
Carbon Metabolic Profiling as a Response to Previous Plant Mycotrophy and Soil Disturbance in Wheat Growth


Abstract
1. Introduction
2. Materials and Methods
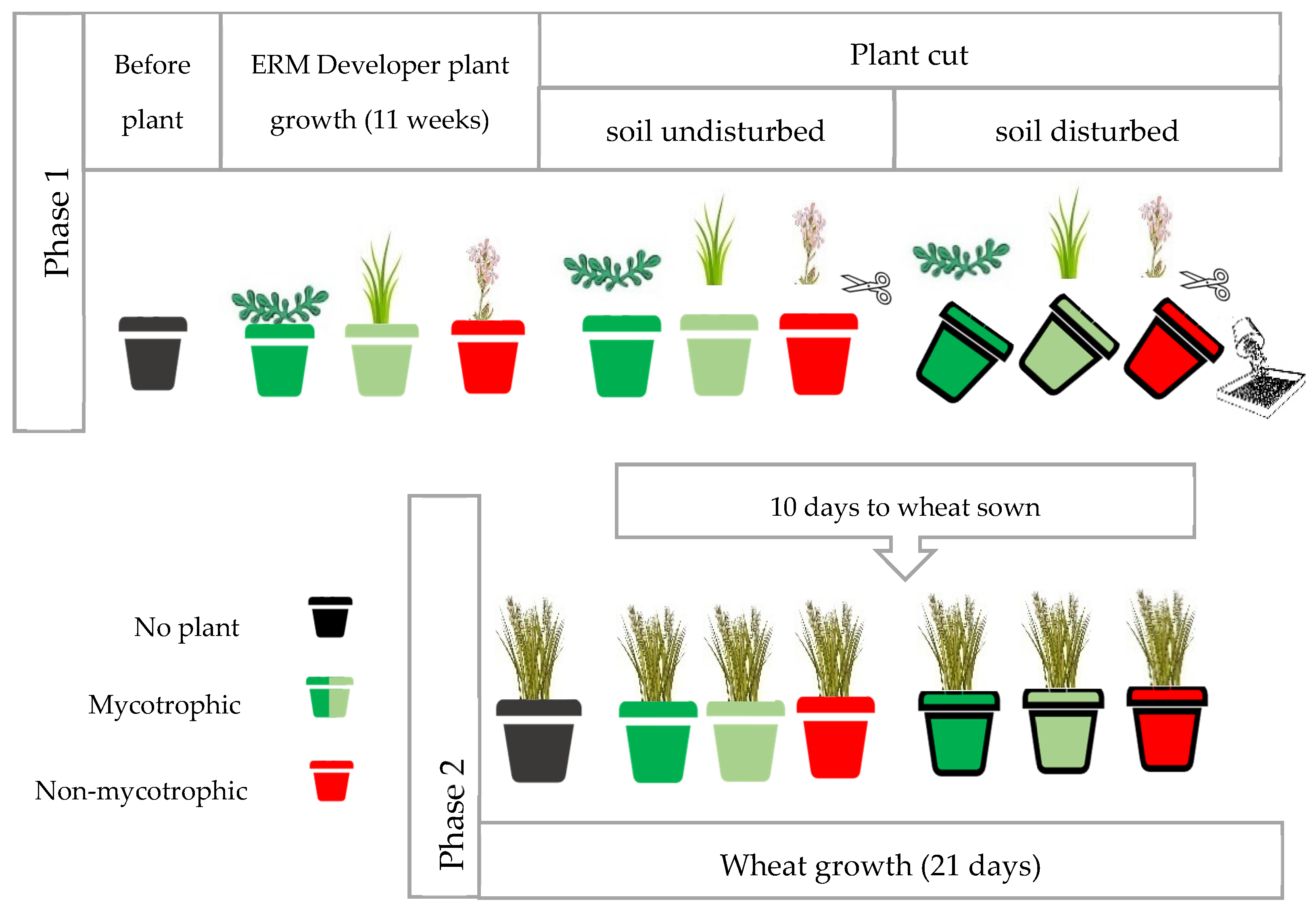
2.1. Experimental Design
2.2. Bacterial Count Estimation
2.3. BIOLOGTM Ecoplates
2.4. Statistical Analysis
3. Results
3.1. Metabolic Differences in Phase 1
3.2. Metabolic Differences in Phase 2
3.3. Overall Metabolic Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Insam, H.; Goberna, M. Section 4 update: Use of Biolog® for the Community Level Physiological Profiling (CLPP) of environmental samples. In Molecular Microbial Ecology Manual; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2008; pp. 2755–2762. [Google Scholar] [CrossRef]
- Gałązka, A.; Gawryjołek, K.; Grządziel, J.; Frąc, M.; Księżak, J. Microbial community diversity and the interaction of soil under maize growth in different cultivation techniques. Plant Soil Environ. 2017, 63, 264–270. [Google Scholar] [CrossRef]
- Adams, T.C.; Brye, K.R.; Savin, M.C.; Lee, J.A.; Gbur, E.E. Microbial carbon substrate utilization differences among high- and average-yield soybean areas. Agriculture 2017, 7, 48. [Google Scholar] [CrossRef]
- Lan, X.; Du, H.; Peng, W.; Liu, Y.; Fang, Z.; Song, T. Functional diversity of the soil culturable microbial community in eucalyptus plantations of different ages in Guangxi, South China. Forests 2019, 10, 1083. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Root Exudates and Their Molecular Interactions with Rhizospheric Microbes. In Plant, Soil and Microbes; Springer International Publishing: Cham, Switzerland, 2016; pp. 59–77. [Google Scholar] [CrossRef]
- Jansa, J.; Bukovská, P.; Gryndler, M. Mycorrhizal hyphae as ecological niche for highly specialized hypersymbionts—Or just soil free-riders? Front. Plant Sci. 2013, 4, 134. [Google Scholar] [CrossRef]
- Toljander, J.F.; Lindahl, B.D.; Paul, L.R.; Elfstrand, M.; Finlay, R.D. Influence of arbuscular mycorrhizal mycelial exudates on soil bacterial growth and community structure. FEMS Microbiol. Ecol. 2007, 61, 295–304. [Google Scholar] [CrossRef]
- Goss, M.J.; Carvalho, M.; Brito, I. Challenges to Agriculture Systems. In Functional Diversity of Mycorrhiza and Sustainable Agriculture—Management to Overcome Biotic and Abiotic Stresses; Academic Press and Elsevier: London, UK, 2017; Chapter 1; p. 231. [Google Scholar]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef]
- Brito, I.; Goss, M.J.; Alho, L.; Brígido, C.; van Tuinen, D.; Félix, M.R.; Carvalho, M. Agronomic management of AMF functional diversity to overcome biotic and abiotic stresses. The role of plant sequence and intact extraradical mycelium. Fungal Ecol. 2019, 40, 72–81. [Google Scholar] [CrossRef]
