


Nitric Oxide Modulates Salt Stress Tolerance in Lettuce

 , ,
, ,
Abstract
1. Introduction
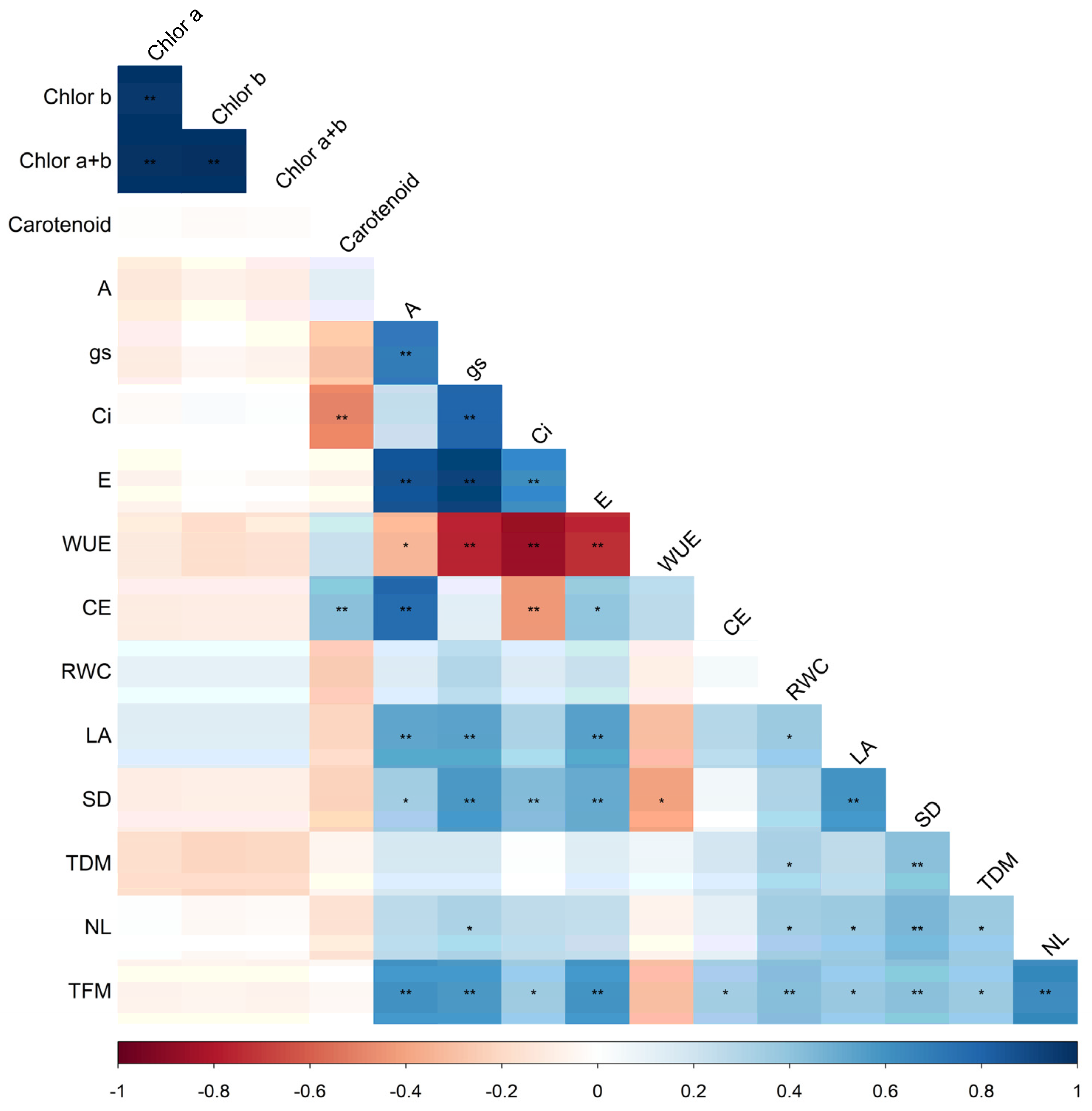
2. Results
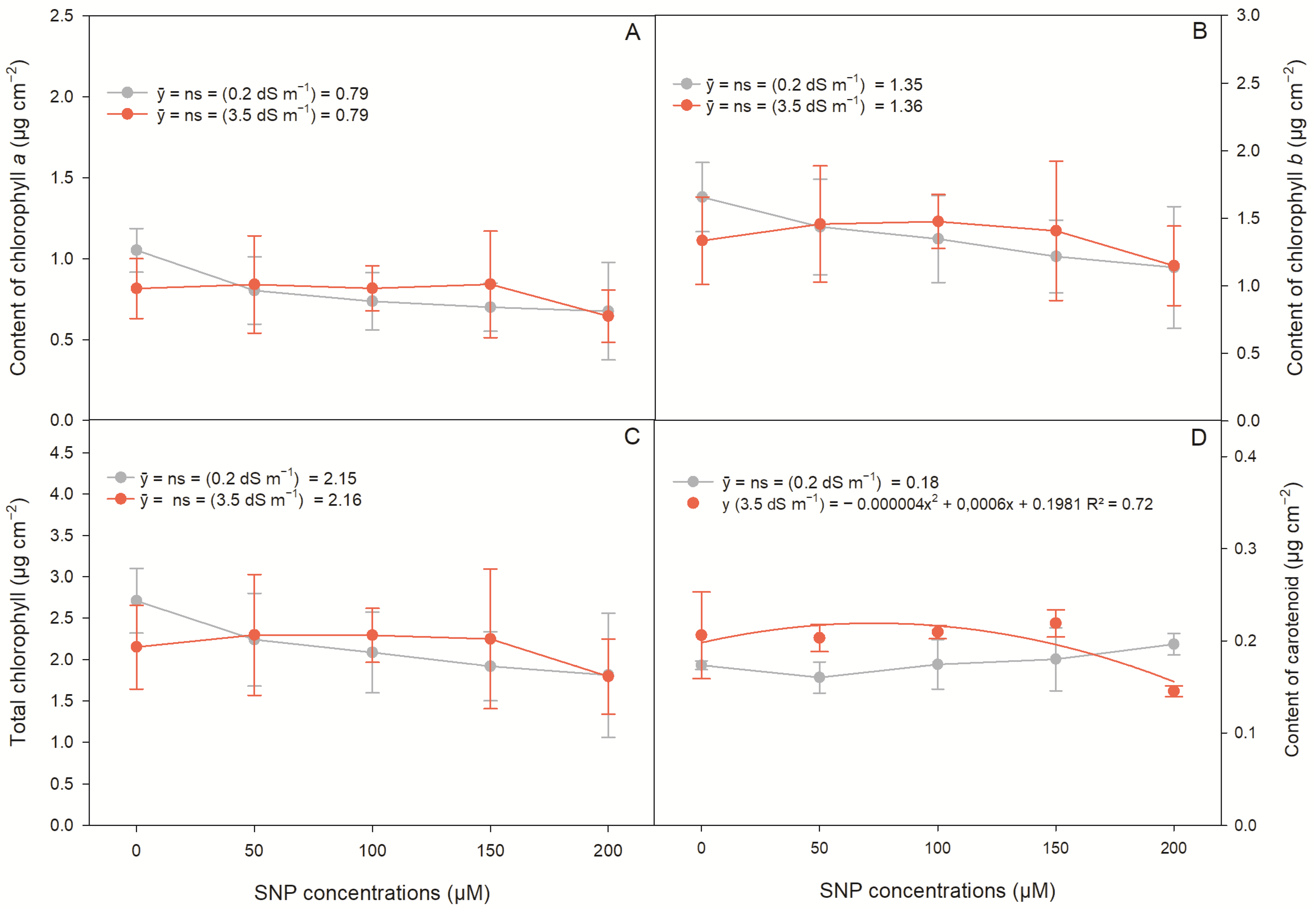
2.1. Photosynthetic Pigments in Lettuce Plants
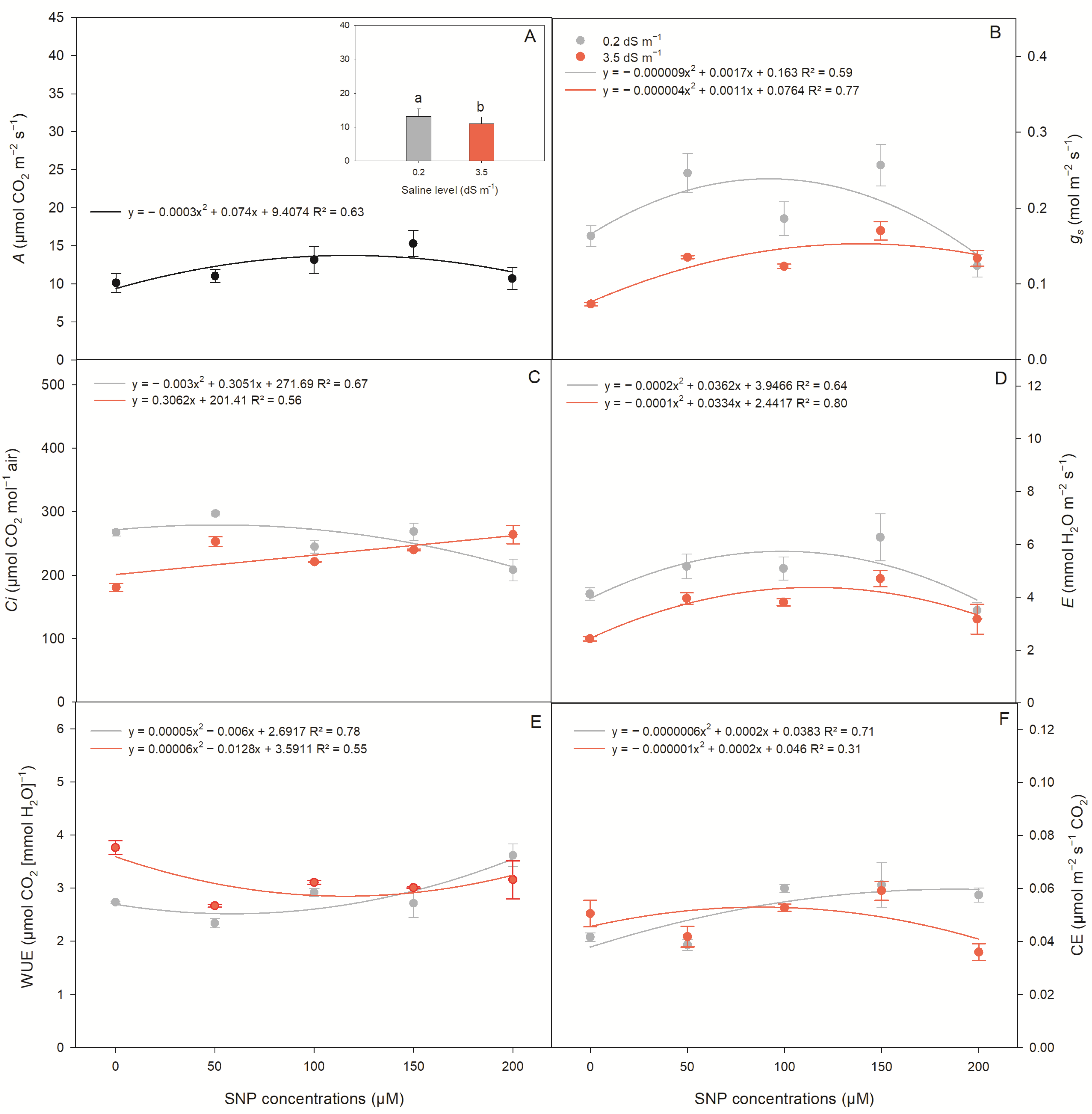
2.2. Gas Exchange in Lettuce Plants
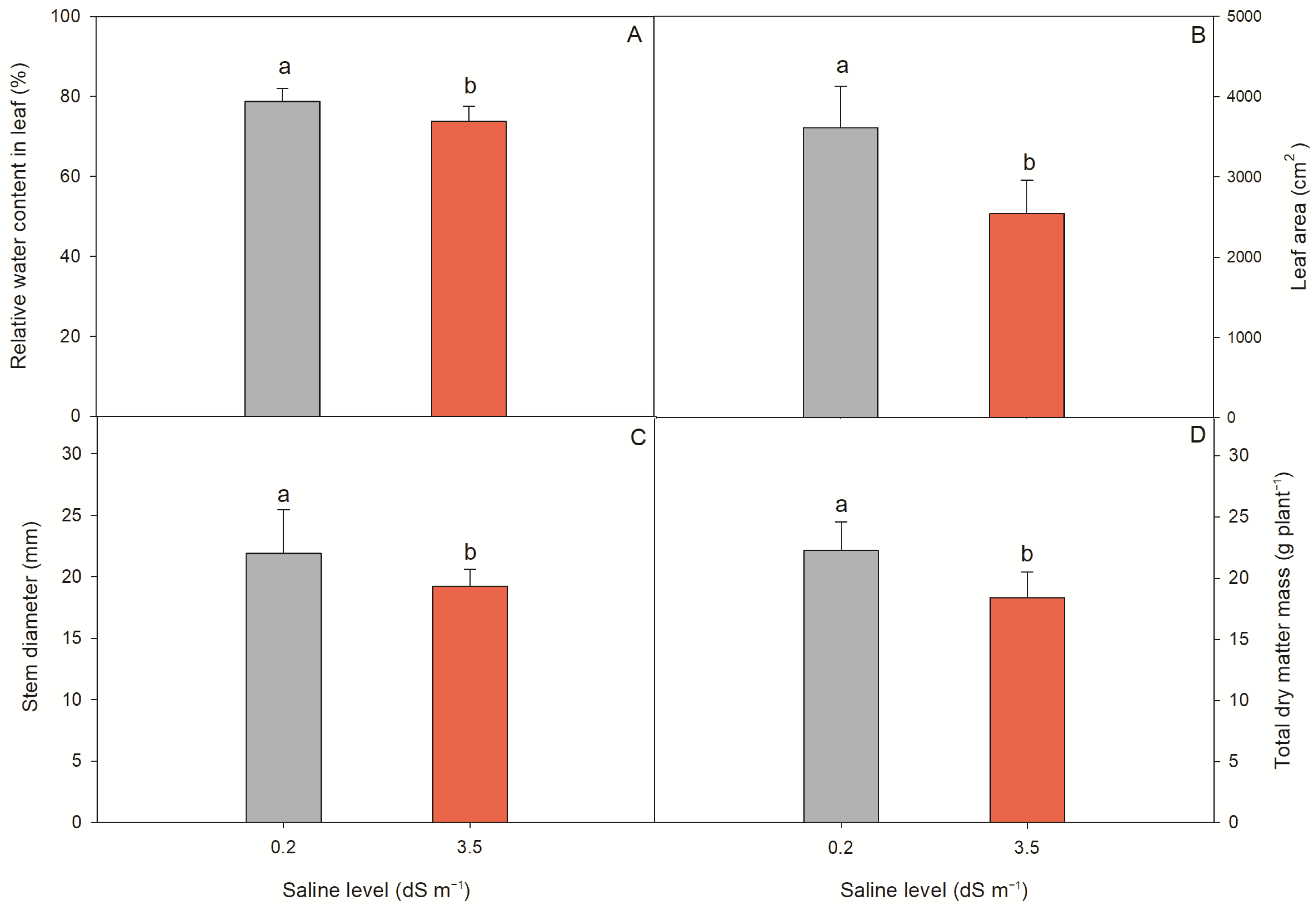
2.3. Morpho-Physiological Parameters of Lettuce Plants
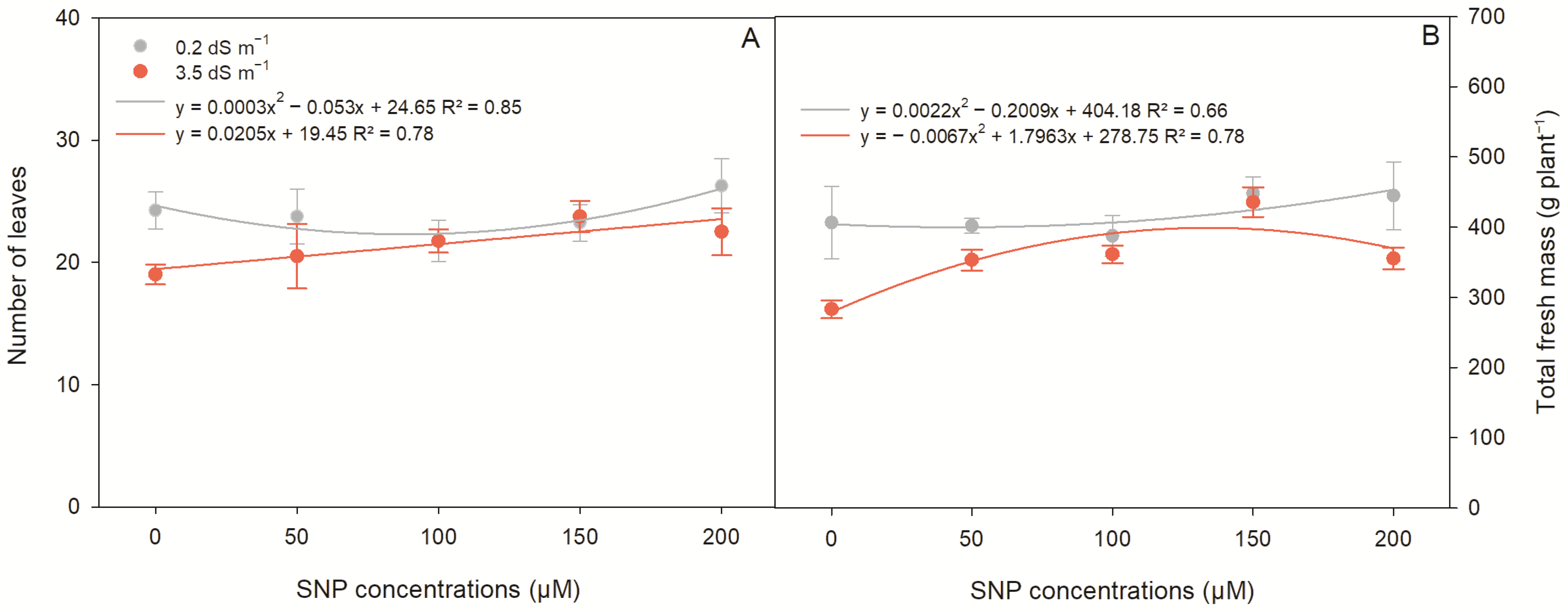
2.4. Productive Parameters of Lettuce Plants
3. Discussion
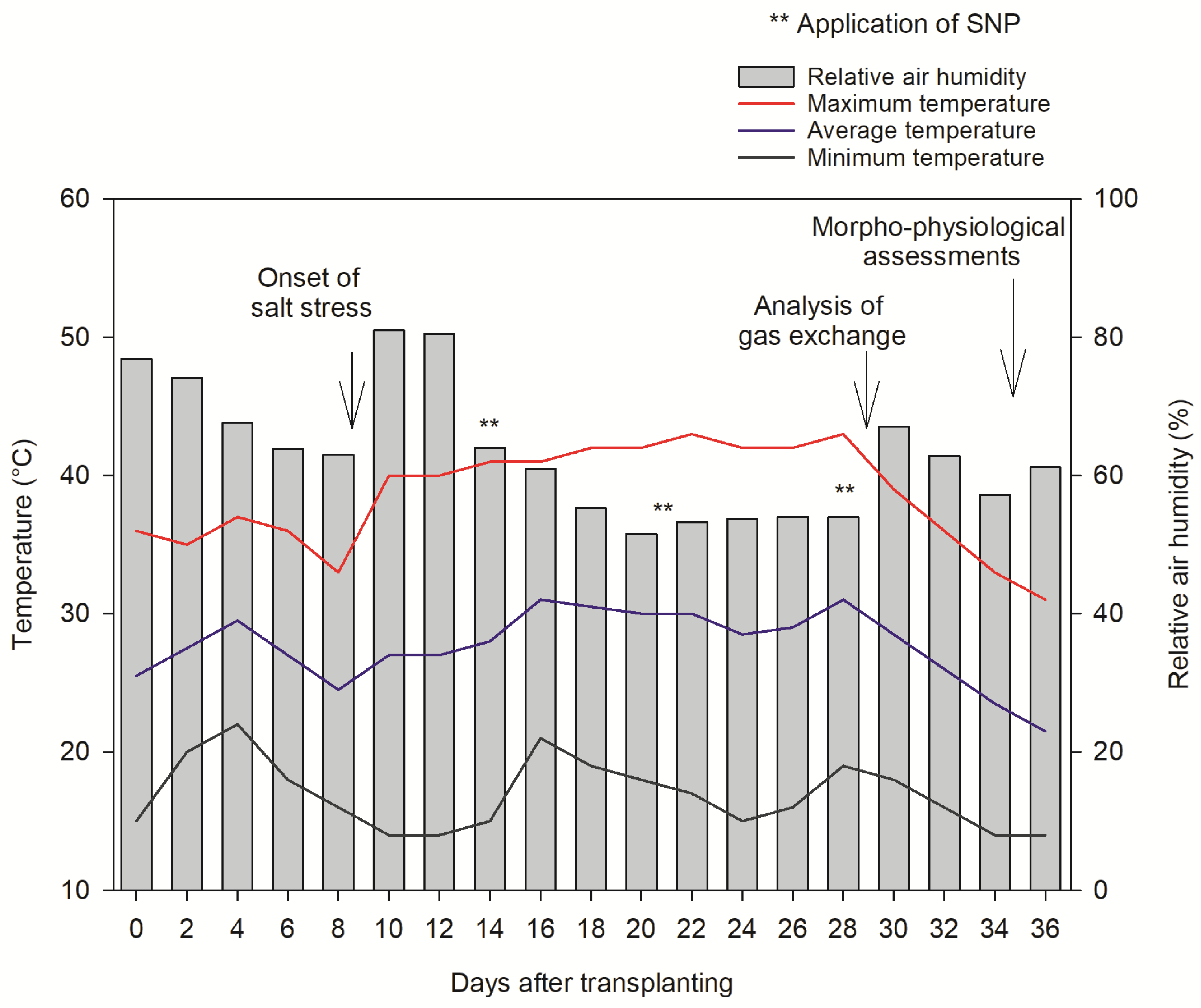
4. Materials and Methods
4.1. Experiment Design
4.2. Crop Management and Fertirrigation
4.3. Parameters Analyzed
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, D.K.; Chaudhari, S.K.; Singh, A. In salt affected soils agroforestry is a promising option. Indian Farming 2014, 63, 19–22. [Google Scholar]
- Farouk, S.; Al-Huqail, A.A. Sustainable biochar and/or melatonin improve salinity tolerance in borage plants by modulating osmotic adjustment, antioxidants, and ion homeostasis. Plants 2022, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.V.; Oliveira, J.A.; Pereira, M.G.; Farnese, F.S. Nitric oxide and phytohormone interactions in the response of Lactuca sativa to salinity stress. Planta 2019, 250, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Nachshon, U. Cropland soil salinization and associated hydrology: Trends, processes and examples. Water 2018, 10, 1030. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, M.; Fu, M.; Qin, J.; Chen, J.; Feng, Y.; Li, G.; Zhai, M.; Zhang, Y.; Ning, T. Water sources effects on soil salinity and biomass production of agriculture and agroforest systems in raised fields. J. Plant Interact. 2020, 15, 303–312. [Google Scholar] [CrossRef]
- Liu, D.; She, D.; Mu, X. Water flow and salt transport in bare saline-sodic soils subjected to evaporation and intermittent irrigation with saline/distilled water. Land. Degrad. Dev. 2019, 30, 1204–1218. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Li, L.; Fu, Q.; Li, Q.; Naeem, M.A.; He, X.; Zhang, Z.; Jacobsen, S.-E. Biochar mitigates combined effects of drought and salinity stress in Quinoa. Agronomy 2020, 10, 912. [Google Scholar] [CrossRef]
