1. Introduction
The Parkinson’s disease is characterized by a spectrum of symptoms that can differ in nature and intensity, making the prediction of treatment outcomes challenging [
1]. Interestingly, the therapeutic potential of hyperbaric oxygen therapy (HBOT) for Parkinson’s was sometimes identified in an unanticipated manner [
2]. For example, a diabetic patient undergoing HBOT for a foot ulcer unexpectedly reported a marked alleviation in Parkinson’s symptoms. Animal studies consistently indicate that HBOT exhibits anti-inflammatory properties [
3], which could be beneficial in addressing the inflammatory conditions observed in the substantia nigra region of the brain in Parkinson’s patients. Anecdotal evidence further suggests that some Parkinson’s patients, even those with advanced stages of the disease, have shown significant improvements after HBOT sessions.
The realm of hyperbaric oxygen therapy (HBOT) in treating neurodegenerative diseases [
4], particularly Parkinson’s disease (PD), is burgeoning with potential. As we delve deeper into this field, a multitude of studies have emerged, shedding light on the transformative effects of HBOT on neuronal health, motor function, and overall quality of life for patients. For instance, recent research has illuminated the capacity of HBOT to target specific brain circuits, enhance neurotrophic factors, and even modulate epigenetic pathways, offering a beacon of hope for those grappling with the debilitating effects of PD.
For instance, research has demonstrated that HBOT can significantly increase the number of TH-positive neurons in MPTP-treated mice, enhancing the neurotrophic factor BDNF while reducing apoptotic signaling and attenuating inflammatory mediators in the midbrain [
5]. This treatment also promotes mitochondrial biogenesis and improves locomotor activity and grip strength in these mice.
Further insights [
6] highlighted the potential of HBOT in targeting specific brain circuits involved in “Kinesia Paradoxa”, including the noradrenergic system, basal ganglia, and the cerebellum circuit. This study presented evidence supporting the “Norepinephrine Hypothesis”, suggesting a role for HBOT in increasing norepinephrine levels, which could restore motor deficits in Parkinson’s disease patients. When considering the combination of treatments, the research indicates that combining donepezil with HBOT and functional rehabilitation training can significantly enhance therapeutic effectiveness in Parkinson’s disease dementia (PDD) patients. This combination not only improves cognitive function, self-care ability, and quality of life but also significantly reduces inflammatory markers like serum IL-1β and IL-6 [
7].
In a broader context, the potential of HBOT as a therapeutic intervention for neurodegenerative diseases has been explored, with findings emphasizing its promising effects in conditions associated with neurodegeneration and functional impairments. A special focus has been given to the role of epigenetics in these effects [
8]. Lastly, in a study focused on spinocerebellar ataxias (SCAs), HBOT was found to attenuate motor coordination and cognitive impairment in SCA17 mice, with effects persisting for about a month post-treatment. SCA17 is a rare subtype of SCAs (spinocerebellar ataxias), notable for its association with a myriad of neurological symptoms including motor coordination and cognitive impairments, often leading to a substantial reduction in the quality of life of affected individuals.
This neuroprotective effect of HBOT might be attributed to the promotion of BDNF production and the reduction of neuroinflammation [
9].
Despite the promising strides made in this domain, the field is still in its infancy. The intricacies of HBOT’s impact on the human brain, especially in the context of neurodegenerative diseases, remain vast and largely uncharted. While the preliminary results are indeed encouraging, they underscore the pressing need for more comprehensive, large-scale studies. Only through rigorous research, meticulous analysis, and collaborative efforts can we truly harness the full potential of HBOT and pave the way for groundbreaking therapeutic interventions in the future.
On the other hand, machine learning (ML) methodologies have been extensively applied to enhance the understanding and management of Parkinson’s disease (PD). A comprehensive review of the literature reveals the utilization of ML models in conjunction with Internet of Things technologies, such as smart devices and various sensors, to optimize predictions and estimations regarding different aspects of PD [
10]. These models are trained on data acquired via these technologies and address a myriad of PD-related problems, offering insights into the most effective algorithms and commonly addressed issues in PD management. Another study provides an extensive overview of the application of ML in categorizing PD, emphasizing the use of diverse data modalities and artificial intelligence techniques to facilitate informed and systematic clinical decision-making [
11]. These studies collectively underscore the pivotal role of ML in advancing diagnostic processes and therapeutic interventions for PD, highlighting its potential in contributing to more nuanced and effective approaches in PD treatment and management.