- Carvalho, M.; Brito, I.; Alho, L.; Goss, M.J. Assessing the progress of colonization by arbuscular mycorrhiza of four plant species under different temperature regimes. J. Plant Nutr. Soil Sci. 2015, 178, 515–522. [Google Scholar] [CrossRef]
- Goss, M.J.; Carvalho, M.; Brito, I. The Significance of an Intact Extraradical Mycelium and Early Root Colonization in Managing Arbuscular Mycorrhizal Fungi. In Functional Diversity of Mycorrhiza and Sustainable Agriculture; Goss, M.J., De Carvalho, M., Brito, I., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Chapter 6; pp. 112–131. [Google Scholar]
- Rachwał, K.; Gustaw, K.; Kazimierczak, W.; Waśko, A. Is soil management system really important? Comparison of microbial community diversity and structure in soils managed under organic and conventional regimes with some view on soil properties. PLoS ONE 2021, 16, e0256969. [Google Scholar] [CrossRef]
- Smith, C.R.; Blair, P.L.; Boyd, C.; Cody, B.; Hazel, A.; Hedrick, A.; Kathuria, H.; Khurana, P.; Kramer, B.; Muterspaw, K.; et al. Microbial community responses to soil tillage and crop rotation in a corn/soybean agroecosystem. Ecol. Evol. 2016, 6, 8075–8084. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.J.; Lin, S.; Liu, T.Q.; Cao, C.G.; Li, C.F. Effects of conservation tillage on topsoil microbial metabolic characteristics and organic carbon within aggregates under a rice (Oryza sativa L.)—Wheat (Triticum aestivum L.) cropping system in Central China. PLoS ONE 2016, 11, e0146145. [Google Scholar] [CrossRef] [PubMed]
- Hartman, K.; van der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.C.; Schlaeppi, K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 14. [Google Scholar] [CrossRef]
- Wipf, H.M.L.; Xu, L.; Gao, C.; Spinner, H.B.; Taylor, J.; Lemaux, P.; Mitchell, J.; Coleman-Derr, D. Agricultural Soil Management Practices Differentially Shape the Bacterial and Fungal Microbiomes of Sorghum bicolor. Appl. Environ. Microbiol. 2021, 87, e02345-20. [Google Scholar] [CrossRef]
- Gryta, A.; Frac, M.; Oszust, K. Genetic and metabolic diversity of soil microbiome in response to exogenous organic matter amendments. Agronomy 2020, 10, 546. [Google Scholar] [CrossRef]
- Lladó, S.; Baldrian, P. Community-level physiological profiling analyses show potential to identify the copiotrophic bacteria present in soil environments. PLoS ONE 2017, 12, e0171638. [Google Scholar] [CrossRef]
- Gajda, A.M.; Czyż, E.A.; Furtak, K.; Jończyk, K. Effects of crop production practices on soil characteristics and metabolic diversity of microbial communities under winter wheat. Soil Res. 2019, 57, 124–131. [Google Scholar] [CrossRef]
- Goss, M.J.; Carvalho, M. Manganese toxicity: The significance of magnesium for the sensitivity of wheat plants. Plant Soil 1992, 139, 91–98. [Google Scholar] [CrossRef]