- Carillo, P.; Raimondi, G.; Kyriacou, M.C.; Pannico, A.; El-Nakhel, C.; Cirillo, V.; Colla, G.; Pascale, S.; Rouphael, Y. Morpho-physiological and homeostatic adaptive responses triggered by omeprazole enhace lettuce tolerance to salt stress. Sci. Hortic. 2019, 249, 22–30. [Google Scholar]
- Hniličková, H.; Hnilička, F.; Martinkova, J.; Kraus, K. Efects of salt stress on water status, photosynthesis and chlorophyll fuorescence of rocket. Plant Soil. Environ. 2017, 63, 362–367. [Google Scholar] [CrossRef]
- Kim, H.; Jeong, H.; Jeon, J.; Bae, S. Efects of irrigation with saline water on crop growth and yield in greenhouse cultivation. Water 2016, 8, 127. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Adhikari, B.; Olorunwa, O.J.; Wilson, J.C.; Barickman, T.C. Morphological and Physiological Response of Different Lettuce Genotypes to Salt Stress. Stresses 2021, 1, 285–304. [Google Scholar] [CrossRef]
- Cavalcante, D.A.; Leite Júnior, B.R.D.C.; Lima, A.A.; Tribst, M.C.; Cristianini, M. Sanitização de alface americana com água ozonizada para inativação de Escherichia coli O157:H7. Segurança Aliment. Nutr. 2014, 21, 373–378. [Google Scholar] [CrossRef][Green Version]
- Taiz, L.; Zeiger, E. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017; 888p. [Google Scholar]
- Sami, F.; Faizan, M.; Faraz, A.; Siddiqui, H.; Yusuf, M.; Hayat, S. Nitric oxide-mediated integrative alterations in plant metabolism to confer abiotic stress tolerance, NO crosstalk with phytohormones and NO-mediated post translational modifications in modulating diverse plant stress. Nitric Oxide 2018, 73, 22–38. [Google Scholar] [CrossRef]
- Sousa, L.F.; Menezes-Silva, P.E.; Lourenço, L.L.; Galmés, J.; Guimarães, A.C.; Silva, A.F.; Lima, A.P.R.; Henning, L.M.M.; Costa, A.C.; Silva, F.G.; et al. Improving water use efficiency by changing hydraulic and stomatal characteristics in soybean exposed to drought: The involvement of nitric oxide. Physiol. Plant 2019, 168, 576–589. [Google Scholar] [CrossRef]
