
Stomatal and Non-Stomatal Leaf Responses during Two Sequential Water Stress Cycles in Young Coffea canephora Plants


 , , , ,
, , , ,  ,
,  ,
,  and
and 
Abstract

1. Introduction
2. Results
2.1. Soil and Leaf Water Potentials during Two Subsequent Water Stress Cycles
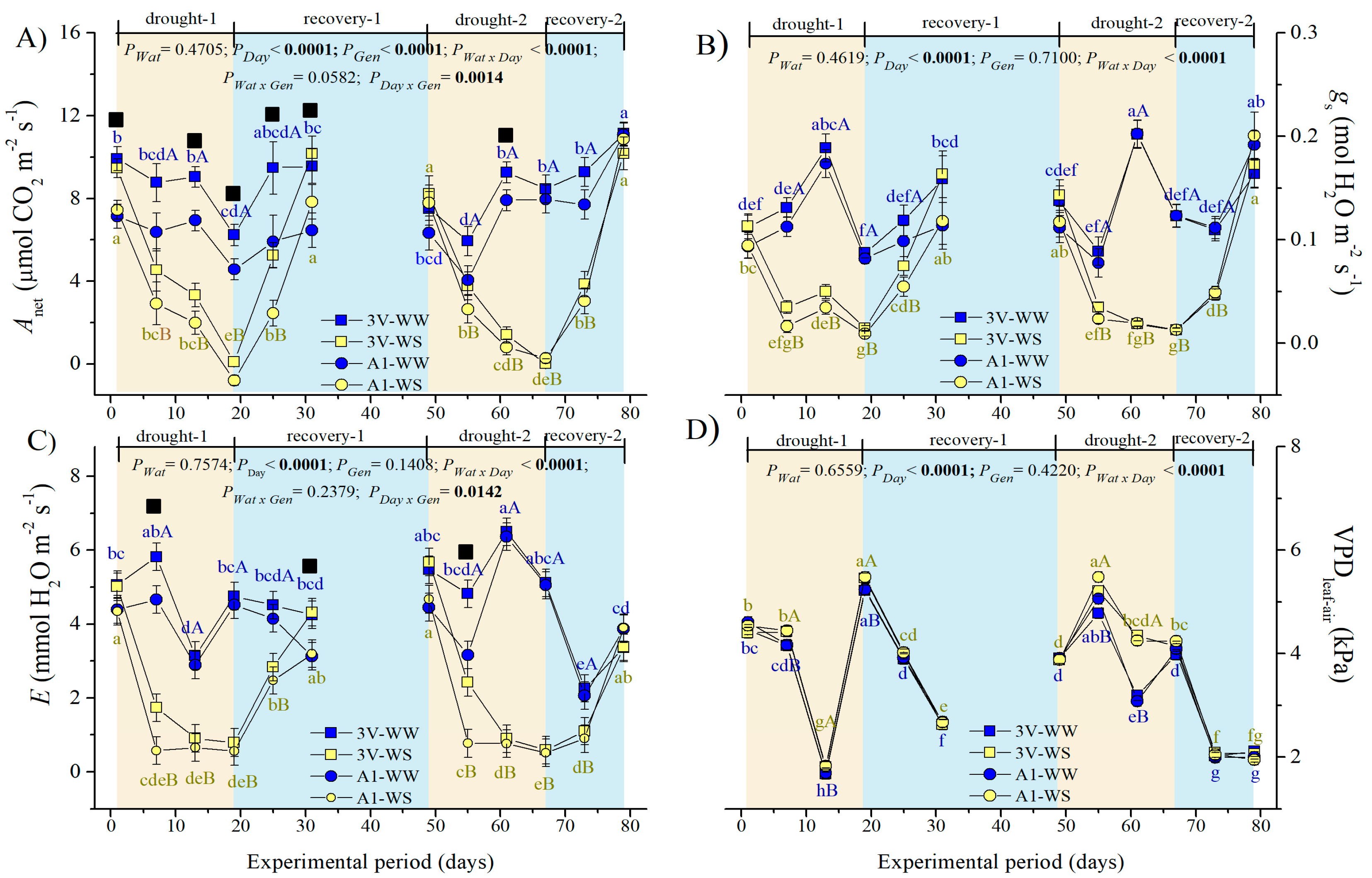
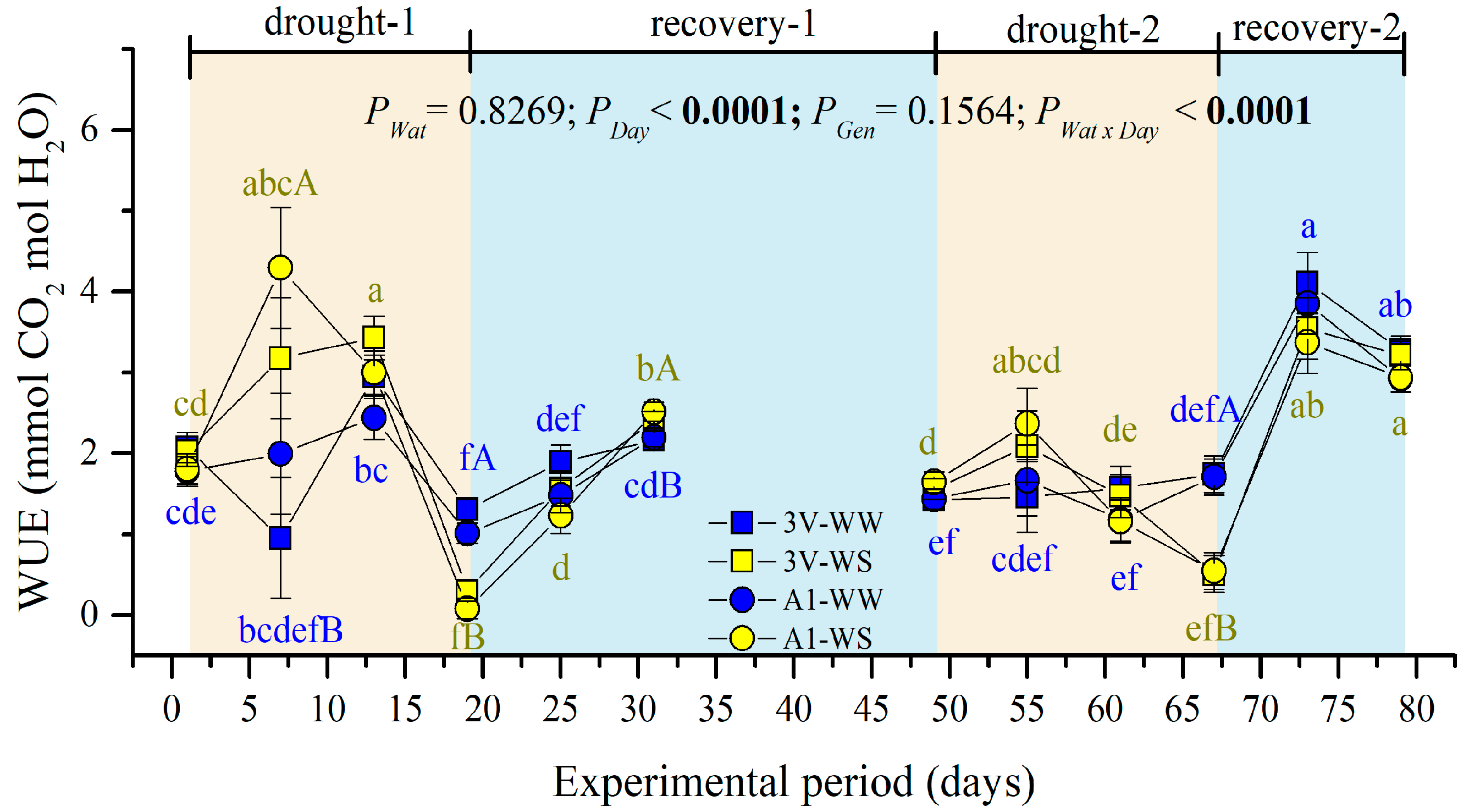
2.2. Leaf Gas Exchanges during Two Subsequent Water Stress Cycles
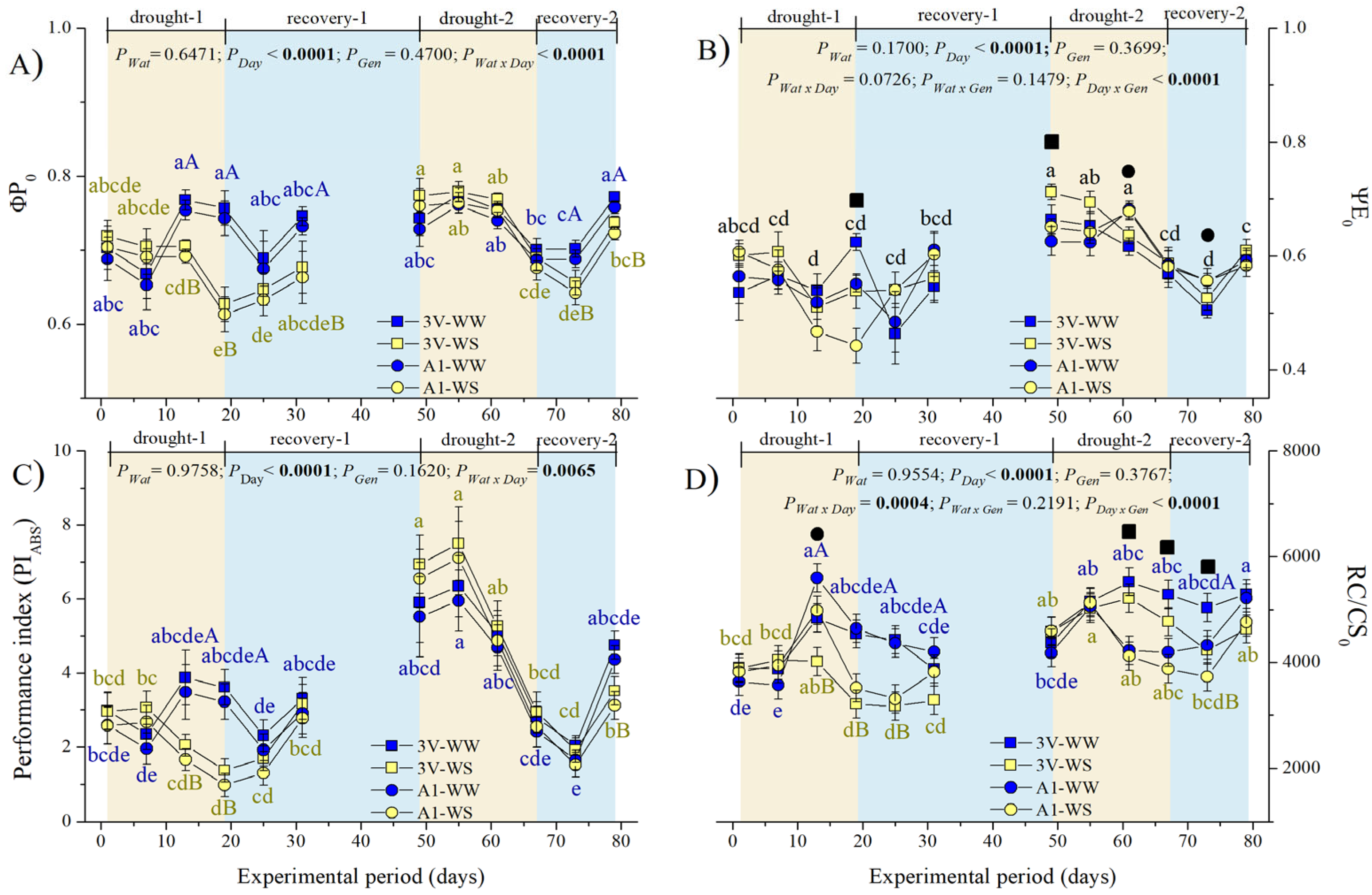
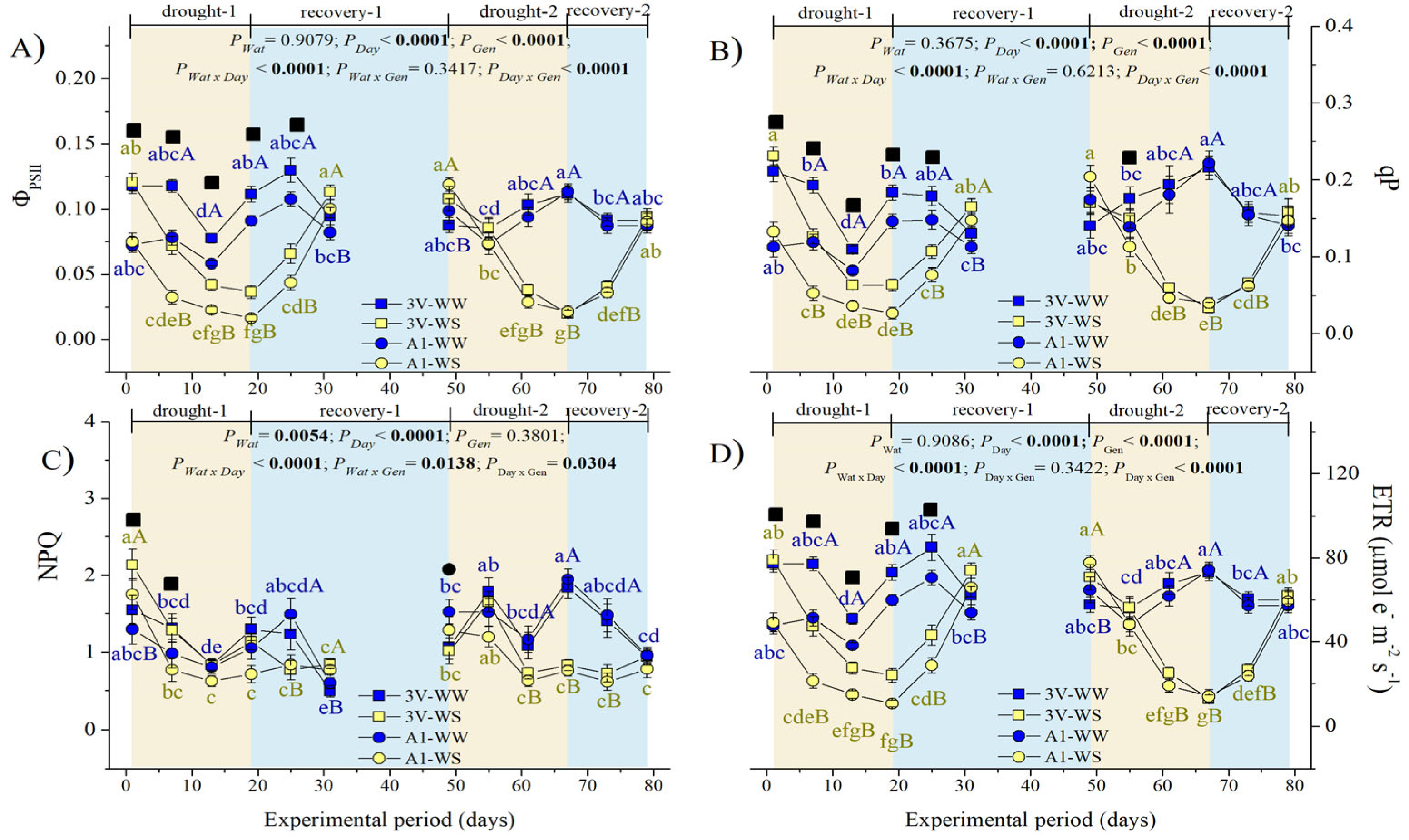
2.3. Leaf Chlorophyll a Fluorescence during Two Subsequent Water Stress Cycles
