Abstract
Life does not start at birth but at conception. Embryonic development is a particularly difficult period in which genetic and environmental factors can interact to contribute to risk. In utero and early neonatal exposure to maternal stress are linked with psychiatric disorders, and the underlying mechanisms are currently being elucidated. This study examined novel relationships between maternal noise exposure causing oxidative-stress-induced neurobehavioral changes in cognition and autism spectrum disorder (ASD) in offspring. Pregnant Wistar albino rats were exposed to noise (100 dBA/4 h). There were three groups of pregnant rats exposed to noise during gestation, as well as a control group: early gestational stress (EGS), which occurs between the 1st and 10th days of pregnancy; late gestational stress (LGS), which occurs between the 11th day and the delivery day; and full-term gestational stress (FGS), which occurs during the entire pregnancy period. Maternal stress effects on the offspring were analyzed. This study observed that noise exposure becomes a psychosocial stressor in the prenatal period of motherhood. In the EGS and LGS groups, female rats showed continuous midterm abortion and stillbirth during noise exposure. The noise-exposed group exhibited significant changes in cognition, obsessive–compulsive behavior, fear, and anxiety. Corticosterone and oxidative stress markers increased, and the antioxidant level was significantly decreased in the noise-exposed group. Therefore, maternal noise exposure causes recurrent abortions and stillbirths, increases oxidative stress, and impairs the offspring’s neurodevelopment.
1. Introduction
A fetus’s early experiences significantly impact its health later in life. An individual’s pregnancy and motherhood are critical periods because they involve many neurobehavioral changes, including neuroendocrine, molecular, and physiological changes. These changes affect the mother’s mood, cognition, and stress regulation during gestation, parturition, and postpartum [1]. In line with scientific evidence, maternal stress, anxiety, and depression during pregnancy are associated with adverse mental health outcomes in offspring throughout their lives [2,3]. Recent research suggests that environmental pollutants are critical in fetal health [4,5,6,7].
Noise is a common environmental pollutant that is more than just a nuisance. Noise is defined as an unwanted sound; if the noise level exceeds 90decibel (dBA), it becomes a source of stress and impacts physical and psychological health [8]. The World Health Organization (WHO) has reported that at least 1 million healthy life years are lost annually due to environmental noise in high-income Western European nations [9].
Continuous noise exposure impairs hearing. Besides these auditory consequences, noise also triggers non-classical auditory-responsive brain areas, e.g., the lateral amygdala and striatum. It activates the hypothalamic–pituitary–adrenal (HPA) axis via the amygdala and other circuits, leading to the release of stress-related hormones, corticosterone, and adrenocorticotropic hormone (ACTH) [1]. During pregnancy, noise-exposure-induced production of corticosterone enters the placenta. Indeed, placental 11β-HSD2 inactivates the majority of maternal glucocorticoids passed to the fetus, and 11-β-hydroxysteroid dehydrogenase-type 1 (11β-HSD1) generates cortisol from inactive cortisone, which increases the stress hormones in fetal circulation [10]. From this view, this study aimed to evaluate the association of maternal noise stress with aberrant corticosterone-induced stillbirth, preterm birth, redox balance, and neurobehavioral alterations in offspring.
2. Results
2.1. Maternal Noise Exposure Altered the Gestational Length in Offspring
Maternal noise exposure altered the gestational period of the offspring (Figure 1). Recurrent abortions were shown in EGS and FGS groups. Stillbirths were observed in the progeny of LGS (12.6%) and FGS groups (27.6%). No stillbirths and abortions occurred in the control group. Figure 1A depicts the stillbirths of the pups, showing that the FGS group showed significantly higher stillbirths than LGS. Neonatal mortality was observed in EGS, LGS, and FGS groups. The FGS group had a higher mortality rate (28.6%) than the EGS (2%) and LGS (14.3%). No neonatal mortality was observed in the control group. Figure 1B illustrates the neonatal mortality rate. No congenital disabilities occurred in the offspring of the control or EGS groups. The percentage of congenital disabilities in the LGS and FGS groups was 23.3% and 10.3%, respectively. The data confirm that the FGS group has significantly higher congenital disabilities than the LGS group. Figure 1C depicts the congenital disabilities observed in the offspring of LGS and FGS groups.