- Corpas, F.J.; Leterrier, M.; Valderrama, R.; Airaki, M.; Chaki, M.; Palma, J.M.; Barroso, J.B. Nitric oxide imbalance provokes a nitrosative response in plants under abiotic stress. Plant Sci. 2011, 181, 604–611. [Google Scholar] [CrossRef]
- Neil, S.J.; Desikan, R.; Hancock, J.T. Nitric oxide signalling in plants. New Phytol. 2003, 159, 11–35. [Google Scholar] [CrossRef]
- Khurana, A.; Khurana, J.P.; Babbar, S.B. Nitric oxide induces flowering in the Duckweed Lemma aequinoctialis Welw. (Syn. L. paucicostata Hegelm.) under noninductive conditions. J. Plant Growth Regul. 2011, 30, 378–385. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Effect of sodium nitroprusside on physiological and anatomical features of salt-stressed Raphanus sativus. Plant Physiol. Biochem. 2021, 169, 160–170. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Nitrate reductase rather than nitric oxide synthase activity is involved in 24-epibrassinolide-induced nitric oxide synthesis to improve tolerance to iron deficiency in strawberry (Fragaria × annassa) by up regulating the ascorbate-glutathione cycle. Plant Physiol. Biochem. 2020, 151, 486–499. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Huqail, A.A. Sodium nitroprusside application regulates antioxidant capacity, improves phytopharmaceutical production and essential oil yield of marjoram herb under drought. Ind. Crops Prod. 2020, 158, 113034. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Ghamdi, A.A.M. Sodium nitroprusside application enhances drought tolerance in marjoram herb by promoting chlorophyll biosynthesis and enhancing osmotic adjustment capacity. Arab. J. Geosci. 2021, 14, 430. [Google Scholar] [CrossRef]
- Gomes, M.A.C.; Pestana, I.A.; Santa-Catarina, C.; Hauser-Davis, R.A.; Suzuki, M.S. Salinity effects on photosynthetic pigments, proline, biomass and nitric oxide in Salvinia auriculata Aubl. Acta Limnol. Bras. 2017, 29, e9. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z. Exogenous nitric oxide protect garlic plants against oxidative stress induced by salt stress. Plant Stress 2022, 5, 100101. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Khan, E.A.; Misra, A.N. Mitigation effect of exogenous nitric oxide (NO) on some metabolic compounds of maize seedling grown under salt stress. J. Phys. Conf. Ser. 2019, 1294, 052008. [Google Scholar] [CrossRef]
