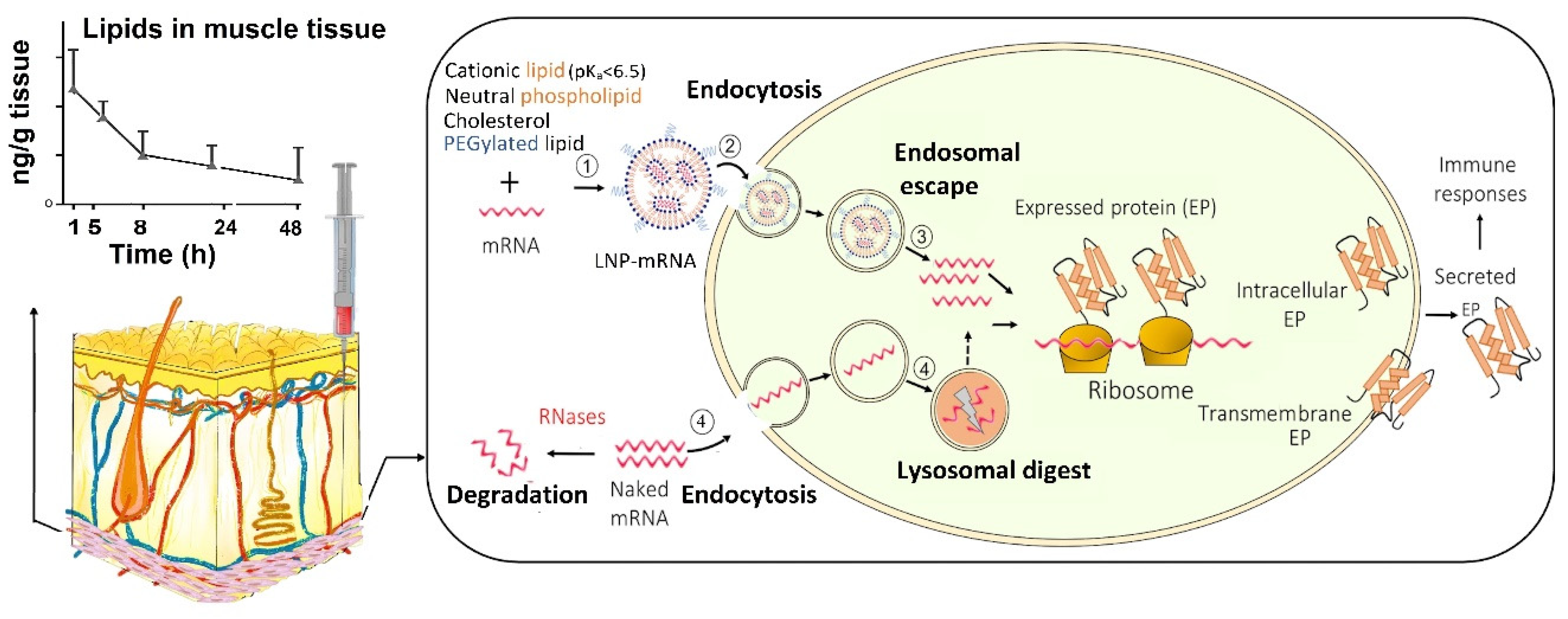
4. Mechanisms for Nanoparticle-Mediated Intracellular Delivery
4.1. The Need of Innovative Approach for Intracellular Delivery of mRNA
4.2. Formulation Compositions of Covid-19 mRNA Vaccines
4.3. How Does LNPs Assist mRNA Delivery into the Ribosomes Inside Cells
5. Manufacturing and Stability
5.1. Manufacturing Process
5.2. Stability and Storage
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Entry Link on the Encyclopedia Platform
Abbreviations
| ACE2 | Angiotensin converting enzyme II |
| APCs | Antigen presenting cells |
| CDC | Centers for Disease Control and Prevention |
| COVID-19 | Coronavirus disease-2019 |
| DSPC | 1,2-Distearoyl-sn-glycero-3-phosphocholine |
| EUA | Emergency Use Authorisation |
| FDA | Food and Drug Administration |
| LNPs | Lipid nanoparticles |
| MHC | Major histocompatibility complex |
| mRNA | Messenger ribonucleic acid |
| PEG | Poly(ethylene glycol) |
| RNases | Ribonucleases |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| WHO | World Health Organisation |
References
- Zhou, P.; Yang, X.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N. BNT162b2 mRNA COVID-19 vaccine: First approval. Drugs 2021, 81, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. FDA Briefing Document: Moderna COVID-19 Vaccine. Available online: https://www.fda.gov/media/144434/download (accessed on 3 May 2021).
- European Medicines Agency. Comirnaty: EPAR-Product Information. Available online: https://www.ema.europa.eu/en/documents/product-information/comirnaty-epar-product-information_en.pdf (accessed on 3 May 2021).