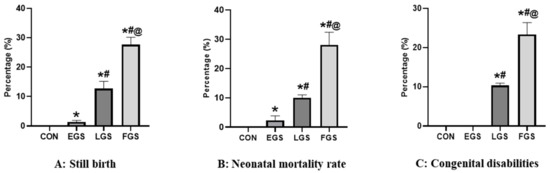
Figure 1.
Maternal noise exposure altered the gestational length in offspring. Maternal noise exposure significantly increased the stillbirth (A), neonatal mortality rate (B), and congenital disabilities (C) in LGS and FGS groups. The data represent the mean ± SD (p < 0.05—significance) (*—versus control, #—versus EGS, @—versus LGS).
2.2. Maternal Noise Exposure Decreased the Body Weight and Discrete Regions of the Brain in Offspring
The body weight of the newborn pups on the 1st and 45th postnatal days significantly differed between the experimental groups (Figure 2). The body weight of the newborn pups of the FGS group (4.16 ± 0.25) was substantially lower than either the control (6.6 ± 1.15) or EGS groups (5.66 ± 0.50) and the LGS group (4.7 ± 0.25) (Figure 2A). Compared to LGS, FGS had significantly lower body weight, and EGS had significantly higher body weight. The body weight of the offspring on the 45th day of postnatal life in the FGS group (42 ± 2 g) was significantly lower than in the control (76.09 g ± 2.5 g) and EGS groups (59.6 ± 0.56 g) and LGS group (51 ± 2.64 g) group (Figure 2B). Compared to EGS, LGS had significantly lower body weight, and the control group had significantly higher body weight. In discrete regions of brain weight, prefrontal cortex, and hippocampus, the weight was significantly reduced in the FGS group compared to the control, EGS, and LGS animals. The EGS group showed a significantly decreased brain weight, prefrontal cortex, and hippocampus weight compared to the control and LGS groups (Table 1). All experimental groups had no significant change in the cerebellum and hypothalamus weight.
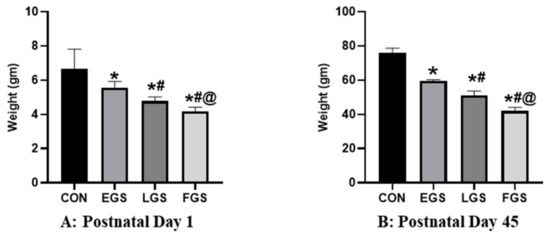
Figure 2.
Maternal noise exposure decreased the body weight and discrete regions of the brain in offspring. On postnatal day 1 (A) and 45th day (B), the weight of the EGS-, LGS-, and FGS-group animals was significantly reduced compared to the control group. The data represent the mean ± SD. The significance level was p < 005, * versus control, # versus EGS, and @ versus LGS.

Table 1.
Maternal noise exposure decreased weight of the discrete regions of the offspring’s brain.