- Alnusairi, G.S.H.; Mazrou, Y.S.A.; Qari, S.H.; Elkelish, A.A.; Soliman, M.H.; Eweis, M.; Abdelaal, K.; El-Samad, G.A.; Ibrahim, M.F.M.; ElNahhas, N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants 2021, 10, 1693. [Google Scholar] [CrossRef] [PubMed]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.I.A.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A.S.; et al. Exogenous Sodium Nitroprusside Mitigates Salt Stress in Lentil (Lens culinaris Medik.) by Affecting the Growth, Yield, and Biochemical Properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef] [PubMed]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. NO to drought-multifunctional role of nitric oxide in plant drought: Do we have all the answers? Plant Sci. 2015, 239, 44–55. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, L.; Wang, X.; Wang, Z.; Zhang, H.; Chen, J.; Liu, X.; Wang, Y.; Li, C. Beneficial effects of exogenous melatonin on overcoming salt stress in sugar beets (Beta vulgaris L.). Plants 2021, 10, 886. [Google Scholar] [CrossRef]
- Ren, J.; Ye, J.; Yin, L.; Li, G.; Deng, X.; Wang, S. Exogenous melatonin improves salt tolerance by mitigating osmotic, ion, and oxidative stresses in maize seedlings. Agronomy 2020, 10, 663. [Google Scholar] [CrossRef]
- Khan, I.; Muhammad, A.; Chattha, M.U.; Skalicky, M.; Chattha, M.B.; Ayub, M.A.; Anwar, M.R.; Soufan, W.; Hassan, M.U.; Rahman, M.A.; et al. Mitigation of salinity-induced oxidative damage, growth, and yield reduction in fine rice by sugarcane press mud application. Front. Plant Sci. 2022, 13, 840900. [Google Scholar] [CrossRef] [PubMed]
- Silveira, N.M.; Frungillo, L.; Marcos, F.C.; Pelegrino, M.T.; Miranda, M.T.; Seabra, A.B.; Salgado, I.; Machado, E.C.; Ribeiro, R.V. Exogenous nitric oxide improves sugarcane growth and photosynthesis under water deficit. Planta 2016, 244, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Jabeen, Z.; Fayyaz, H.A.; Irshad, F.; Hussain, N.; Hassan, M.N.; Li, J.; Rehman, S.; Haider, W.; Yasmin, H.; Mumtaz, S.; et al. Sodium nitroprusside application improves morphological and physiological attributes of soybean (Glycine max L.) under salinity stress. PLoS ONE 2021, 16, e0248207. [Google Scholar] [CrossRef] [PubMed]