- Wu, Z.; Li, T. Nanoparticle-mediated cytoplasmic delivery of messenger RNA vaccines: Challenges and future perspectives. Pharm. Res. 2021, 38, 473–478. [Google Scholar] [CrossRef]
- Brenner, S.; Jacob, F.; Meselson, M. An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature 1961, 190, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines-a new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Control. Release 2021, 333, 511–520. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, J.; Xiao, T.; Peng, H.; Sterling, S..; Walsh, R., Jr.; Rawson, S.; Rits-Volloch, S.; Chen, B. Distinct conformational states of SARS-CoV-2 spike protein. Science 2020, 369, 1586–1592. [Google Scholar] [CrossRef]
- Zeng, C.; Zhang, C.; Walker, P.G.; Dong, Y. Formulation and delivery technologies for mRNA vaccines. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–40. [Google Scholar]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and challenges in the delivery of mRNA-based vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef]
- Blakney, A.K.; Ip, S.; Geall, A.J. An update on self-amplifying mRNA vaccine development. Vaccines 2021, 9, 97. [Google Scholar] [CrossRef]
- Kutzler, M.A.; Weiner, D.B. DNA vaccines: Ready for prime time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Polack, F.; Thomas, S.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Banerji, A.; Wickner, P.G.; Saff, R.; Stone, C.A.; Robinson, L.B.; Long, A.A.; Wolfson, A.R.; Williams, P.; Khan, D.A.; Phillips, E.; et al. mRNA vaccines to prevent COVID-19 disease and reported allergic reactions: Current evidence and suggested approach. J. Allergy Clin. Immunol. Pract. 2021, 9, 1423–1437. [Google Scholar] [CrossRef]
- Greenhawt, M.; Abrams, E.M.; Oppenheimer, J.; Leek, T.K.V.; Mack, D.P.; Singer, A.G.; Shaker, M. The COVID-19 pandemic in 2021: Avoiding overdiagnosis of anaphylaxis risk while safely vaccinating the world. J. Allergy Clin. Immunol. Pract. 2021, 9, 1438. [Google Scholar] [CrossRef] [PubMed]
- Reichumuth, A.M.; Oberli, M.; Jaklenec, A.; Langer, R.B.D. mRNA vaccine delivery using lipid nanoparticles. Ther. Deliv. 2016, 7, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Fotin-Mleczek, M.; Roth, G.; Becker, C.; Dam, T.C.; Verdurmen, W.P.R.; Brock, R.; Probst, J.; Schlake, T. Protein expression from exogenous mRNA: Uptake by receptor-mediated endocytosis and trafficking via the lysosomal pathway. RNA Biol. 2011, 8. [Google Scholar] [CrossRef]
- Aldosari, B.N.; Alfagih, I.M.; Almurshedi, A.S.; Hinrichs, J. Lipid nanoparticles as delivery systems for RNA-based vaccines. Pharmaceutics 2021, 13, 206. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Dong, Y. Nanoscale platforms for messenger RNA delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2018, 11, 1530. [Google Scholar] [CrossRef]
- Hassett, K.J.; Benenato, K.E.; Jacquinet, E.; Lee, A.; Woods, A.; Yuzhakov, O.; Himansu, S.; Deterling, J.; Geilich, B.M.; Ketova, T.; et al. Optimization of lipid nanoparticles for intramuscular administration of mRNA vaccines. Mol. Ther. Nucleic Acids 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Crommelin, D.J.A.; Anchordoquy, T.J.; Volkin, D.B.; Jiskoot, W.; Mastrobattista, E. Addressing the cold reality of mRNA vaccine stability. J. Pharm. Sci. 2021, 110, 997–1001. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Vaccine Name | Developer (s) | Formulation [5] | Stage of Development (Timeline) |
|---|---|---|---|
| CVnCoV mRNA | CureVac, Germany | LNP-mRNA | Phase IIb/III NCT 04652102 (December 2020–May 2022) |
| ARCT-021 | Arcturus Therapeutics/Duke-NUS Medical School, USA and Singapore | LUNAR® (pH-sensitive LNP-mediated delivery of saRNA) | Phase II NCT04668339 (January 2021–April 2022) NCT04728347 (January 2021–June 2022) |
| LNP-nCoVsaRNA-02 | Imperial College London, UK | LNP-saRNA(proprietary, cationic, PEGylated) | Phase I NCT04934111 (September 2021–August 2022) |
| ARCoV | PLAAMS */ Walvax Biotech, China | LNP-mRNA | Phase Ib ChiCTR2000034112 (June 2020–December 2021) ChiCTR2000039212 (October 2020–December 2021) |
| Lipid | BNT162b2 | mRNA-1273 |
|---|---|---|
| Ionisable cationic lipid (A) | ALC-0315 | SM-102 |
| PEGylated lipid (B) | 2-[(polyethylene glycol)-2000]-N,N-ditetradecylacetamide (ALC-0159) | 1,2-dimyristoyl-rac-glycero3-methoxypolyethylene glycol-2000 |
| Neutral lipid (C) | DSPC | DSPC |
| A:B:C:cholesterol molar ratios | 46.3:1.6:9.4:42.7 | 50:1.5:10:38.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Yang, K.; Shelling, A.N.; Wu, Z. Nanotechnology-Enabled COVID-19 mRNA Vaccines. Encyclopedia 2021, 1, 773-780. https://doi.org/10.3390/encyclopedia1030059
Gao Y, Yang K, Shelling AN, Wu Z. Nanotechnology-Enabled COVID-19 mRNA Vaccines. Encyclopedia. 2021; 1(3):773-780. https://doi.org/10.3390/encyclopedia1030059
Chicago/Turabian StyleGao, Yu, Kaiyun Yang, Andrew N. Shelling, and Zimei Wu. 2021. "Nanotechnology-Enabled COVID-19 mRNA Vaccines" Encyclopedia 1, no. 3: 773-780. https://doi.org/10.3390/encyclopedia1030059
APA StyleGao, Y., Yang, K., Shelling, A. N., & Wu, Z. (2021). Nanotechnology-Enabled COVID-19 mRNA Vaccines. Encyclopedia, 1(3), 773-780. https://doi.org/10.3390/encyclopedia1030059