The exploration of hyperbaric oxygen therapy (HBOT) in the context of Parkinson’s disease (PD) has predominantly been rooted in traditional research methodologies [
12]. Notably absent from this landscape is the integration of modern machine learning (ML) frameworks, which have the potential to revolutionize our understanding of the disease’s intricacies [
13]. While several studies have explored molecular biology to understand underlying mechanisms, many have not fully utilized advanced computational methods. Our endeavor represents a pioneering effort in this direction. By employing an ML approach, we aim to meticulously examine the behavior of key genes implicated in PD. This innovative methodology allows us to unravel the intricate relationships between these genes and the therapeutic effects of HBOT, offering a fresh perspective and potentially groundbreaking insights into the treatment of PD.
3. Results and Discussion
We ended up with a combined list of genes (
Table S1) that are strongly related to PD and decided to explore closely at the top 100 genes since it’s been demonstrated that in scRNA-seq, typically only a few dozen to a couple of hundred genes play a pivotal role in the dataset [
18]. Also, focusing on the top 100 genes facilitated a more in-depth exploration of their biological functions, interactions, and roles in the context of the study, allowing for more meaningful interpretations and conclusions.
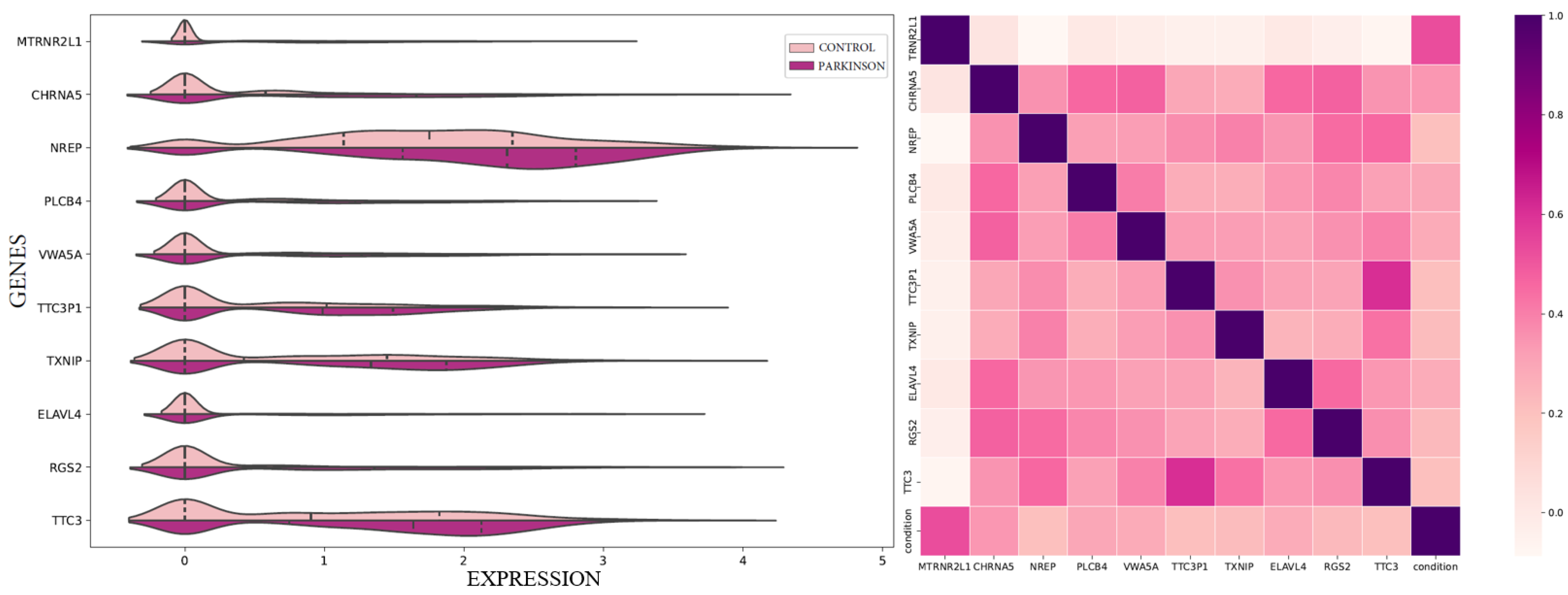
Also, by concentrating on the top genes, we’re likely capturing the most important ones that have the biggest impact on PD. A key part of our study was to see how these genes are related to HBOT. This is important because if we know which genes are affected by this therapy, it could help doctors treat PD more effectively in the future.