- Melo, H.F.; Souza, E.R.; Duarte, H.H.F.; Cunha, J.C.; Santos, H.R.B. Gas exchange and photosynthetic pigments in bell pepper irrigated with saline water. Rev. Bras. Eng. Agricola Ambient. 2017, 21, 38–43. [Google Scholar] [CrossRef]
- Alzahib, R.H.; Migdadi, H.M.; Al Ghamdi, A.A.; Alwahibi, M.S.; Ibrahim, A.A.; Al-Selwey, W.A. Assessment of morpho-physiological, biochemical and antioxidant responses of tomato landraces to salinity stress. Plants 2021, 10, 696. [Google Scholar] [CrossRef]
- Jamil, M.; Rehman, S.U.; Lee, K.J.; Kim, J.M.; Kim, H.; Rha, E.S. Salinity reduced growth PS2 photochemistry and chlorophyll content in radish. Sci. Agric. 2007, 64, 111–118. [Google Scholar] [CrossRef]
- Sarmento, J.D.A.; Morais, P.L.D.; Almeida, M.L.B.; Sousa Neto, O.N.; da Silva Dias, N. Qualidade e conservação da alface cultivada com rejeito da dessalinização. Rev. Caatinga 2014, 27, 90–97. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of photosynthesis during abiotic stress-induced photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; Abd Allah, E.F.; Gucel, S.; Tran, L.-S.P. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef]
- Rahman, A.; Hossain, M.D.S.; Mahmud, J.-A.L.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Manganese-induced salt stress tolerance in rice seedlings: Regulation of ion homeostasis, antioxidant defense and glyoxalase systems. Physiol. Mol. Biol. Plants 2016, 22, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.M.R.; Santos, J.C.C.; Rosa, V.R.; Santos, A.L.F.; Silva, M.A. Tolerance to water deficiency in safflower (Carthamus tinctorius L.) modulated by potassium fertilization. Acta Physiol. Plant 2022, 44, 99. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Souto Filho, L.T.; Laime, E.M.O.; Fernandes, P.D.; Suassuna, J.F.; Silva, R.C. Efeito da salinidade nas trocas gasosas de duas cultivares de cana-de-açúcar. Eng. Ambient. 