2.3. Maternal Noise Exposure Altered the Offspring’s Cognitive and Emotional Functioning
Maternal noise exposure alterd the offspring’s neurobehavior, including impaired spatial learning and memory, increased fear, anxiety, and repetitive behavior (Figure 3). In this study, the results of the T-maze showed increased latency and number of days taken to make the correct choice in EGS- and FGS-group animals compared to the control and LGS animals (Figure 3A). The EGS group’s (11.6 ± 1.15) latency significantly increased compared to the control (7.3 ± 0.57) and LGS (14 ± 1), and the FGS group’s (18.4 ± 0.48) latency was significantly higher than the EGS group. Similarly, the number of days the EGS (11.3 ± 1.5) group took increased compared to the control (8 ± 1) and was lesser than the LGS group (13 ± 1) and FGS group (16.6 ± 1.52). In the open-field test, the EGS group (16.3 ± 15.2) showed decreased time spent in the central square when compared to the control (226.6 ± 25.1), and increased time compared to the LGS (126.6 ± 7.63) and FGS group (94.3 ± 9.29). The LGS- (173.6 ± 7.6) and FGS-exposed animals (205 ± 9.2) showed an increased time spent in peripheral squares compared to the control (73.3 ± 25.12) and EGS animals (136.6 ± 15.2), which increased the fear and anxiety of the animals (Figure 3B). In the marble burying test, the number of marbles buried was higher in the LGS (11 ± 1) and FGS (15 ± 1) groups compared to the control (5 ± 1) and EGS (9 ± 1) groups. Compared to the control, the number of marbles buried by the EGS group was significantly higher, while the number buried by the LGS and FGS groups was lower (Figure 3C). In the NST, the amount of cotton shredding was significantly increased in the LGS (0.59 ± 0.02) and FGS groups (0.83 ± 0.05) compared to the control (0.06 ± 0.05) and EGS (0.20 ± 0.007) groups (D).
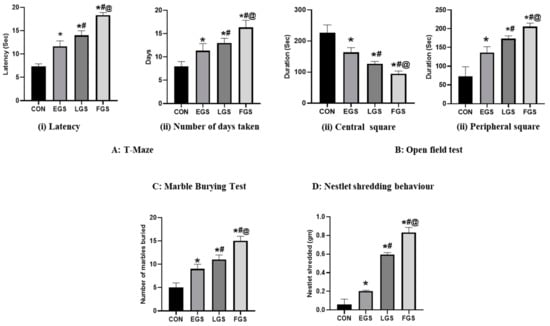
Figure 3.
Maternal noise exposure altered the offspring’s cognitive and emotional functioning. (A) T-maze results confirmed that maternal noise impairs spatial memory and learning in EGS, LGS, and FGS groups. (B) The open-field test showed the reduced time spent in the central square indicated the fear and anxiety-like behavior of the EGS, LGS, and FGS animals compared to the control. MBT (C) and NSB (D) behaviors showed autistic and repetitive obsessive–compulsive behavior in stress groups compared to the control animals. The data represent the mean ± SD. The significance level was p < 0.05, * versus control, # versus EGS, and @ versus LGS.
2.4. Maternal Noise Exposure Impairs Corticosterone and the Redox Balance in the Offspring’s Brain
The CORT level (Figure 4) in plasma was significantly increased in the LGS (56.92 ± 0.7) and FGS (60.41 ± 1.3) groups compared to the control (42.94 ± 0.5) and EGS (51.34 ± 1.5) group animals. The CORT level in the mPFC was significantly increased in EGS (179 ± 1.4), LGS (188 ± 2.8), and FGS (212 ± 2.7) compared to control (157 ± 3.5). Similarly, the CORT level in the CB was significantly higher in the EGS (83.6 ± 4.3), LGS (91.2 ± 0.9), and FGS (108.2 ± 3.6) groups when compared to the control (76.8 ± 1.4). The CORT level in the HI was significantly increased in the LGS (122.7 ± 1.7) and FGS (157 ± 4.2) groups compared to the control- (94.2 ± 3.1) and EGS (107.8 ± 3.7)-group animals. Additionally, the CORT level in the HY was significantly increased in the EGS (58 ± 2.8), LGS (66.5 ± 2.1), and FGS (77 ± 1.4) compared to the control (48.5 ± 3.5). The LPO level (Table 2A) was significantly increased in the FGS group compared to the control-, EGS-, and LGS-group animals. The mPFC and hippocampus LPO levels were increased compared to the control-, EGS-, and LGS-group animals. There was no significant change in LPO level in the hypothalamus and cerebellum. The SOD level (Table 2B) in the hypothalamus was shown to be increased significantly in the control group when compared to the EGS, LGS, and FGS groups. In comparison, the prefrontal cortex, cerebellum, and hippocampus were insignificant. The catalase level (Table 2C) was significantly altered in the mPFC and hippocampus and no significant change in the hypothalamus or the cerebellum was observed in all experimental groups.