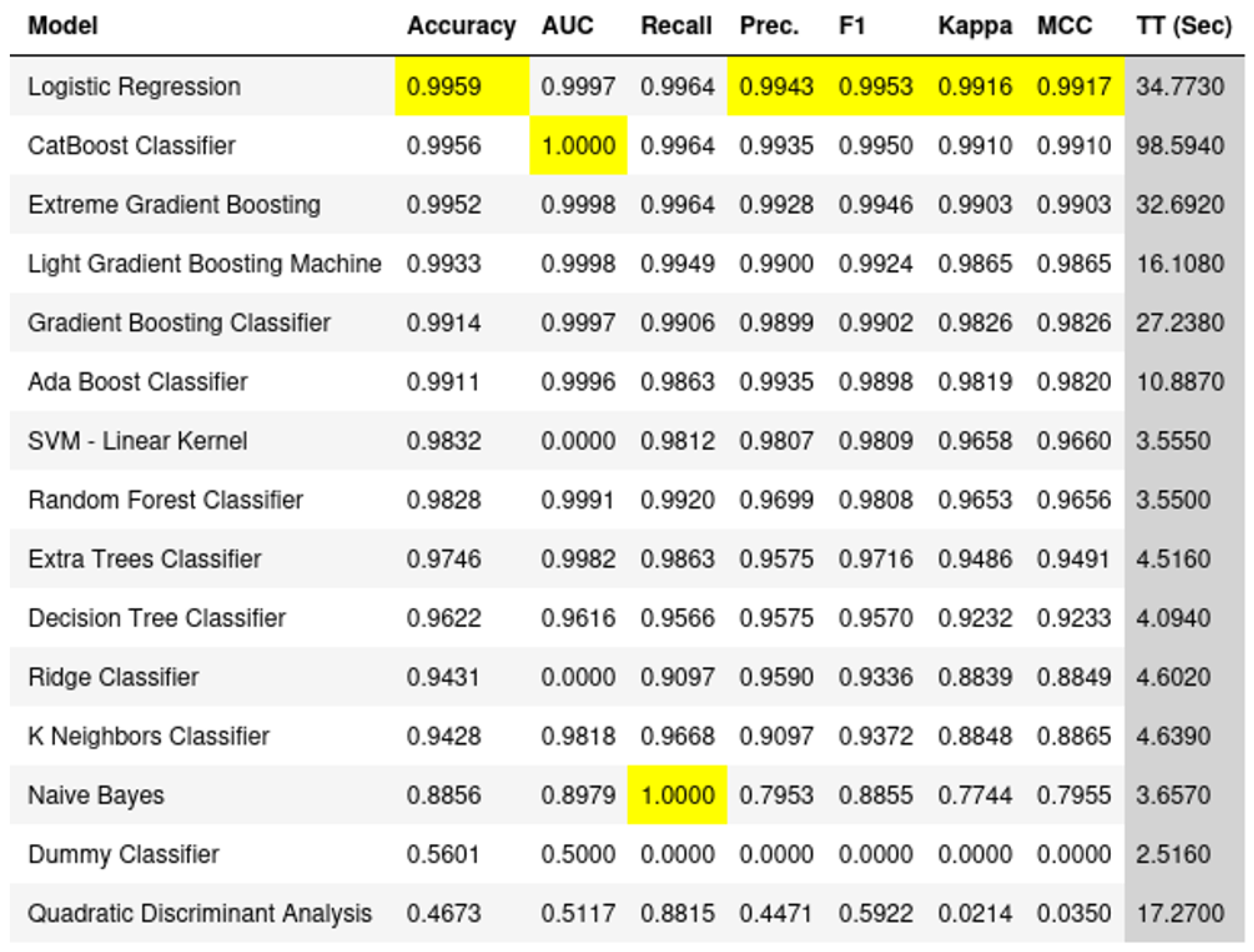
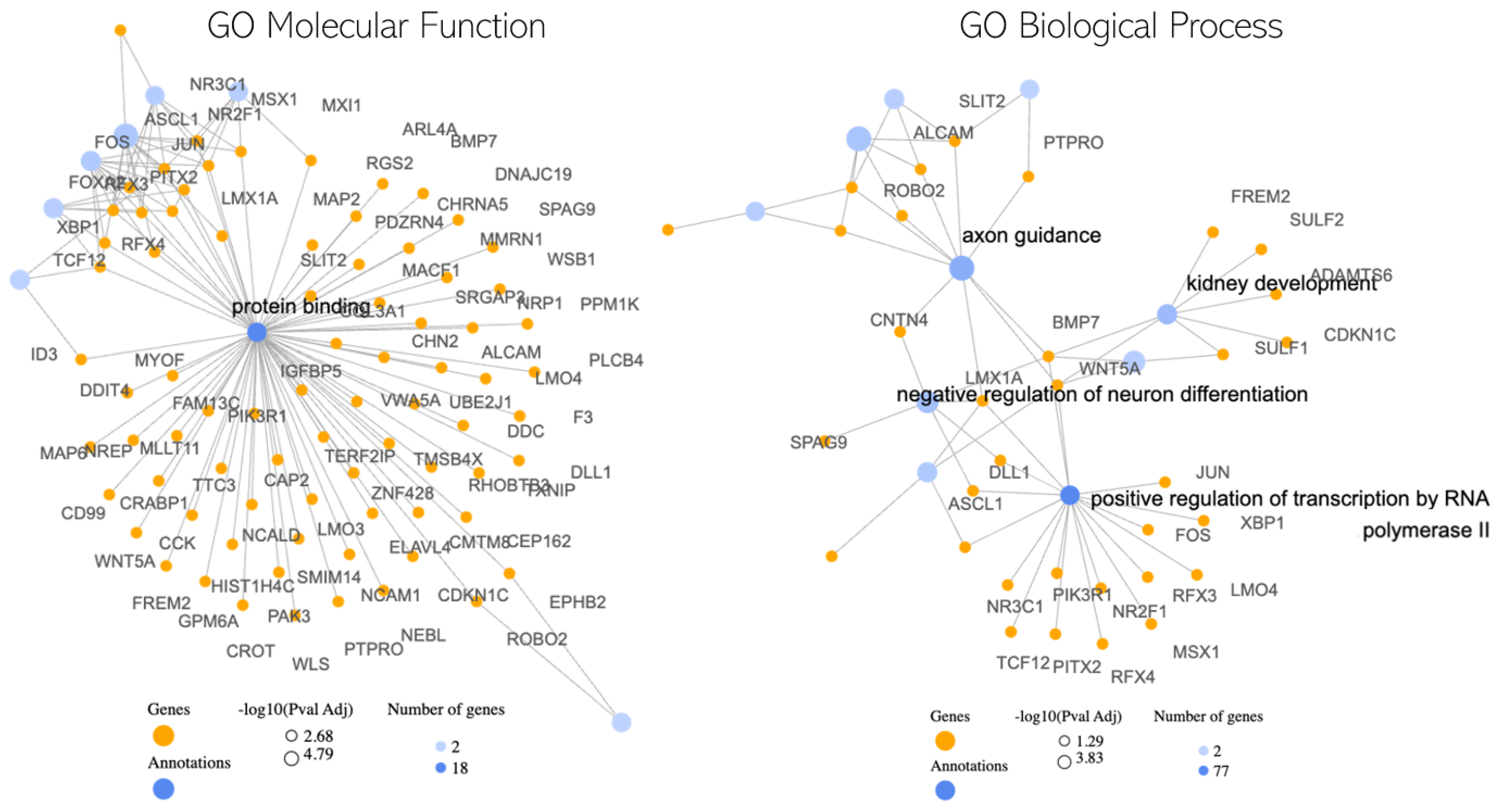
Our study had three four parts. First, we examine the classification performance using the 100 key genes regarding differentiating healthy samples from Parkinson’s disease samples. Furthermore, we looked at how HBOT affects each of the top 100 genes one by one. This helped us figure out which specific genes might be good targets for treatment. Next, we checked how our chosen genes fit into bigger groups of genes and how they might be linked to other diseases or treatments. This gave us a better idea of the bigger picture and how these genes work in the body. Lastly, we used a simple visual tool zooming out of the genes to show how our top genes are connected to each other along with the associated gene ontologies.
