
Anomaly Detection Utilizing One-Class Classification—A Machine Learning Approach for the Analysis of Plant Fast Fluorescence Kinetics

Abstract
1. Introduction
2. Results and Discussion
2.1. Data Overview
2.2. Screening of “Anomalies” for Model Fine-Tuning
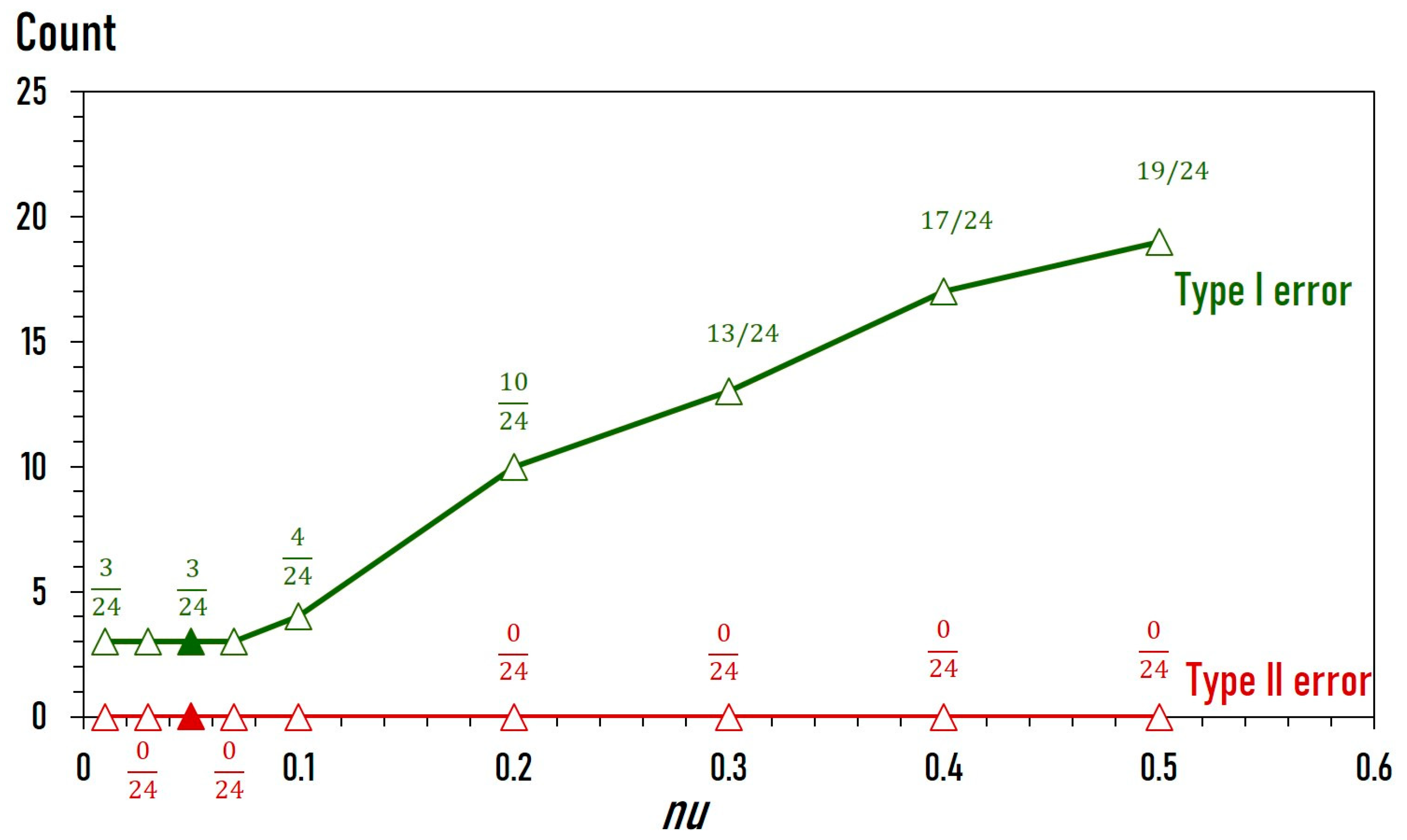
2.3. Training and Fine-Tuning of Classification Model
2.4. Classification of the Remaining Data
2.5. Comparison Between “Normal” and “Anomaly” Measurements
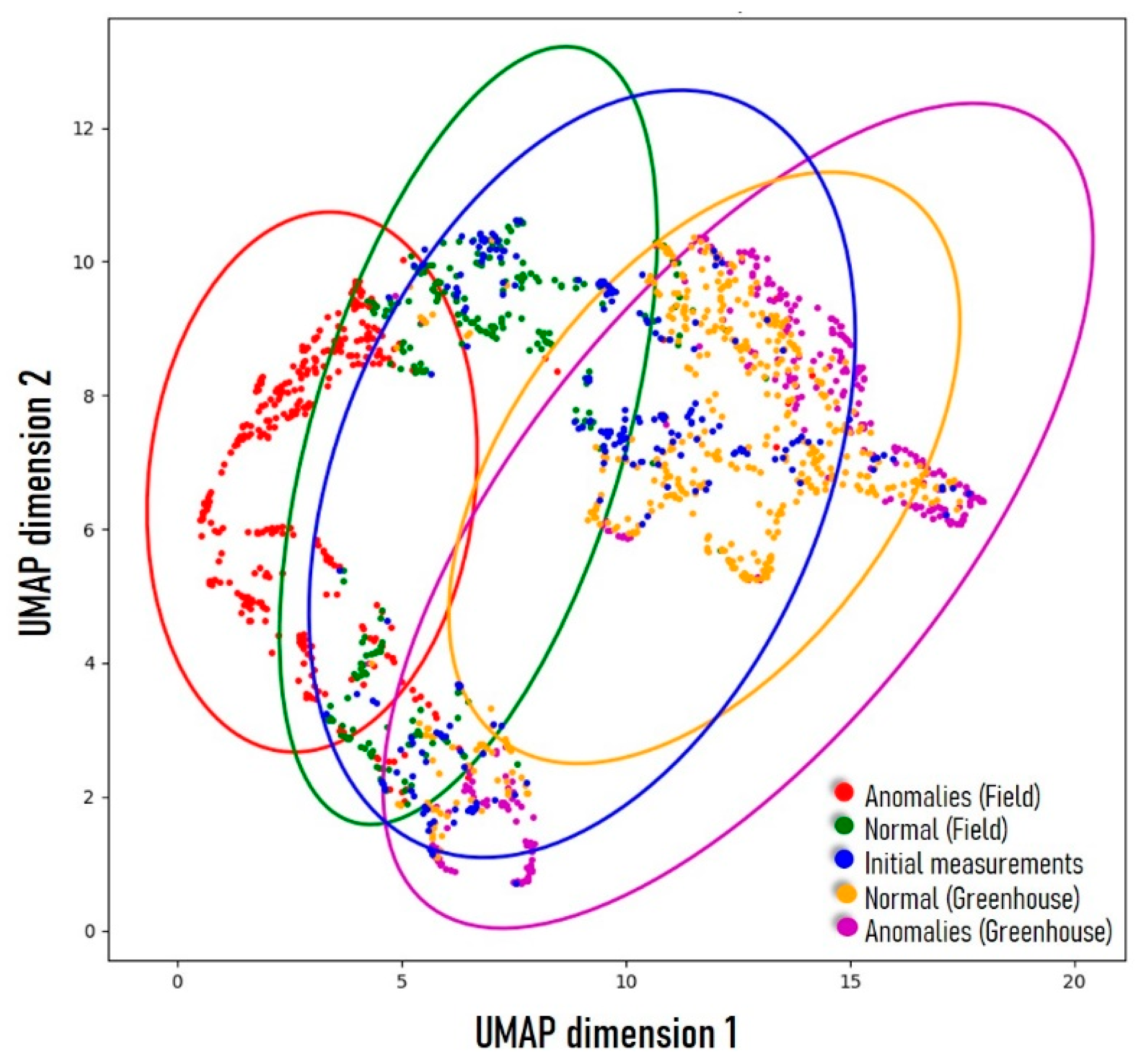
2.6. UMAP Visualization of All Measurements
2.7. k-Means Clustering of “Anomalies”
2.8. Comparison Between “Anomalies” Types 1 and 2
2.9. Interpretation of the Results
2.10. Summary and Outlook
3. Materials and Methods
3.1. Plant Cultivation
3.2. Measurements of Fast Chlorophyll Fluorescence Kinetics
3.3. Data Overview and Data Pre-Processing
3.4. One-Class Support Vector Machine
3.5. UMAP Data Visualization
3.6. k-Means Clustering
3.7. Statistical Analysis
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic Chlorophyll a Fluorescence Transient in Plants and Cyanobacteria*. Photochem. Amp. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about In Vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jajoo, A.; Mehta, P.; Bharti, S. Analysis of elevated temperature—Induced inhibition of photosystem II using chlorophyll a fluorescence induction kinetics in wheat leaves (Triticum aestivum). Plant Biol. 2010, 13, 1–6. [Google Scholar] [CrossRef]
- Yang, J.; Kong, Q.; Xiang, C. Effects of low night temperature on pigments, chl a fluorescence and energy allocation in two bitter gourd (Momordica charantia L.) genotypes. Acta Physiol. Plant. 2008, 31, 285–293. [Google Scholar] [CrossRef]
- Jedmowski, C.; Brüggemann, W. Imaging of fast chlorophyll fluorescence induction curve (OJIP) parameters, applied in a screening study with wild barley (Hordeum spontaneum) genotypes under heat stress. J. Photochem. Photobiol. B Biol. 2015, 151, 153–160. [Google Scholar] [CrossRef]