- Brígido, C.; van Tuinen, D.; Brito, I.; Alho, L.; Goss, M.J.; Carvalho, M. Management of the biological diversity of AM fungi by combination of host plant succession and integrity of extraradical mycelium. Soil Biol. Biochem. 2017, 112, 237–247. [Google Scholar] [CrossRef]
- Brito, I.; Carvalho, M.; Alho, L.; Goss, M.J. Managing arbuscular mycorrhizal fungi for bioprotection: Mn toxicity. Soil Biol. Biochem. 2014, 68, 78–84. [Google Scholar] [CrossRef]
- Albino, U.B.; Andrade, G. Evaluation of the Functional Group of Microorganisms as Bioindicators on the Rhizosphere Microcosm. In Handbook of Microbial Biofertilizers; Rai, M.K., Ed.; Haworth: New York, NY, USA, 2007; Chapter 2; p. 532. [Google Scholar]
- Goulart, K.C.S. Perfil Metagenômico de Solo Sob Cultivo de Cana-de-Açúcar com Perspectiva na Produção de Bioenergia. Ph.D. Thesis, Universidade Estadual Paulista, Sao Paulo, Brazil, 2013. [Google Scholar]
- Souza, L.M.; Schlemmer, F.; Alencar, P.M.; Lopes, A.A.D.C.; Passos, S.R.; Xavier, G.R.; Fernandes, M.F.; Mendes, I.d.C.; Reis Junior, F.B.d. Estrutura metabólica e genética de comunidades bacterianas em solo de cerrado sob diferentes manejos. Pesqui. Agropecu. Bras. 2012, 47, 269–276. [Google Scholar] [CrossRef]
- Ge, Z.; Du, H.; Gao, Y.; Qiu, W. Analysis on metabolic functions of stored rice microbial communities by BIOLOG ECO microplates. Front. Microbiol. 2018, 9, 1375. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ge, Z.; Poudel, D.R. Application and Optimization of Biolog EcoPlates in Functional Diversity Studies of Soil Microbial Communities. MATEC Web Conf. 2015, 22, 04015. [Google Scholar] [CrossRef]
- Janušauskaite, D.; Kadžiene, G.; Auškalniene, O. The effect of tillage system on soil microbiota in relation to soil structure. Pol. J. Environ. Stud. 2013, 22, 1387–1391. [Google Scholar]
- Young, I.M.; Ritz, K. Tillage, habitat space and function of soil microbes. Soil Tillage Res. 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Bending, G.D.; Putland, C.; Rayns, F. Changes in microbial community metabolism and labile organic matter fractions as early indicators of the impact of management on soil biological quality. Biol. Fertil. Soils 2000, 31, 78–84. [Google Scholar] [CrossRef]
- Marschner, P.; Yang, C.H.; Lieberei, R.; Crowley, D.E. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]
- Teixeira, R.A.; Soares, T.G.; Fernandes, A.R.; de Souza Braz, A.M. Grasses and legumes as cover crop in no-tillage system in northeastern Pará Brazil. Acta Amaz. 2014, 44, 411–418. [Google Scholar] [CrossRef]