3.4. Graph-Based Analysis—Interconnectivity and Associations
In our graph-based analysis (
Figure 4), we focused on understanding the relationships between genes and their associated gene ontologies, which describe their roles in molecular functions and broader biological processes. A key aspect of this was examining gene-gene interactions using protein-protein interaction (PPI) networks. These PPI networks provide a structured representation of how proteins, and by extension the genes that code for them, interact within a cell. By mapping our selected genes onto these networks, we gained insights into potential functional relationships that these genes might have with one another.
Alongside this, we aimed to determine how our genes fit within larger biological contexts. To do this, we conducted an enrichment analysis, which checks if certain biological categories or functions are more common among our selected genes than would be expected by chance [
25]. It uses the standard hypergeometric distribution test, also known as the Fisher exact test, for this purpose, a widely accepted statistical method in gene enrichment analysis. By comparing our gene set to reference sets, this test helped us identify specific biological processes or molecular functions that our genes are likely involved in.
In our exploration of the potential molecular interplay between hyperbaric oxygen therapy (HBOT) and Parkinson’s disease (PD), the graph-based analysis has revealed several noteworthy findings. The central observation is the pronounced role of the “Protein Binding” term. Acting as a hub, this term suggests a nexus of interactions, with several genes like MAP2, CAP2, and WSB1 being pivotal [
26,
27,
28]. The hub-like nature of “Protein Binding” implies that these genes might be central to many protein–protein interactions, potentially modulating a variety of cellular processes that could be influenced by HBOT in the context of PD [
29].
Among these genes, for instance, MAP2 is known for its role in stabilizing microtubules [
30], which are essential for maintaining cell structure and facilitating intracellular transport. Any modulation in its activity could impact neuronal health and function, making it a potential target of interest in PD and its response to HBOT. Furthermore, other gene ontology (GO) terms that stood out include “axon guidance” [
31], “negative regulation of neuron differentiation” [
32], and “positive regulation of transcription by RNA polymerase II” [
33]. The presence of “axon guidance” is particularly intriguing, as it plays a crucial role in the proper formation of neural circuits. Disruptions in this process could contribute to neurodegenerative conditions like PD. The regulation of neuron differentiation and transcription further suggests that HBOT might influence the broader landscape of gene expression and neuronal development in PD.
In considering the translational potential of our findings, it is pivotal to acknowledge the prospective clinical implications. The identified genomic correlations with hyperbaric oxygen therapy (HBOT) in Parkinson’s disease (PD) suggest avenues for the development of personalized and optimized treatment strategies, potentially enhancing therapeutic outcomes and patient quality of life. These genomic insights could inform the creation of targeted therapies and predictive models, allowing for individualized treatment plans based on specific genomic profiles. However, the realization of these clinical applications necessitates rigorous validation through clinical trials, collaborative integration into clinical workflows, adherence to ethical and regulatory standards, and comprehensive educational outreach to stakeholders about the benefits and limitations of such interventions.
Our study, while offering significant insights, is subject to several limitations. The single-cell RNA-seq data utilized may not fully capture the intricate cellular heterogeneity inherent to Parkinson’s disease due to its inherent resolution and depth limitations, and the public datasets employed may harbor biases stemming from variations in sample collection, processing, and sequencing technologies across different studies. Additionally, the machine learning techniques applied in our analysis are susceptible to biases from the training data, model assumptions, and algorithmic constraints, potentially impacting the reliability of our identified gene correlations. Furthermore, the generalizability of our findings is constrained, necessitating validation in diverse and larger Parkinson’s disease populations to confirm their universal applicability and clinical relevance.