2.3.1. Dynamics of Non-Modulated Fluorescence
2.3.2. Dynamics of Modulated Chlorophyll a Fluorescence
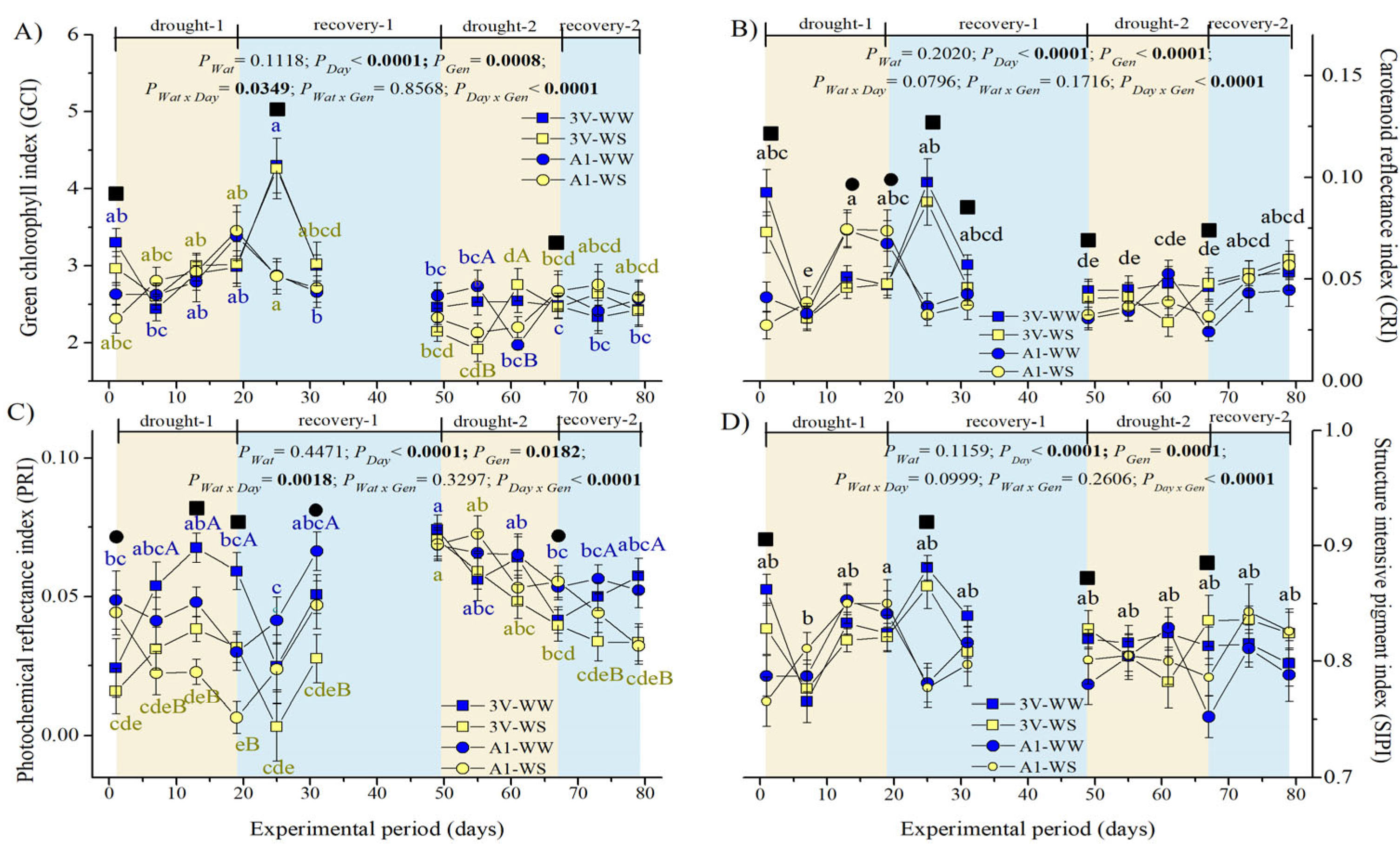
2.4. Spectral Reflectance Indices of Leaf Adaxial Surface during Two Subsequent Water Stress Cycles
2.5. Leaf Anatomy after Two Drought Cycles
3. Discussion
4. Materials and Methods
4.1. Experimental Conditions
4.1.1. Micrometeorological Measurements
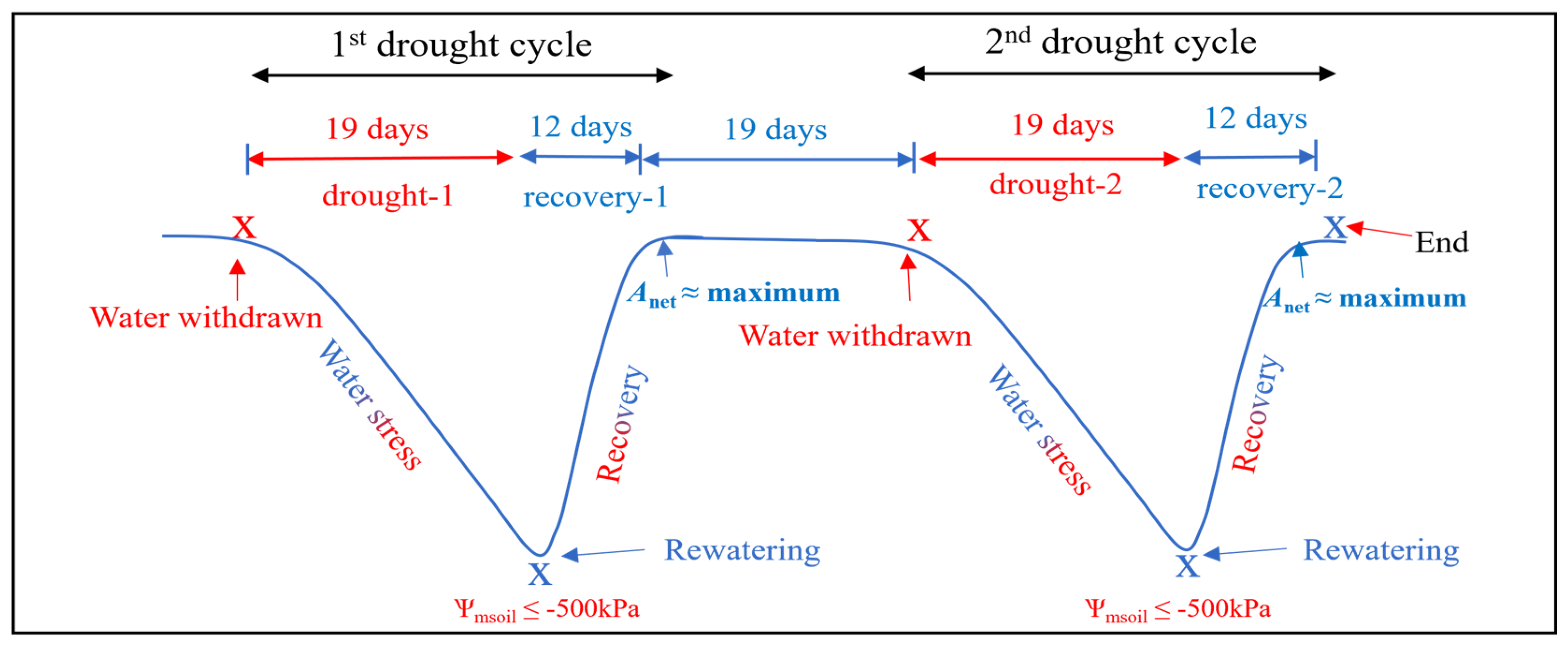
4.1.2. Plant Watering Conditions and Water Deficit Implementation
4.2. Leaf Water Potential Measurements
4.3. Leaf Gas Exchange Measurements