- Singh-Tomar, R.; Mathur, S.; Allakhverdiev, S.I.; Jajoo, A. Changes in PS II heterogeneity in response to osmotic and ionic stress in wheat leaves (Triticum aestivum). J. Bioenerg. Biomembr. 2012, 44, 411–419. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Loboda, T. Photosystem II of barley seedlings under cadmium and lead stress. Plant Soil Environ. 2007, 53, 511–516. [Google Scholar] [CrossRef]
- Redillas, M.C.F.R.; Jeong, J.S.; Strasser, R.J.; Kim, Y.S.; Kim, J.-K. JIP analysis on rice (Oryza sativa cv Nipponbare) grown under limited nitrogen conditions. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 827–832. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. In Acta Physiologiae Plantarum; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; Volume 38. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M. Special issue in honour of Prof. Reto J. Strasser—Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynt. 2020, 58, 275–292. [Google Scholar] [CrossRef]
- Stirbet, A. Govindjee On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Srivastava, A.; Strasser, R.J.; Govindjee. Greening of Peas: Parallel Measurements of 77 K Emission Spectra, OJIP Chlorophyll a Fluorescence Transient, Period Four Oscillation of the Initial Fluorescence Level, Delayed Light Emission, and P700. Photosynthetica 1999, 36, 365. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Strasser, R.J. In Vivo Assessment of Stress Impact on Plant’s Vitality: Applications in Detecting and Evaluating the Beneficial Role of Mycorrhization on Host Plants. Mycorrhiza 2008, 679–703. [Google Scholar] [CrossRef]
- Strauss, A.J.; Krüger, G.H.J.; Strasser, R.J.; Heerden, P.D.R.V. Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient O-J-I-P. Environ. Exp. Bot. 2006, 56, 147–157. [Google Scholar] [CrossRef]
- Oukarroum, A.; Madidi, S.E.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and re-watering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar] [CrossRef]
- Jedmowski, C.; Bayramov, S.; Brüggemann, W. Comparative analysis of drought stress effects on photosynthesis of Eurasian and North African genotypes of wild barley. Photosynthetica 2014, 52, 564–573. [Google Scholar] [CrossRef]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Duarte, B.; Pedro, S.; Marques, J.C.; Adão, H.; Caçador, I. Zostera noltii development probing using chlorophyll a transient analysis (JIP-test) under field conditions: Integrating physiological insights into a photochemical stress index. Ecol. Indic. 