2014, 11, 67–82. [Google Scholar]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Tabassum, M.A. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. 2015, 22, 4907–4921. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Du, Y.; Hou, Y.J.; Zhao, Y.; Hsu, C.C.; Yuan, F.; Zhu, J.K. Nitric oxide negatively regulates abscisic acid signaling in guard cells by S-nitrosylation of OST1. Proc. Natl. Acad. Sci. USA 2015, 112, 613–618. [Google Scholar] [CrossRef]
- Lakzayi, M.; Sabbagh, E.; Rigi, K.; Keshtehgar, A. Effect of salicylic acid on activities of antioxidant en-zymes, flowering and fruit yield and the role on reduce of drought stress. Int. J. Farming Allied Sci. 2014, 3, 980–987. [Google Scholar]
- Oliveira, F.A.; Oliveira, M.K.T.; Lima, L.A.; Alves, R.C.; Régis, L.R.L.; Santos, S.T. Estresse salino e biorregulador vegetal em feijão caupi. Irriga 2017, 22, 314–329. [Google Scholar] [CrossRef]
- Oliveira, F.A.; Medeiros, J.F.; Oliveira, M.K.T.; Souza, A.A.T.; Fereira, J.A.; Souza, M.S. Interação entre salinidade e bioestimulante na cultura do feijão caupi. Rev. Bras. Eng. Agricola Ambient. 2013, 17, 465–471. [Google Scholar] [CrossRef]
- Paulus, D.; Paulus, E.; Nava, G.A.; Moura, C.A. Crescimento, consumo hídrico e composição mineral de alface cultivada em hidroponia com águas salinas. Rev. Ceres 2012, 59, 110–117. [Google Scholar] [CrossRef]
- Moraes, D.P.; Fernandes, A.L.M.; Dias, N.S.; Cosme, C.R.; Souza Neto, O.N. Rejeito salino e solução nutritiva em alface cultivada em sistema hidropônico. Magistra 2014, 26, 353–360. [Google Scholar]
- Sanz, L.; Albertos, P.; Mateos, I.; Sánchez-Vicente, I.; Lechón, T.; Fernández-Marcos, M.; Lorenzo, O. Nitric oxide (NO) and phytohormones crosstalk during early plant development. J. Exp. Bot. 2015, 66, 2857–2868. [Google Scholar] [CrossRef] [PubMed]
- Silveira, N.M.; Marcos, F.C.C.; Frungillo, L.; Moura, B.B.; Seabra, A.B.; Salgado, I.; Machado, E.C.; Hancock, J.T.; Ribeiro, R.V. S-nitrosoglutathione spraying improves stomatal conductance, Rubisco activity and antioxidant defense in both leaves and roots of sugarcane plants under water deficit. Physiol. Plant 2017, 160, 383–395. [Google Scholar] [CrossRef]