4.4. Chlorophyll a Fluorescence Evaluation
4.4.1. Non-Modulated Chlorophyll a Fluorescence Evaluation
4.4.2. Modulated Chlorophyll a Fluorescence Evaluation
4.5. Leaf Spectral Indices
4.6. Leaf Anatomy
4.7. Statistical Analysis and Experimental Design
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Davis, A.P.; Rakotonasolo, F. Six new species of coffee (Coffea) from northern Madagascar. Kew Bull. 2021, 76, 497–511. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Filho, J.A.M.; Rodrigues, W.P.; Baroni, D.F.; Pireda, S.; Campbell, G.; de Souza, G.A.R.; Filho, A.C.V.; Arantes, S.D.; Arantes, L.d.O.; da Cunha, M.; et al. Linking root and stem hydraulic traits to leaf physiological parameters in Coffea canephora clones with contrasting drought tolerance. J. Plant Physiol. 2021, 258, 153355. [Google Scholar] [CrossRef]
- CONAB. Clima Mais Favorável e Bienalidade Positiva Apontam Produção Estimada em 58,08 milhões de Sacas de Café. 2024. Available online: https://www.conab.gov.br/ultimas-noticias/5362-clima-mais-favoravel-e-bienalidade-positiva-apontam-producao-estimada-em-58-08-milhoes-de-sacas-de-cafe (accessed on 25 July 2024).