2017, 76, 219–229. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee, G. Chlorophyll a fluorescence induction: Can just a one-second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. In Nature; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2015; Volume 521, pp. 436–444. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Oukarroum, A.; Alexandrov, V.; Kouzmanova, M.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Allakhverdiev, S.I.; Goltsev, V. Identification of nutrient deficiency in maize and tomato plants by In Vivo chlorophyll a fluorescence measurements. Plant Physiol. Biochem. 2014, 81, 16–25. [Google Scholar] [CrossRef]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I. Drought-induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1490–1498. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bąba, W.; Gediga, K.; Goltsev, V.; Samborska, I.A.; Cetner, M.D.; Dimitrova, S.; Piszcz, U.; Bielecki, K.; Karmowska, K. Chlorophyll fluorescence as a tool for nutrient status identification in rapeseed plants. Photosynth. Res. 2017, 136, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.; Liu, Y.; Captoline, I.; Li, X.; Ye, D.; Wu, R. Citrus Huanglongbing detection based on polyphasic chlorophyll a fluorescence coupled with machine learning and model transfer in two citrus cultivars. Comput. Electron. Agric. 2021, 187, 106289. [Google Scholar] [CrossRef]
- Bresson, J.; Vasseur, F.; Dauzat, M.; Koch, G.; Granier, C.; Vile, D. Quantifying spatial heterogeneity of chlorophyll fluorescence during plant growth and in response to water stress. Plant Methods 2015, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, H.; Chen, H.; Fan, C.; Tai, Y.; Chen, X.; Zhang, W.; He, T.; Gao, Z. Spatiotemporal Heterogeneity of Chlorophyll Content and Fluorescence Response within Rice (Oryza sativa L.) Canopies under Different Cadmium Stress. Agronomy 2022, 13, 121. [Google Scholar] [CrossRef]