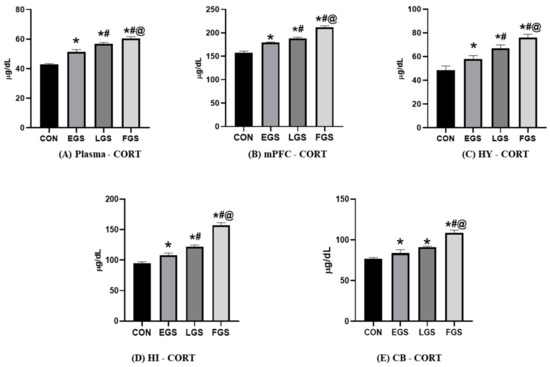
Figure 4.
Maternal noise exposure altered corticosterone levels in the offspring’s brain: (A) shows the plasma corticosterone of all experimental groups; (B–E) shows the corticosterone level in discrete brain regions of EFS, LGS, FGS, and control animals. The data represent the mean ± SD. The significance level was p < 0.05, * versus control, # versus EGS, and @ versus LGS.
Table 2.
Maternal noise exposure decreased antioxidant levels and increased oxidative stress in the discrete regions of the offspring’s brain.
3. Discussion
The mother’s body provides the resources and humoral factors essential for the fetus’s intrauterine development [11]. Several factors have been associated with the stunting of children and poor neurodevelopment outcomes, including perinatal malnutrition, inflammation, environmental factors, and maternal illiteracy [12]. Noise exposure may adversely affect health, including birth outcomes. Studies have examined effects such as preterm birth, low birth weight, small size for gestational age, and congenital malformations. In addition to causing biological reactions to stress, noise exposure can increase the risk of adverse prenatal outcomes, including adverse pregnancy outcomes [13].
Our study results show that exposure to noise during early and full-term gestation causes recurrent abortion, and late gestational periods cause stillbirth, preterm birth, and congenital disabilities in newborn pups. Various studies confirmed that maternal stress causes recurrent abortion, stillbirth, preterm birth, and congenital disabilities in offspring [14,15,16,17,18]. A meta-analysis study has mentioned that high noise exposure during pregnancy increases the risk of early gestational age, gestational hypertension, and congenital disabilities in newborns [19]. Research has shown that activating the HPA axis increases blood levels of glucocorticoids [20]. It has also been demonstrated that high levels of glucocorticoids adversely affect the uterus and fetus and inhibit the secretion of pituitary luteinizing hormone, estrogen, and progesterone by the ovary. A lack of progesterone during pregnancy can also be attributed to stress hormone-induced inhibitory effects on female reproductive organs, resulting in the symptoms known as “hypothalamic” amenorrhea and fetal loss [21]. Studies have previously shown that noise exposure stimulates HPA axis activity and increased corticosterone production [22]. Thus, exposure to noise results in increased glucocorticoid levels, which impair uteroplacental function, resulting in recurrent abortions and stillbirths among mothers exposed to noise.