- Venancio, L.P.; Filgueiras, R.; Mantovani, E.C.; Amaral, C.H.D.; da Cunha, F.F.; Silva, F.C.d.S.; Althoff, D.; dos Santos, R.A.; Cavatte, P.C. Impact of drought associated with high temperatures on Coffea canephora plantations: A case study in Espírito Santo State, Brazil. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ingrao, C.; Strippoli, R.; Lagioia, G.; Huisingh, D. Water scarcity in agriculture: An overview of causes, impacts and approaches for reducing the risks. Heliyon 2023, 9, e18507. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Grandis, A.; Arenque, B.C.; Buckeridge, M.S. Impacts of climate changes on crop physiology and food quality. Food Res. Int. 2010, 43, 1814–1823. [Google Scholar] [CrossRef]
- Silva, P.E.M.; Cavatte, P.C.; Morais, L.E.; Medina, E.F.; DaMatta, F.M. The functional divergence of biomass partitioning, carbon gain and water use in Coffea canephora in response to the water supply: Implications for breeding aimed at improving drought tolerance. Environ. Exp. Bot. 2013, 87, 49–57. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Manavalan, L.P.; Guttikonda, S.K.; Tran, L.S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Xue, W.; Xiong, L.; Yu, X.; Luo, L.; Cui, K.; Jin, D.; Xing, Y.; Zhang, Q. Genetic basis of drought resistance at reproductive stage in rice: Separation of drought tolerance from drought avoidance. Genetics 2006, 172, 1213–1228. [Google Scholar] [CrossRef]
- Tardieu, F. Plant response to environmental conditions: Assessing potential production, water demand, and negative effects of water deficit. Front. Physiol. 2013, 4, 35373. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheats in a mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Hammer, G.L.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paszkiewicz, S.; Cooper, M. Can changes in canopy and/or root system architecture explain historical maize yield trends in the US corn belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Steele, K.A.; Price, A.H.; Witcombe, J.R.; Shrestha, R.; Singh, B.N.; Gibbons, J.M.; Virk, D.S. QTLs associated with root traits increase yield in upland rice when transferred through marker-assisted selection. Theor. Appl. Genet. 2012, 126, 101–108. [Google Scholar] [CrossRef] [PubMed]
- De Lima, V.J.; Júnior, A.T.D.A.; Kamphorst, S.H.; Bispo, R.B.; Leite, J.T.; Santos, T.d.O.; Schmitt, K.F.M.; Chaves, M.M.; de Oliveira, U.A.; Santos, P.H.A.D.; et al. Combined dominance and additive gene effects in trait inheritance of drought-stressed and full irrigated popcorn. Agronomy 2019, 9, 782. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Knipfer, T.; Fricke, W.; McElrone, A.J. Aquaporins and root water uptake. In Plant Aquaporins. from Signaling and Communication in Plants; Chaumont, F., Tyerman, S.D., Eds.; Springer International Publishing: New York, NY, USA, 2017; Volume 10, pp. 133–153. [Google Scholar]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef]
- Ramalho, J.; Chaves, M.M. Drought effects on plant water relations and carbon gain in two lines of Lupinus albus L. Eur. J. Agron. 1992, 1, 271–280. [Google Scholar] [CrossRef]
- Ennajeh, M.; Vadel, A.M.; Cochard, H.; Khemira, H. Comparative impacts of water stress on the leaf anatomy of a drought-resistant and a drought-sensitive olive cultivar. J. Hortic. Sci. Biotechnol. 2010, 85, 289–294. [Google Scholar] [CrossRef]
- Martins, S.C.V.; Sanglard, M.L.; Morais, L.E.; Menezes-Silva, P.E.; Mauri, R.; Avila, R.T.; Vital, C.E.; Cardoso, A.A.; DaMatta, F.M. How do coffee trees deal with severe natural droughts? An analysis of hydraulic, diffusive and biochemical components at the leaf level. Trees 2019, 33, 1679–1693. [Google Scholar] [CrossRef]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the impact of drought in Coffea genotypes: Transcriptomic analysis supports a common high resilience to moderate water deficit but a genotype dependent sensitivity to severe water deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Change 2019, 152, 167–178. [Google Scholar] [CrossRef]
- Martins, S.C.V.; Galmés, J.; Cavatte, P.C.; Pereira, L.F.; Ventrella, M.C.; DaMatta, F.M. Understanding the low photosynthetic rates of sun and shade coffee leaves: Bridging the gap on the relative roles of hydraulic, diffusive and biochemical constraints to photosynthesis. PLoS ONE 2014, 9, e95571. [Google Scholar] [CrossRef]
- Peloso, A.F.; Tatagiba, S.D.; Reis, E.F.; Pezzopane, J.E.M.; Amaral, F.F.T. Photosynthetic limitations in leaves of Arabic coffee promoted by the water deficit. Coffee Sci. 2017, 12, 389–399. [Google Scholar] [CrossRef]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; et al. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiol. 2021, 41, 708–727. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.C.; Junior, W.Q.R.; Ramos, M.L.G.; Rocha, O.C.; Veiga, A.D.; Silva, N.H.; Brasileiro, L.d.O.; Santana, C.C.; Soares, G.F.; Malaquias, J.V.; et al. Physiological changes of arabica coffee under different intensities and durations of water stress in the Brazilian Cerrado. Plants 2022, 11, 2198. [Google Scholar] [CrossRef] [PubMed]
- Melo, E.F.; Fernandes-Brum, C.N.; Pereira, F.J.; de Castro, E.M.; Chalfun-Júnior, A. Anatomic and physiological modifications in seedlings of Coffea arabica cultivar Siriema under drought conditions. Cienc. Agrotec. 2014, 38, 25–33. [Google Scholar] [CrossRef]
- Rodrigues, A.P.; Pais, I.P.; Leitão, A.E.; Dubberstein, D.; Lidon, F.C.; Marques, I.; Semedo, J.N.; Rakocevic, M.; Scotti-Campos, P.; Campostrini, E.; et al. Uncovering the wide protective responses in Coffea spp. leaves to single and superimposed exposure of warming and severe water deficit. Front. Plant Sci. 2024, 14, 1320552. [Google Scholar] [CrossRef]
- Beyel, V.; Bruggemann, W. Differential inhibition of photosynthesis during pre-flowering drought stress in Sorghum bicolor gen-otypes with different senescence traits. Physiol. Plant. 2005, 124, 249–259. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S.; Wang, Y.L.; Han, G.X.; Li, Y.J. Changes in chlorophyll fluorescence in maize plants with imposed rapid de-hydration at different leaf ages. J. Plant Growth Regul. 2008, 27, 83–92. [Google Scholar] [CrossRef]