- Badem, A.; Söylemez, S. Effects of nitric oxide and silicon application on growth and productivity of pepper under salinity stress. J. King Saud. Univ. Sci. 2022, 34, 102189. [Google Scholar] [CrossRef]
- Cunha, A.R.; Martins, D. Classificação climática para os municípios de Botucatu e São Manuel, SP. Irriga 2009, 14, 1–11. [Google Scholar] [CrossRef]
- Furlani, P.R.; Silveira, L.C.P.; Bolonhezi, D.; Faquim, V. Cultivo Hidropônico de Plantas; Instituto Agronômico: Campinas, Brazil, 1999; 52p. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b; as well as total carotenoids; using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Ferreira, D.F. Sisvar: A computer statistical analysis system. Cienc. Agrotecnologia 2011, 35, 1039–1042. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosynthetic Pigments | |||||
|---|---|---|---|---|---|
| Source of Variation | DF | Mean Squares | |||
| Cla | Clb | Cla + Clb | Carotenoids | ||
| Saline levels (SL) | 1 | 0.00003 ns | 0.00032 ns | 0.00015 ns | 0.00390 ** |
| SNP conc. (SC) 1 | 4 | 0.07817 ns | 0.15612 ns | 0.43721 ns | 0.00096 ns |
| SL × SC | 4 | 0.04235 ns | 0.07828 ns | 0.23439 ns | 0.00316 ** |
| CV (%) | 27.18 | 25.89 | 26.18 | 11.98 | |
| Saline levels | Test of means | ||||
| 0.2 dS m−1 | 0.79 a | 1.35 a | 2.15 a | 0.17 b | |
| 3.5 dS m−1 | 0.79 a | 1.36 a | 2.15 a | 0.19 a | |
| Gas Exchange | |||||||
|---|---|---|---|---|---|---|---|
| Source of Variation | DF | Mean Squares | |||||
| A | gs | Ci | E | WUE | CE | ||
| Saline levels (SL) | 1 | 46.29 ** | 0.0462 ** | 6521.46 ** | 14.81 ** | 0.76 ** | 0.00014 ** |
| SNP conc. (SC) 1 | 4 | 36.98 ** | 0.0130 ** | 3299.54 ** | 6.90 ** | 0.95 ** | 0.00053 ** |
| SL × SC | 4 | 1.17 ns | 0.0043 ** | 5328.74 ** | 0.56 * | 0.55 * | 0.00026 ** |
| CV (%) | 6.71 | 9.22 | 3.93 | 9.97 | 5.72 | 7.70 | |
| Saline levels | Test of means | ||||||
| 0.2 dS m−1 | 13.14 a | 0.19 a | 257.36 a | 4.82 a | 2.85 b | 0.051 a | |
| 3.5 dS m−1 | 10.99 b | 0.12 b | 231.82 b | 3.61 b | 3.13 a | 0.048 b | |