- Arief, M.A.A.; Kim, H.; Kurniawan, H.; Nugroho, A.P.; Kim, T.; Cho, B.-K. Chlorophyll Fluorescence Imaging for Early Detection of Drought and Heat Stress in Strawberry Plants. Plants 2023, 12, 1387. [Google Scholar] [CrossRef]
- Seliya, N.; Abdollah Zadeh, A.; Khoshgoftaar, T.M. A literature review on one-class classification and its potential applications in big data. J. Big Data 2021, 8, 122. [Google Scholar] [CrossRef]
- Tran, N.T.; Jokic, L.; Keller, J.; Geier, J.U.; Kaldenhoff, R. Impacts of Radio-Frequency Electromagnetic Field (RF-EMF) on Lettuce (Lactuca sativa)—Evidence for RF-EMF Interference with Plant Stress Responses. Plants 2023, 12, 1082. [Google Scholar] [CrossRef]
- Shomali, A.; Aliniaeifard, S.; Bakhtiarizadeh, M.R.; Lotfi, M.; Mohammadian, M.; Vafaei Sadi, M.S.; Rastogi, A. Artificial neural network (ANN)-based algorithms for high light stress phenotyping of tomato genotypes using chlorophyll fluorescence features. Plant Physiol. Biochem. 2023, 201, 107893. [Google Scholar] [CrossRef]
- Jedmowski, C.; Ashoub, A.; Brüggemann, W. Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiol. Plant. 2012, 35, 345–354. [Google Scholar] [CrossRef]
- Küpper, H.; Benedikty, Z.; Morina, F.; Andresen, E.; Mishra, A.; Trtílek, M. Analysis of OJIP Chlorophyll Fluorescence Kinetics and QA Reoxidation Kinetics by Direct Fast Imaging. Plant Physiol. 2018, 179, 369–381. [Google Scholar] [CrossRef]
- Morales, L.O.; Shapiguzov, A.; Safronov, O.; Leppälä, J.; Vaahtera, L.; Yarmolinsky, D.; Kollist, H.; Brosché, M. Ozone responses in Arabidopsis: Beyond stomatal conductance. Plant Physiol. 2021, 186, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Müller, A.; Nothman, J.; Louppe, G.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar] [CrossRef]
- Sainburg, T.; McInnes, L.; Gentner, T.Q. Parametric UMAP Embeddings for Representation and Semisupervised Learning. Neural Comput. 2021, 33, 1–27. [Google Scholar] [CrossRef]
- MacQueen, J. Some methods for classification and analysis of multivariate observations. In Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability; University of California Press: Bakerley, CA, USA, 1965; Volume 1, pp. 281–297. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Experiments | Greenhouse Experiment | Total | |