- Guóth, A.; Tari, I.; Gallé, A.; Csiszár, J.; Pécsváradi, A.; Cseuz, L.; Erdei, L. Comparison of the drought stress responses of tolerant and sensitive wheat cultivars during grain filling: Changes in flag leaf photosynthetic activity, ABA levels, and grain yield. J. Plant Growth Regul. 2009, 28, 167–176. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Rodrigues, A.P.; Lidon, F.C.; Marques, L.M.C.; Leitão, A.E.; Fortunato, A.S.; Pais, I.P.; Silva, M.J.; Scotti-Campos, P.; Lopes, A.; et al. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp. PLoS ONE 2018, 13, e0198694. [Google Scholar] [CrossRef] [PubMed]
- Scotti-Campos, P.; Pais, I.P.; Ribeiro-Barros, A.I.; Martins, L.D.; Tomaz, M.A.; Rodrigues, W.P.; Campostrini, E.; Semedo, J.N.; Fortunato, A.S.; Martins, M.Q.; et al. Lipid profile adjustments may contribute to warming acclimation and to heat impact mitigation by elevated [CO2] in Coffea spp. Environ. Exp. Bot. 2019, 167, 103856. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Jorge, T.; Osorio, S.; Pott, D.M.; Lidon, F.C.; DaMatta, F.M.; Marques, I.; Ribeiro-Barros, A.I.; Ramalho, J.C.; António, C. Primary metabolite profile changes in Coffea spp. promoted by single and combined exposure to drought and elevated CO2 concentration. Metabolites 2021, 11, 427. [Google Scholar] [CrossRef]
- Marques, I.; Gouveia, D.; Gaillard, J.-C.; Martins, S.; Semedo, M.C.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Armengaud, J.; Ramalho, J.C. Next-generation proteomics reveals a greater antioxidative response to drought in Coffea arabica than in Coffea canephora. Agronomy 2022, 12, 148. [Google Scholar] [CrossRef]
- Driesen, E.; de Proft, M.; Saeys, W. Drought stress triggers alterations of adaxial and abaxial stomatal development in basil leaves increasing water-use efficiency. Hortic. Res. 2023, 10, uhad075. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, J.; Bertin, N.; Bidel, L.P.R.; Urban, L. A user’s view of the parameters derived from the induction curves of maximal chlorophyll a fluorescence: Perspectives for analyzing stress. Front. Plant Sci. 2016, 7, 1679. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Glob. Change Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef]
- Misra, A.N.; Misra, M.; Singh, R. Chlorophyll fluorescence in plant biology. In Biophysics; Misra, A.N., Ed.; Intech: Shanghai, China, 2012; Volume 7, pp. 171–192. [Google Scholar]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Yin, Z.; Meng, F.; Song, H.; He, X.; Xu, X.; Yu, D. Mapping quantitative trait loci associated with chlorophyll a fluorescence parameters in soybean (Glycine max (L.) Merr.). Planta 2010, 231, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, W.P.; Vieira, H.D.; Campostrini, E.; Figueiredo, F.A.M.M.A.; Ferraz, T.M.; Partelli, F.L.; Ramalho, J.C. Physiological aspects, growth and yield of Coffea spp. in areas of high altitude. Aust. J. Crop Sci. 2016, 10, 666–674. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Reboredo, F.H.; Partelli, F.L.; et al. Protective response mechanisms to heat stress in interaction with high [CO2] conditions in Coffea spp. Front. Plant Sci. 2016, 7, 947. [Google Scholar] [CrossRef] [PubMed]
- Dubberstein, D.; Lidon, F.C.; Rodrigues, A.P.; Semedo, J.N.; Marques, I.; Rodrigues, W.P.; Gouveia, D.; Armengaud, J.; Semedo, M.C.; Martins, S.; et al. Resilient and sensitive key points of the photosynthetic machinery of Coffea spp. to the single and superimposed exposure to severe drought and heat stresses. Front. Plant Sci. 2020, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Sukhova, E.; Yudina, L.; Gromova, E.; Ryabkova, A.; Vodeneev, V.; Sukhov, V. Influence of local burning on difference reflectance indices based on 400–700 nm wavelengths in leaves of pea seedlings. Plants 2021, 10, 878. [Google Scholar] [CrossRef]
- Sun, P.; Grignetti, A.; Liu, S.; Casacchia, R.; Salvatori, R.; Pietrini, F.; Loreto, F.; Centritto, M. Associated changes in physiological parameters and spectral reflectance indices in olive (Olea europaea L.) leaves in response to different levels of water stress. Int. J. Remote Sens. 2008, 29, 1725–1743. [Google Scholar] [CrossRef]
- Kior, A.; Sukhov, V.; Sukhova, E. Application of reflectance indices for remote sensing of plants and revealing actions of stressors. Photonics 2021, 8, 582. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.; Ávila, R.T.; Morais, L.E.; Martins, S.C.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R.; et al. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef]
- Marques, I.; Fernandes, I.; Paulo, O.S.; Batista, D.; Lidon, F.C.; Rodrigues, A.P.; Partelli, F.L.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Ramalho, J.C. Transcriptomic analyses reveal that Coffea arabica and Coffea canephora have more complex responses under combined heat and drought than under individual stressors. Int. J. Mol. Sci. 2024, 25, 7995. [Google Scholar] [CrossRef]
- Galle, A.; Florez-Sarasa, I.; El Aououad, H.; Flexas, J. The Mediterranean evergreen Quercus ilex and the semi-deciduous Cistus albidus differ in their leaf gas exchange regulation and acclimation to repeated drought and re-watering cycles. J. Exp. Bot. 2011, 62, 5207–5216. [Google Scholar] [CrossRef] [PubMed]
- Rakočević, M.; Costes, E.; Campostrini, E.; Ramalho, J.C.; Ribeiro, R.V. Drought responses in Coffea arabica as affected by genotype and phenophase. II—Photosynthesis at leaf and plant scales. Exp. Agric. 2024, 60, e7. [Google Scholar] [CrossRef]
- Perrone, I.; Pagliarani, C.; Lovisolo, C.; Chitarra, W.; Roman, F.; Schubert, A. Recovery from water stress affects grape leaf petiole transcriptome. Planta 2012, 235, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful ‘memories’ of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Iwasaki, M.; Paszkowski, J. Epigenetic memory in plants. EMBO J. 2014, 33, 1–12. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Munné-Bosch, S. Stress memory and the inevitable effects of drought: A physiological perspective. Front. Plant Sci. 2016, 7, 143. [Google Scholar] [CrossRef]
- Rakočević, M.; Matsunaga, F.T.; Pazianotto, R.A.A.; Ramalho, J.C.; Costes, E.; Ribeiro, R.V. Drought responses in Coffea arabica as affected by genotype and phenophase. I—Leaf distribution and branching. Exp. Agric. 2024, 60, e7. [Google Scholar] [CrossRef]
- Tyree, M.T.; Sperry, J.S. Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Answers from a model. Plant Physiol. 1988, 88, 574–580. [Google Scholar] [CrossRef]
- Sperry, J.S.; Adler, F.R.; Campbell, G.S.; Comstock, J.P. Limitation of plant water use by rhizosphere and xylem conductance: Results from a model. Plant Cell Environ. 1998, 21, 347–359. [Google Scholar] [CrossRef]