Author Contributions
Conceptualization, E.B. and P.V.; methodology, E.B. and A.G.V.; software, E.B.; validation, A.G.V. and M.G.K.; formal analysis, E.B. and A.G.V.; data curation, E.B. and A.G.V.; writing—original draft preparation, E.B. and A.G.V.; writing—review and editing, M.G.K. and P.V.; supervision, P.V.; funding acquisition, P.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research is funded by the European Union and Greece (Partnership Agreement for the Development Framework 2014–2020) under the Regional Operational Programme Ionian Islands 2014–2020, project title: “Study of Clinical trial protocols with biomarkers that define the evolution of non-genetic neurodegenerative diseases- NEUROTRIAL”, project number: 5016089.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This study uses a public dataset available at Gene Expression Omnibus. The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Wilmshurst, P.; Bewley, S.; Murray, P. Hyperbaric oxygen therapy for the treatment of long COVID. Clin. Med. 2023, 23, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Taslipinar, M.Y.; Aydin, I.; Kaldirim, U.; Aydin, F.N.; Agilli, M.; Eyi, Y.E.; Cayci, T. Hyperbaric oxygen treatment and N-acetylcysteine ameliorate acetaminophen-induced liver injury in a rat model. Hum. Exp. Toxicol. 2013, 32, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Kidd, P.M. Multiple sclerosis, an autoimmune inflammatory disease: Prospects for its integrative management. Altern. Med. Rev. 2001, 6, 540–566. [Google Scholar] [PubMed]
- Hsu, H.T.; Yang, Y.L.; Chang, W.H.; Fang, W.Y.; Huang, S.H.; Chou, S.H.; Lo, Y.C. Hyperbaric oxygen therapy improves Parkinson’s disease by promoting mitochondrial biogenesis via the SIRT-1/PGC-1α pathway. Biomolecules 2022, 12, 661. [Google Scholar] [CrossRef]
- Banou, E. Hyperbaric oxygen therapy effect on “Kinesia Paradoxa” brain circuits. GeNeDis 2020: Genetics and Neurodegenerative Diseases. GeNeDis 2020: Genet. Neurodegener. Dis. 2021; 1339, 139–146. [Google Scholar]
- Fan, A.; Zhou, J. Effect of the combination of donepezil with hyperbaric oxygen therapy and functional rehabilitation training on parkinson’s disease dementia and the neurological function system. Int. J. Clin. Exp. Med. 2020, 13, 5867–5875. [Google Scholar]
- Mensah-Kane, P.; Sumien, N. The potential of hyperbaric oxygen as a therapy for neurodegenerative diseases. GeroScience 2023, 45, 747–756. [Google Scholar] [CrossRef]
- Shi, Q.; Luo, Q.; Gong, Q.; Wang, G. Effects of rTMS Combined with Hyperbaric Oxygen-acupuncture-rehabilitation Therapy on Motor Function, Serum CRP and Plasma Dopamine in Patients with Parkinson’s Disease. Chin. Gen. Pract. 2020, 23, 3460. [Google Scholar]
- Giannakopoulou, K.M.; Roussaki, I.; Demestichas, K. Internet of things technologies and machine learning methods for Parkinson’s disease diagnosis, monitoring and management: A systematic review. Sensors 2022, 22, 1799. [Google Scholar] [CrossRef]
- Rana, A.; Dumka, A.; Singh, R.; Panda, M.K.; Priyadarshi, N.; Twala, B. Imperative role of machine learning algorithm for detection of Parkinson’s disease: Review, challenges and recommendations. Diagnostics 2022, 12, 2003. [Google Scholar] [CrossRef]
- Atzeni, F.; Masala, I.F.; Cirillo, M.; Boccassini, L.; Sorbara, S.; Alciati, A. Hyperbaric oxygen therapy in fibromyalgia and the diseases involving the central nervous system. Clin. Exp. Rheumatol. 2020, 38, 0094–0098. [Google Scholar]
- Quazi, S. Artificial intelligence and machine learning in precision and genomic medicine. Med. Oncol. 2022, 39, 120. [Google Scholar] [CrossRef] [PubMed]
- Novak, G.; Kyriakis, D.; Grzyb, K.; Bernini, M. Single-cell transcriptomics of human iPSC differentiation dynamics reveal a core molecular network of Parkinson’s disease. Commun. Biol. 2022, 5, 49. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ge, X.; Peng, F.; Li, W.; Li, J.J. Wilcoxon rank-sum test still outperforms dearseq after accounting for the normalization impact in semi-synthetic RNA-seq data simulation. bioRxiv 2022, 2022-06. [Google Scholar] [CrossRef]