|---|---|---|---|
| Initial measurements | 120 | 108 | 228 |
| Week 1 | 80 | 108 | 188 |
| Week 2 | 199 | 216 | 415 |
| Week 3 | 159 | 216 | 375 |
| Week 4 | 158 | 216 | 374 |
| Total | 716 | 864 | 1580 |
| Field Experiments | Greenhouse Experiment | Total | |
|---|---|---|---|
| Initial measurements | 8/120 (6.7%) | 6/108 (5.6%) | 14/228 |
| Week 1 | 26/80 (32.5%) | 41/108 (38.0%) | 67/188 |
| Week 2 | 91/199 (45.7%) | 50/216 (23.1%) | 141/415 |
| Week 3 | 121/159 (76.1%) | 83/216 (38.4%) | 204/375 |
| Week 4 | 115/158 (72.8%) | 123/216 (56.9%) | 238/374 |
| Total | 361/716 | 303/864 | 664/1580 |
| Field Experiments | Greenhouse Experiment | |||
|---|---|---|---|---|
| “Anomalies” Type 1 | “Anomalies” Type 2 | “Anomalies” Type 1 | “Anomalies” Type 2 | |
| Initial measurements | 8/120 (6.7%) | 0/108 (0%) | 0/120 (6.7%) | 6/108 (0%) |
| Week 1 | 23/80 (28.8%) | 3/80 (3.8%) | 6/108 (5.6%) | 35/108 (32.4%) |
| Week 2 | 80/199 (40.2%) | 11/199 (5.5%) | 0/216 (0%) | 50/216 (23.1%) |
| Week 3 | 121/159 (76.1%) | 0/159 (0%) | 1/216 (0.5%) | 82/216 (37.9%) |
| Week 4 | 113/158 (71.5%) | 2/158 (1.3%) | 1/216 (0.5%) | 122/216 (56.5%) |
| Total | 345/716 | 16/716 | 8/864 | 295/864 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, N.T. Anomaly Detection Utilizing One-Class Classification—A Machine Learning Approach for the Analysis of Plant Fast Fluorescence Kinetics. Stresses 2024, 4, 773-786. https://doi.org/10.3390/stresses4040051
Tran NT. Anomaly Detection Utilizing One-Class Classification—A Machine Learning Approach for the Analysis of Plant Fast Fluorescence Kinetics. Stresses. 2024; 4(4):773-786. https://doi.org/10.3390/stresses4040051
Chicago/Turabian StyleTran, Nam Trung. 2024. "Anomaly Detection Utilizing One-Class Classification—A Machine Learning Approach for the Analysis of Plant Fast Fluorescence Kinetics" Stresses 4, no. 4: 773-786. https://doi.org/10.3390/stresses4040051
APA StyleTran, N. T. (2024). Anomaly Detection Utilizing One-Class Classification—A Machine Learning Approach for the Analysis of Plant Fast Fluorescence Kinetics. Stresses, 4(4), 773-786. https://doi.org/10.3390/stresses4040051