- Whitehead, D. Regulation of stomatal conductance and transpiration in forest canopies. Tree Physiol. 1998, 18, 633–644. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Niu, Y.; Zhang, H.; Han, W.; Li, G.; Tang, J.; Peng, X. Maize canopy temperature extracted from UAV thermal and RGB imagery and its application in water stress monitoring. Front. Plant Sci. 2019, 10, 1270. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Ribas-Carbo, M.; Bota, J.; Galmés, J.; Henkle, M.; Martinez-Canellas, S.; Medrano, H. Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol. 2006, 172, 73–82. [Google Scholar] [CrossRef]
- Aliche, E.B.; Prusova-Bourke, A.; Ruiz-Sanchez, M.; Oortwijn, M.; Gerkema, E.; van As, H.; Visser, R.G.F.; van der Linden, C.G. Morphological and physiological responses of the potato stem transport tissues to dehydration stress. Planta 2020, 251, 45. [Google Scholar] [CrossRef]
- Aro, E.-M.; Virgin, I.; Andersson, B. Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta Bioenerg. 1993, 1143, 113–134. [Google Scholar] [CrossRef]
- Fantuzzi, A.; Allgöwer, F.; Baker, H.; McGuire, G.; Te, W.K.; Gamiz-Hernandez, A.P.; Kaila, V.R.I.; Rutherford, A.W. Bicarbonate-controlled reduction of oxygen by the QA semiquinone in photosystem II in membranes. Proc. Natl. Acad. Sci. USA 2022, 119, e2116063119. [Google Scholar] [CrossRef]
- Guidi, L.; Lo Piccolo, E.; Landi, M. Chlorophyll fluorescence, photoinhibition and abiotic stress: Does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci. 2019, 10, 174. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Koh, S.C.; Cohu, C.; Muller, O.; Stewart, J.; Adams, W., III. Non-photochemical fluorescence quenching in contrasting plant species and environments. In Non-Photochemical Quenching and Energy Dissipation in Plants, Algae and Cyanobacteria; Demmig-Adams, B., Garab, G., Adams, W., III, Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 40, pp. 531–552. [Google Scholar]
- Nakamura, Y.; Tsujimoto, K.; Ogawa, T.; Noda, H.M.; Hikosaka, K. Correction of photochemical reflectance index (PRI) by optical indices to predict non-photochemical quenching (NPQ) across various species. Remote Sens. Environ. 2024, 305, 114062. [Google Scholar] [CrossRef]
- Muller-Moulé, P.; Conklin, P.L.; Niyogi, K.K. Ascorbate deficiency can limit violaxanthin de-epoxidase activity in vivo. Plant Physiol. 2002, 128, 970–977. [Google Scholar] [CrossRef]
- Chen, X.; Li, W.; Lu, Q.; Wen, X.; Li, H.; Kuang, T.; Li, Z.; Lu, C. The xanthophyll cycle and antioxidative defense system are enhanced in the wheat hybrid subjected to high light stress. J. Plant Physiol. 2011, 167, 1828–1836. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, Y.; Li, P.; Zhang, F.; Liu, H.; Zheng, G. Drought stress induces oxidative stress and the antioxidant defense system in ascorbate-deficient vtc1 mutants of Arabidopsis thaliana. Acta Physiol. Plant. 2013, 35, 1189–1200. [Google Scholar] [CrossRef]
- Quinlan, R.F.; Shumskaya, M.; Bradbury, L.M.; Beltrán, J.; Ma, C.; Kennelly, E.J.; Wurtzel, E.T. Synergistic interactions between carotene ring hydroxylases drive lutein formation in plant carotenoid biosynthesis. Plant Physiol. 2012, 160, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Sudrajat, D.J.; Siregar, I.Z.; Khumaida, N.; Siregar, U.J.; Mansur, I. Adaptability of white jabon (Anthocephalus cadamba MIQ.) seedling from 12 populations to drought and waterlogging. Agrivita J. Agric. Sci. 2015, 37, 130–143. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Chaves, A.R.; Pinheiro, H.A.; Ducatti, C.; Loureiro, M.E. Drought tolerance of two field-grown clones of Coffea canephora. Plant Sci. 2003, 164, 111–117. [Google Scholar] [CrossRef]
- Pinheiro, H.A.; DaMatta, F.M.; Chaves, A.R.M.; Loureiro, M.E.; Ducatti, C. Drought tolerance is associated with rooting depth and stomatal control of water use in clones of Coffea canephora. Ann. Bot. 2005, 96, 101–108. [Google Scholar] [CrossRef]
- Reddy, K.S.; Sekhar, K.M.; Sreeharsha, R.V.; Reddy, A.R. Hydraulic dynamics and photosynthetic performance facilitate rapid screening of field grown mulberry (Morus spp.) genotypes for drought tolerance. Environ. Exp. Bot. 2019, 157, 320–330. [Google Scholar] [CrossRef]
- Nteve, G.-M.; Kostas, S.; Polidoros, A.N.; Madesis, P.; Nianiou-Obeidat, I. Adaptation mechanisms of olive tree under drought stress: The potential of modern omics approaches. Agriculture 2024, 14, 579. [Google Scholar] [CrossRef]
- Guedes, F.A.d.F.; Nobres, P.; Ferreira, D.C.R.; Menezes-Silva, P.E.; Ribeiro-Alves, M.; Correa, R.L.; DaMatta, F.M.; Alves-Ferreira, M. Transcriptional memory contributes to drought tolerance in coffee (Coffea canephora) plants. Environ. Exp. Bot. 2018, 147, 220–233. [Google Scholar] [CrossRef]
- Rakočević, M.; Baroni, D.F.; de Souza, G.A.R.; Bernado, W.P.; Almeida, C.M.; Matsunaga, F.T.; Rodrigues, W.P.; Ramalho, J.C.; Campostrini, E. Correlating Coffea canephora 3D architecture to plant photosynthesis at a daily scale and vegetative biomass allocation. Tree Physiol. 2023, 43, 556–574. [Google Scholar] [CrossRef]
- Sousa, J.S.; Neves, J.C.L.; Martinez, H.E.P.; Alvarez, V.H.V. Relationship between coffee leaf analysis and soil chemical analysis. Rev. Bras. Cienc. Solo 2018, 42, e0170109. [Google Scholar] [CrossRef]
- Rakočević, M.; Matsunaga, F.T. Variations in leaf growth parameters within the tree structure of adult Coffea arabica in relation to seasonal growth, water availability and air carbon dioxide concentration. Ann. Bot. 2018, 122, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Schölander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap pressure in vascular plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Rakočević, M. Coffee plant architecture. Advances in Botanical Research. In Coffee—A Glimpse into the Future, 114th ed.; DaMatta, F.M., Ramalho, J.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2024. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M. Stress in plants, from daily rhythm to global changes, detected and quantified by the JIP-test. Chim. Nouv. 2001, 75, 3321–3326. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Advances in Photosynthesis and Respiration; Sharkey, T.D., Eaton-Rye, J.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relation between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Krall, J.; Edwards, G. Environmental effects on the relationship between the quantum yields of carbon assimilation and in vivo PSII electron transport in maize. Funct. Plant Biol. 1991, 18, 267–278. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Miloslavina, Y.; Jahns, P.; Holzwarth, A.R. On the relationship between non-photochemical quenching and pho-toprotection of photosystem II. Biochim Biophys Acta. 2012, 1817, 760–769. [Google Scholar] [CrossRef]