- Sofo, A.; Ricciuti, P.; Fausto, C.; Mininni, A.N.; Crecchio, C.; Scagliola, M.; Malerba, A.D.; Xiloyannis, C.; Dichio, B. The metabolic and genetic diversity of soil bacterial communities depends on the soil management system and C/N dynamics: The case of sustainable and conventional olive groves. Appl. Soil Ecol. 2019, 137, 21–28. [Google Scholar] [CrossRef]
- Bucher, A.E.; Lanyon, L.E. Evaluating soil management with microbial community-level physiological profiles. Appl. Soil Ecol. 2005, 29, 59–71. [Google Scholar] [CrossRef]
- Bi, Y.; Xiao, L.; Liu, R. Response of arbuscular mycorrhizal fungi and phosphorus solubilizing bacteria to remediation abandoned solid waste of coal mine. Int. J. Coal Sci. Technol. 2019, 6, 603–610. [Google Scholar] [CrossRef]
- De La Luz Mora, M.; Rosas, A.; Ribera, A.; Rengel, Z. Differential tolerance to Mn toxicity in perennial ryegrass genotypes: Involvement of antioxidative enzymes and root exudation of carboxylates. Plant Soil 2009, 320, 79–89. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Veríssimo, A.C.S.; Pinto, D.C.G.A.; Sierra-Garcia, I.N.; Granada, C.E.; Cremades, J.; Silva, H.; Cunha, Â. Engineering the Rhizosphere Microbiome with Plant Growth Promoting Bacteria for Modulation of the Plant Metabolome. Plants 2024, 13, 2309. [Google Scholar] [CrossRef] [PubMed]
- Byrne, S.L.; Foito, A.; Hedley, P.E.; Morris, J.A.; Stewart, D.; Barth, S. Early response mechanisms of perennial ryegrass (Lolium perenne) to phosphorus deficiency. Ann. Bot. 2011, 107, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gay, L.M.; Tuveng, T.R.; Agger, J.W.; Westereng, B.; Mathiesen, G.; Horn, S.J.; Vaaje-Kolstad, G.; van Aalten, D.M.F.; Eijsink, V.G.H. Structure and function of a broad-specificity chitin deacetylase from Aspergillus nidulans FGSC A4. Sci. Rep. 2017, 7, 1746. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Prasad, V.; Zehra, A.; Gupta, V.K.; Upadhyay, R.S. Mannitol metabolism during pathogenic fungal-host interactions under stressed conditions. Front. Microbiol. 2015, 6, 1019. [Google Scholar] [CrossRef]
- Conceição, T.A.; Andrade, G.; Brito, I. Influence of Intact Mycelium of Arbuscular Mycorrhizal Fungi on Soil Microbiome Functional Profile in Wheat under Mn Stress. Plants 2022, 11, 2598. [Google Scholar] [CrossRef]
- Marschner, H. Mechanisms of adaptation of plants to acid soils. Plant Soil 1991, 134, 1–20. [Google Scholar] [CrossRef]
- Arora, P.K. Bacterial degradation of monocyclic aromatic amine. Front. Microbiol. 2015, 6, 820. [Google Scholar] [CrossRef]