- Chen, T.; He, T.; Benesty, M.; Khotilovich, V.; Tang, Y.; Cho, H.; Zhou, T. Xgboost: Extreme gradient boosting. R Package Version 0.4-2 2015, 1, 1–4. [Google Scholar]
- Paplomatas, P.; Krokidis, M.G.; Vlamos, P.; Vrahatis, A.G. An ensemble feature selection approach for analysis and modeling of transcriptome data in alzheimer’s disease. Appl. Sci. 2023, 13, 2353. [Google Scholar] [CrossRef]
- Chatzilygeroudis, K.I.; Vrahatis, A.G.; Tasoulis, S.K.; Vrahatis, M.N. Feature Selection in single-cell RNA-seq data via a Genetic Algorithm. In Proceedings of the Learning and Intelligent Optimization: 15th International Conference, LION 15, Athens, Greece, 20–25 June 2021; Revised Selected Papers 15. Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 66–79. [Google Scholar]
- Tezgin, D.; Giardina, C.; Perdrizet, G.A.; Hightower, L.E. The effect of hyperbaric oxygen on mitochondrial and glycolytic energy metabolism: The caloristasis concept. Cell Stress Chaperones 2020, 25, 667–677. [Google Scholar] [CrossRef]
- Bowles, K.R.; Silva, M.C.; Whitney, K.; Bertucci, T.; Berlind, J.E.; Lai, J.D.; Temple, S. ELAVL4, splicing, and glutamatergic dysfunction precede neuron loss in MAPT mutation cerebral organoids. Cell 2021, 184, 4547–4563. [Google Scholar] [CrossRef]
- Iwakoshi, N.N.; Lee, A.H.; Glimcher, L.H. The X-box binding protein-1 transcription factor is required for plasma cell differentiation and the unfolded protein response. Immunol. Rev. 2003, 194, 29–38. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 1–14. [Google Scholar] [CrossRef]
- Zhou, L.; Too, H.P. Mitochondrial localized STAT3 is involved in NGF induced neurite outgrowth. PLoS ONE 2011, 6, e21680. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Pathoanatomy of Parkinson’s disease. J. Neurol. 2000, 247, II3–II10. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moreno, A.; López-Domínguez, R.; Villatoro-García, J.A.; Ramirez-Mena, A.; Aparicio-Puerta, E.; Hackenberg, M.; Pascual-Montano, A.; Carmona-Saez, P. Functional Enrichment Analysis of Regulatory Elements. Biomedicines 2022, 10, 590. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.R.; Ilyin, S.; Plata-Salaman, C.R. Abnormal patterns of microtubule-associated protein-2 (MAP-2) immunolabeling in neuronal nuclei and Lewy bodies in Parkinson’s disease substantia nigra brain tissues. Neurosci. Lett. 2001, 306, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, A.; De Rosa, A.; Pelucchi, S.; Garofalo, M.; Marciano, B.; Nuzzo, T.; Usiello, A. Analysis of mRNA and protein levels of CAP2, DLG1 and ADAM10 genes in post-mortem brain of schizophrenia, Parkinson’s and Alzheimer’s disease patients. Int. J. Mol. Sci. 2022, 23, 1539. [Google Scholar] [CrossRef] [PubMed]
- Nucifora Jr, F.C.; Nucifora, L.G.; Ng, C.H.; Arbez, N.; Guo, Y.; Roby, E.; Ross, C.A. Ubiqutination via K27 and K29 chains signals aggregation and neuronal protection of LRRK2 by WSB1. Nat. Commun. 2016, 7, 11792. [Google Scholar] [CrossRef] [PubMed]
- Ruf, W.P.; Freischmidt, A.; Grozdanov, V.; Roth, V.; Brockmann, S.J.; Mollenhauer, B.; Danzer, K.M. Protein binding partners of dysregulated miRNAs in Parkinson’s Disease Serum. Cells 2021, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005, 6, 1–10. [Google Scholar]
- Lesnick, T.G.; Papapetropoulos, S.; Mash, D.C.; Ffrench-Mullen, J.; Shehadeh, L.; De Andrade, M.; Maraganore, D.M. A genomic pathway approach to a complex disease: Axon guidance and Parkinson disease. PLoS Genet. 2007, 3, e98. [Google Scholar] [CrossRef]
- Pirooznia, S.K.; Wang, H.; Panicker, N.; Kumar, M.; Neifert, S.; Dar, M.A.; Dawson, T.M. Deubiquitinase CYLD acts as a negative regulator of dopamine neuron survival in Parkinson’s disease. Sci. Adv. 2022, 8, eabh1824. [Google Scholar] [CrossRef]
- Majidinia, M.; Mihanfar, A.; Rahbarghazi, R.; Nourazarian, A.; Bagca, B.; Avci, Ç.B. The roles of non-coding RNAs in Parkinson’s disease. Mol. Biol. Rep. 2016, 43, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).







{kind=link}
{kind=link}
{kind=link}
{kind=link}