Several mediators of the maternal stress response impact the embryonic environment’s redox balance, such as inflammation and cortisol production. This study also demonstrated that maternal noise exposure increased the corticosterone and oxidative stress markers and decreased antioxidant level in plasma and discrete regions of the offspring’s brain. As a result of gestational noise stress, Zahra Jafari et al. found that offspring of dams exposed to gestational noise stress exhibited increased corticosterone levels, altered cognitive function, and amyloid beta development as they aged [23]. Hanafi et al. reported that prenatal noise exposure (90 dBA) to embryonic chicks and newly hatched chicks caused elevated serum corticosterone and ceruloplasmin (CPN) levels [24]. Elevated corticosterone suppresses LTP, synaptic plasticity, and GABAergic modulation, which impairs spatial learning and memory and increases anxiety-like behavior and neurogenesis in the CA3-CA1 pathway of the hippocampus [24,25,26,27]. The young rats born from dams exposed to noise stress during the third trimester of pregnancy showed increased corticosterone, impaired spatial memory, and increased anxiety-like behaviors [28]. Another study showed that prenatal noise stress significantly increased terminal plasma ACTH concentration, impaired balance control, locomotion, horizontal and vertical exploration, and increased anxiety levels in offspring [29]. Therefore, increased CORT levels in the brain and circulation modulates the release of the neurotransmitter, further suppressing the synaptic functions in the hippocampus and impairing cognition and emotional behaviors in noise-exposed rats. The embryonic brain has an exceptionally high risk of oxidative stress, as antioxidant enzymes are expressed at relatively low levels. High concentrations of catalytic metals, including Fe2+ and Cu+, promote hydroxyl radical production from hydrogen peroxide in the cell environment. There is also evidence that maternal stress during pregnancy increases lipid peroxidation (LPO) in the offspring’s postnatal brain, suggesting that redox balance in the brain may be altered for a long time after the baby is born [30]. Liegelin Kavitha Bernhardt et al. reported that prenatal stress increased oxidative stress, including increased malondialdehyde (MDA) levels, reduced glutathione (GSH), superoxide dismutase (SOD), and total antioxidant activity (TAO), while glutathione reductase (GSS-Rd) levels remained low in offspring’s brain [31].
Furthermore, this study confirmed that maternal noise exposure altered spatial learning and memory, increased fear, and anxiety-like behavior, and increased repetitive behaviors in offspring. Scott et al. demonstrated that placental oxidative stress is the critical mediator in transmitting maternal social stress impacts on the offspring’s brain and behavior, precisely due to the reduction in hippocampal dendrite length, GABA receptor subunits, and the number of parvalbumin-positive neurons and basolateral amygdala in male offspring [32]. A calcium-binding protein, PARValbumin (PV), is expressed by inhibitory fast-spiking interneurons in the cerebral cortex. It has also been found that changes in the morphology or function of PV+ interneurons are associated with schizophrenia, epilepsy, bipolar disorder, autism, and many other psychiatric disorders [33]. Our previous studies have demonstrated that excessive noise exposure altered hippocampal morphology, which impaired cognition and caused autistic-like repetitive behavior in rats [8]. From the abovementioned, excessive CORT-induced oxidative stress and decreased antioxidant levels in discrete brain regions alter neurons’ structural and functional processing causing neuropsychiatric behaviors in noise-exposed male progeny. A recent study has confirmed that prenatal and postnatal noise exposure increased plasma corticosterone levels, decreased the number of neurons, and increased the number of apoptotic cells; a decrease in the volume of mPFC and non-neuron cells was observed in noise-exposed Wistar albino offspring [34]. Sajjadi et al. mentioned that pregnant rats being exposed to urban traffic noise exposure during the last trimester of pregnancy led to impaired spatial learning and memory in their offspring, elevated corticosterone concentration, and reduced GRs and MRs gene expression in the hippocampus [35]. It was also found that mild prenatal stress (noise, restraint, strobe lights) changes the structure of the bed nucleus of stria terminals (BNST) and the hippocampus, decreases norepinephrine and dopamine levels in the BNST, and lowers serotonin levels in the hippocampus [36]. The research above shows that excessive CORT-induced oxidative stress and decreased antioxidant levels in discrete brain regions alter brain function. A new finding in our study was that noise increased CORT, which further affected redox homeostasis, the main factor limiting brain structural and functional roles. Therefore, these findings show that noise exposure during pregnancy can lead to long-lasting structural and neurochemical effects in offspring, affecting adult behavior.