- Foyer, C.H.; Ruban, A.V.; Noctor, G. Viewing oxidative stress through the lens of oxidative signalling rather than damage. Biochem. J. 2017, 474, 877–883. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free. Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M. Plant photochemistry, reactive oxygen species, and photoprotection. Photochem 2022, 2, 5–8. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis In Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2007, 75, 272–281. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef]
- Peñuelas, J.; Baret, F.; Filella, I. Semi-empirical indices to assess carotenoids/chlorophyll-a ratio from leaf spectral reflectance. Pho-tosynthetica 1995, 31, 221–230. [Google Scholar]
- Ramalho, J.C.; Rodrigues, A.P.; Semedo, J.N.; Pais, I.P.; Martins, L.D.; Simões-Costa, M.C.; Leitão, A.E.; Fortunato, A.S.; Batista-Santos, P.; Palos, I.M.; et al. Sustained Photosynthetic Performance of Coffea spp. under Long-Term Enhanced [CO2]. PLoS ONE 2013, 8, e82712. [Google Scholar] [CrossRef]
- Neto, B.C.; Silva, F.; Ferreira, T.; Crasque, J.; Arantes, L.; Filho, J.M.; DE Souza, T.; Falqueto, A.; Dousseau-Arantes, S. Responses of wild Piper species to drought and rehydration cycles considering stomatal closure as a marker of the alarm phase. Photosynthetica 2023, 61, 363–376. [Google Scholar] [CrossRef]
- R Core Team. Available online: https://www.r-project.org// (accessed on 14 April 2024).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Anatomy Parameter | Well-Watered (WW) | Water-Stress (WS) | p-Value | ||||
|---|---|---|---|---|---|---|---|
| ‘3V’ | ‘A1’ | ‘3V’ | ‘A1’ | Clone | Water | Interaction | |
| Stomatal density (number stomata mm−2) | 269.5 ± 8.7 aA | 251.5 ± 6.9 bA | 278.0 ± 18.2 aA | 250.7 ± 7.5 bA | 0.0370 | 0.8920 | - |
| Leaf thickness (µm) | 221.7 ± 3.0 bA | 228.6 ± 3.6 aB | 221.7 ± 3.7 bA | 241.3 ± 2.6 aA | <0.0001 | 0.0420 | 0.0500 |
| Thickness of adaxial epidermis (µm) | 21.07 ± 0.6 aA | 20.87 ± 0.5 aA | 20.4 ± 0.4 aA | 21.1 ± 0.6 aA | 0.4810 | 0.6530 | - |
| Thickness of abaxial epidermis (µm) | 14.0 ± 0.4 aA | 13.7 ± 0.5 aA | 13.4 ± 0.3 aA | 13.9 ± 0.3 aA | 0.6020 | 0.4930 | - |
| Thickness of palisade parenchyma (µm) | 54.0 ± 1.6 aA | 48.6 ± 0.9 bA | 46.6 ± 1.4 aB | 46.2 ± 3.0 aA | 0.2980 | 0.0020 | 0.0191 |
| Thickness of spongy parenchyma (µm) | 131.4 ± 3.4 bB | 147.3 ± 3.8 aB | 140.7 ± 3.5 bA | 158.7 ± 3.7aA | <0.0001 | 0.0070 | - |
| Density of xylem vessels (number * 10−5 µm−2) | 172.1 ± 9.2 aA | 190.2 ± 10.5 aA | 188.4 ± 11.1 aA | 182.5 ± 7.7 aA | 0.2430 | 0.8570 | - |
| Medium area of xylem vessel (µm2) | 234.1 ± 9.5 aA | 203.6 ± 11.3 bA | 190.0 ± 11.0 aB | 200.0 ± 9.7 aA | 0.2050 | 0.0370 | 0.0476 |
| Index | Formula | Description | References |
|---|---|---|---|
| Non-Modulated Fluorescence (Pocket PEA) | |||
| ΦP0 | TRo/ABS = [1 − (Fo/Fm)] = Fv/Fm | Maximum quantum yield of primary photochemical reactions | [43,89,90] |
| ΨE0 | ETo/TRo = (1 − VJ) | Probability of electron transfer from QA− to electron transport chain beyond QA | [43,89,90] |
| PIABS | (RC/ABS) · [ΦPo/(1 − ΦPo)] · [Ψo/(1 − Ψo)] | Photosynthetic performance index | [43,89,90] |
| RC/CS0 | (Vj/M0) * (ABS/CS0) | Density of reaction centers capable of QA reduction | [43,89,90] |
| Modulated fluorescence (IRGA LI 6400 XT) | |||
| ΦPSII | (Fm′> − Fs)/Fm′ | Effective quantum yield in PSII photochemistry | [39,91] |
| qP | (Fm′ − Fs)/(Fm′ − Fo′) | Photochemical quenching | [92] |
| NPQ | (Fm − Fm′)/Fm′ | Photo-protective process that removes excess excitation energy within chlorophyll-containing complexes and prevents the likelihood of formation of damaging free radicals. Process is considered adequate under stressful conditions if it can retain an equal fraction of open PSII reaction centers as of non-stress conditions. | [93,94,95,96,97,98] |
| ETR | ΦPSII · PPFD · αlea . f | The linear electron transport rate was calculated using the photochemical efficiency at electron transport of PSII. | [99] |
| Reflectance indexes (CI-710) | |||
| GCI | W554/W677 | Green chlorophyll index | [100] |
| CRI | (1/W510) − (1/W550) | Carotenoid reflectance index | [100] |
| PRI | (W531 − W570)/(W531 + W570) | Photochemical reflectance index | [101] |
| SIPI | (W800 − W445)/(W800 + W680) | Structure intensive pigment index | [102] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baroni, D.F.; de Souza, G.A.R.; Bernado, W.d.P.; Santos, A.R.; Barcellos, L.C.d.S.; Barcelos, L.F.T.; Correia, L.Z.; de Almeida, C.M.; Verdin Filho, A.C.; Rodrigues, W.P.; et al. Stomatal and Non-Stomatal Leaf Responses during Two Sequential Water Stress Cycles in Young Coffea canephora Plants. Stresses 2024, 4, 575-597. https://doi.org/10.3390/stresses4030037
Baroni DF, de Souza GAR, Bernado WdP, Santos AR, Barcellos LCdS, Barcelos LFT, Correia LZ, de Almeida CM, Verdin Filho AC, Rodrigues WP, et al. Stomatal and Non-Stomatal Leaf Responses during Two Sequential Water Stress Cycles in Young Coffea canephora Plants. Stresses. 2024; 4(3):575-597. https://doi.org/10.3390/stresses4030037
Chicago/Turabian StyleBaroni, Danilo F., Guilherme A. R. de Souza, Wallace de P. Bernado, Anne R. Santos, Larissa C. de S. Barcellos, Letícia F. T. Barcelos, Laísa Z. Correia, Claudio M. de Almeida, Abraão C. Verdin Filho, Weverton P. Rodrigues, and et al. 2024. "Stomatal and Non-Stomatal Leaf Responses during Two Sequential Water Stress Cycles in Young Coffea canephora Plants" Stresses 4, no. 3: 575-597. https://doi.org/10.3390/stresses4030037
APA StyleBaroni, D. F., de Souza, G. A. R., Bernado, W. d. P., Santos, A. R., Barcellos, L. C. d. S., Barcelos, L. F. T., Correia, L. Z., de Almeida, C. M., Verdin Filho, A. C., Rodrigues, W. P., Ramalho, J. C., Rakočević, M., & Campostrini, E. (2024). Stomatal and Non-Stomatal Leaf Responses during Two Sequential Water Stress Cycles in Young Coffea canephora Plants. Stresses, 4(3), 575-597. https://doi.org/10.3390/stresses4030037