- González-Moro, M.B.; González-Moro, I.; de la Peña, M.; Estavillo, J.M.; Aparicio-Tejo, P.M.; Marino, D.; González-Murua, C.; Vega-Mas, I. A Multi-Species Analysis Defines Anaplerotic Enzymes and Amides as Metabolic Markers for Ammonium Nutrition. Front. Plant Sci. 2021, 11, 632285. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; DeRito, C.M.; Shapleigh, J.P.; Madsen, E.L.; Buckley, D.H. Phenolic acid-degrading Paraburkholderia prime decomposition in forest soil. ISME Commun. 2021, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Ciarfaglia, A.; Scopa, A.; Camele, I.; Curci, M.; Crecchio, C.; Xiloyannis, C.; Palese, A.M. Soil microbial diversity and activity in a Mediterranean olive orchard using sustainable agricultural practices. Soil Use Manag. 2014, 30, 160–167. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Rice, W.A.; Clayton, G.W. Soil microbial diversity and community structure under wheat as influenced by tillage and crop rotation. Soil Biol. Biochem. 1998, 30, 1733–1741. [Google Scholar] [CrossRef]
- Robin Svensson, J.; Lindegarth, M.; Jonsson, P.R.; Pavia, H. Disturbance-diversity models: What do they really predict and how are they tested? Proc. R. Soc. B Biol. Sci. 2012, 279, 2163–2170. [Google Scholar] [CrossRef]
- Bongiorno, G.; Bünemann, E.K.; Brussaard, L.; Mäder, P.; Oguejiofor, C.U.; de Goede, R.G.M. Soil management intensity shifts microbial catabolic profiles across a range of European long-term field experiments. Appl. Soil Ecol. 2020, 154, 103596. [Google Scholar] [CrossRef]
- Zhang, X.; Johnston, E.R.; Barberán, A.; Ren, Y.; Wang, Z.; Han, X. Effect of intermediate disturbance on soil microbial functional diversity depends on the amount of effective resources. Environ. Microbiol. 2018, 20, 3862–3875. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AWCD | Shannon Diversity (H′) | Shannon Evenness (E) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil Sampling | Mean Plant | Soil Sampling | Mean Plant | Soil Sampling | Mean Plant | |||||||||||||||
| Plants | BP | AP | AD | BP | AP | AD | BP | AP | AD | |||||||||||
| O. compressus | 0.19 | g | 0.75 | c | 1.09 | a | 0.68 | A | 2.60 | c | 3.22 | a | 3.29 | a | 3.04 | A | 0.92 | 0.97 | 0.96 | 0.95 |
| L. rigidum | 0.47 | e | 1.05 | b | 0.57 | B | 2.78 | bc | 3.21 | a | 2.87 | B | 0.91 | 0.96 | 0.93 | |||||
| S. gallica | 0.61 | d | 0.43 | f | 0.41 | C | 2.82 | bc | 3.02 | ab | 2.81 | B | 0.94 | 0.94 | 0.94 | |||||
| Mean soil sampling | 0.19 | C | 0.61 | B | 0.86 | A | 2.60 | C | 2.94 | B | 3.17 | A | 0.92 | 0.94 | 0.95 | |||||
| AWCD | Shannon Diversity (H′) | Shannon Evenness (E) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil Sampling | Mean Plant | Soil Sampling | Mean Plant | Soil Sampling | Mean Plant | |||||||
| Plants | U | D | U | D | U | D | ||||||
| O. compressus | 1.06 | a | 0.90 | c | 0.98 | A | 3.43 | 3.36 | 3.39 | 1.00 | 0.97 | 0.98 |
| L. rigidum | 0.65 | e | 1.01 | b | 0.83 | D | 3.36 | 3.42 | 3.39 | 0.98 | 0.99 | 0.99 |
| S. gallica | 0.78 | d | 1.02 | b | 0.90 | C | 3.30 | 3.41 | 3.36 | 0.97 | 0.99 | 0.98 |
| No Plant | 0.91 c | 0.91 | B | 3.37 | 3.37 | 0.99 | 0.99 | |||||