| Morphophysiological Characteristics | |||||
|---|---|---|---|---|---|
| Source of Variation | DF | Mean Squares | |||
| RWC | LA | SD | TDM | ||
| Saline levels (SL) | 1 | 244.89 ** | 11,425,114.60 ** | 146.6 ** | 69.16 ** |
| SNP conc. (SC) 1 | 4 | 10.88 ns | 395,811.06 ns | 2.91 ns | 4.86 ns |
| SL × SC | 4 | 10.29 ns | 57,283.00 ns | 10.08 ns | 6.00 ns |
| CV (%) | 4.86 | 15.68 | 9.58 | 14.08 | |
| Saline levels | Test of means | ||||
| 0.2 dS m−1 | 78.76 a | 3612.78 a | 22.24 a | 21.90 a | |
| 3.5 dS m−1 | 73.81 b | 2543.90 b | 18.41 b | 19.27 b | |
| Productive Parameters | |||
|---|---|---|---|
| Source of Variation | DF | Mean Squares | |
| NL | TFM | ||
| Saline levels (SL) | 1 | 55.22 ** | 359,883.80 ** |
| SNP conc. (SC) 1 | 4 | 15.41 * | 10,418.80 ** |
| SL × SC | 4 | 8.66 * | 4224.70 ** |
| CV (%) | 7.76 | 7.29 | |
| Saline levels | Test of means | ||
| 0.2 dS m−1 | 23.8 a | 417.74 a | |
| 3.5 dS m−1 | 21.5 b | 357.76 b | |
| pH | EC | N | P | K | Ca | Mg | S | Na |
|---|---|---|---|---|---|---|---|---|
| dS m−1 | mg L−1 | |||||||
| 6.18 | 0.20 | 22.00 | 1.00 | 10.00 | 14.00 | 4.00 | 4.00 | 0.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, I.C.d.S.; Silva, D.M.R.; Bispo, G.L.; Oliveira, F.d.A.d.; Ono, E.O.; Rodrigues, J.D. Nitric Oxide Modulates Salt Stress Tolerance in Lettuce. Stresses 2023, 3, 701-716. https://doi.org/10.3390/stresses3040048
Marques ICdS, Silva DMR, Bispo GL, Oliveira FdAd, Ono EO, Rodrigues JD. Nitric Oxide Modulates Salt Stress Tolerance in Lettuce. Stresses. 2023; 3(4):701-716. https://doi.org/10.3390/stresses3040048
Chicago/Turabian StyleMarques, Isabelly Cristina da Silva, Dayane Mércia Ribeiro Silva, Geane Lourenço Bispo, Francisco de Assis de Oliveira, Elizabeth Orika Ono, and João Domingos Rodrigues. 2023. "Nitric Oxide Modulates Salt Stress Tolerance in Lettuce" Stresses 3, no. 4: 701-716. https://doi.org/10.3390/stresses3040048
APA StyleMarques, I. C. d. S., Silva, D. M. R., Bispo, G. L., Oliveira, F. d. A. d., Ono, E. O., & Rodrigues, J. D. (2023). Nitric Oxide Modulates Salt Stress Tolerance in Lettuce. Stresses, 3(4), 701-716. https://doi.org/10.3390/stresses3040048