| Mean soil sampling | 0.85 | B | 0.96 | A | 3.37 | 3.39 | 0.98 | 0.99 | ||||
| (A) | |||
| Variable C-Type | PC1 | PC2 | |
| 1 | Amines and amides | 0.333 | 0.498 |
| 2 | Amino acids | 0.413 | 0.036 |
| 3 | Carboxylic acids | 0.422 | −0.167 |
| 4 | Phenolic acids | 0.328 | 0.457 |
| 5 | P_Cabon | 0.371 | −0.542 |
| 6 | Carbohydrates | 0.400 | −0.390 |
| 7 | Polymers | 0.368 | 0.261 |
| (B) | |||
| Variable C Source | PC1 | PC2 | |
| 1 | phenylethylamine | 0.174 | −0.254 |
| 1 | putrescine | 0.199 | −0.049 |
| 2 | L-arginine | 0.183 | −0.219 |
| 2 | L-asparagine | 0.148 | 0.189 |
| 2 | L-serine | 0.204 | −0.178 |
| 2 | L-threonine | 0.082 | 0.305 |
| 2 | L-phenylalanine | 0.201 | −0.210 |
| 2 | glycyl-L-glutamic acid | 0.168 | 0.120 |
| 3 | pyruvic acid methyl ester | 0.025 | 0.333 |
| 3 | D-glucosaminic acid | 0.193 | −0.213 |
| 3 | D-galactonic acid γ-lactone | 0.195 | 0.115 |
| 3 | D-galacturonic acid | 0.238 | 0.047 |
| 3 | γ-hydroxybutyric acid | 0.156 | −0.020 |
| 3 | itaconic Acid | 0.104 | 0.257 |
| 3 | α-ketobutyric acid | 0.134 | −0.267 |
| 3 | D-malic acid | 0.213 | −0.005 |
| 4 | 2-hydroxy benzoic acid | 0.190 | −0.118 |
| 4 | 4-hydroxy benzoic acid | 0.187 | −0.057 |
| 5 | glucose-1-phosphate | 0.203 | 0.176 |
| 5 | D,L-α-glycerol phosphate | 0.227 | −0.057 |
| 6 | D-cellobiose | 0.203 | 0.188 |
| 6 | α-D-lactose | 0.203 | 0.191 |
| 6 | β-methyl-D-glucoside | 0.207 | 0.173 |
| 6 | D-xylose | 0.169 | −0.274 |
| 6 | i-erythritol | 0.173 | −0.046 |
| 6 | D-mannitol | 0.190 | 0.199 |
| 6 | N-acetyl-D-glucosamine | 0.231 | 0.125 |
| 7 | tween 40 | 0.143 | 0.082 |
| 7 | tween 80 | 0.092 | 0.108 |
| 7 | α-cyclodextrin | 0.170 | 0.083 |
| 7 | glycogen | 0.181 | −0.216 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conceição, T.; Andrade, G.; Brito, I. Carbon Metabolic Profiling as a Response to Previous Plant Mycotrophy and Soil Disturbance in Wheat Growth. Appl. Microbiol. 2024, 4, 1661-1676. https://doi.org/10.3390/applmicrobiol4040113
Conceição T, Andrade G, Brito I. Carbon Metabolic Profiling as a Response to Previous Plant Mycotrophy and Soil Disturbance in Wheat Growth. Applied Microbiology. 2024; 4(4):1661-1676. https://doi.org/10.3390/applmicrobiol4040113
Chicago/Turabian StyleConceição, Taiana, Galdino Andrade, and Isabel Brito. 2024. "Carbon Metabolic Profiling as a Response to Previous Plant Mycotrophy and Soil Disturbance in Wheat Growth" Applied Microbiology 4, no. 4: 1661-1676. https://doi.org/10.3390/applmicrobiol4040113
APA StyleConceição, T., Andrade, G., & Brito, I. (2024). Carbon Metabolic Profiling as a Response to Previous Plant Mycotrophy and Soil Disturbance in Wheat Growth. Applied Microbiology, 4(4), 1661-1676. https://doi.org/10.3390/applmicrobiol4040113