4. Materials and Methods
4.1. Animal Maintenance
Adult Wistar albino rats (Rattus norvegicus) of both sexes, male (180–220 gm) and female (160–180 gm), were used for this study. The present experimental study was approved by the institutional animal ethics committee (IAEC No.: 03/03/2019). The Wistar albino rats were obtained and maintained in clean polypropylene cages at the Central Animal House Facility, University of Madras (Taramani campus), under a 16 h light/8 h dark cycle at 230 C with 60% humidity and given ad libitum access to food and water when the experimental procedures were conducted. Each cage housed five animals.
4.2. Experimental Design
Three female rats were allowed to mate with one fertile, sexually active male rat for one night. The following day, the vaginal plug was collected from the females. A vaginal smear was prepared and examined under a microscope for the presence of sperm. The presence of sperm confirmed pregnancy; that day was recorded as “day zero” of pregnancy. The pregnant rats were then housed individually in separate cages. Animals were randomly divided into four groups (n = 6) and used for the experimental studies. The pregnant rats were exposed to noise stress using a noise-induction chamber for four hours daily. Male progeny (n = 6) were used for this experimental study. There were three groups of pregnant rats exposed to noise during gestation, as well as a control group: early gestational stress (EGS), which occurs between the 1st and 10th days of pregnancy; late gestational stress (LGS), which occurs between the 11th day and the delivery day, and full-term gestational stress (FGS), which occurs during the entire pregnancy period. After the 45th postnatal day, infant pups were tested using the T-maze test, marble-burying test (MBT), open-field test, and nestlet-shredding-behavior test (NST), and sacrificed (Figure 5).
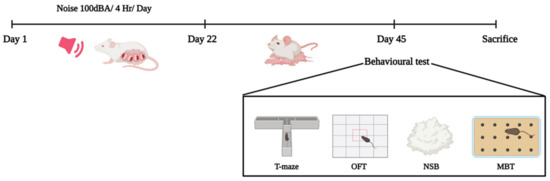
Figure 5.
Experimental design for the maternal-noise-induced neurobehavioral changes in offspring.
4.3. Noise Exposure
The noise was produced by a loudspeaker (15 W) installed 30 cm above the cage and driven by a white-noise generator emitting all the frequencies in the 0–20 kHz range. A precision sound level meter was used to uniformly set the sound intensity to 100 dB in the cage. During the experiment, the noise level peaked at 100 dB immediately after the generator was switched on and lasted 4 h/day [37,38].
4.4. Gestational Length
Gestational length is the number of days taken for parturition from the day of pregnancy. Each group’s gestational length was noted and compared.
4.4.1. Stillbirth
Newborn pups were carefully examined after parturition and dead pups were considered stillbirths. The percentage of stillbirths was calculated as follows:
4.4.2. Congenital Disabilities
Congenital disabilities are any structural or physiological abnormality present at birth that develops before or during birth. The newborn pups were carefully inspected for congenital disabilities, and the percentage of congenital disabilities was calculated as follows:
4.4.3. Neonatal Mortality Rate
Death of newborn pups (stillborn animals excluded) within three days following parturition is called neonatal mortality. The percentage of neonatal mortality was calculated as follows:
4.5. Measurement of Bodyweight
During delivery and after 45 days of postnatal life, the body weight of newborn pups in all experimental groups was measured. The body weights of infant pups were compared among all experimental groups on the day of delivery. Additionally, newborn pups’ body weights were compared among all experimental groups on the 45th postnatal day.
4.6. Behavioral Analysis
4.6.1. T-Maze
T-mazes are used to study how rodents function with memory and spatial learning by applying various stimuli. The T-maze consists of a start box (16.5 cm × 16.5 cm), stem (arm 1) length is 71.1 cm, goal area (arms 2 and 3) lengths are 45.7 cm and arm width is 10.2 cm. A sliding door separates the stem and start box, and T-maze is kept in a dimly lit, sound-attenuated room. After the 1st day of orientation, the rats were trained for the rewarded alternation task. The learning performance test followed the same procedure as the orientation session, except that one arm was baited. The rat must alternate the arms for the reward. The placement of the reward in the subsequent trial will be opposite to the rats’ previous arm entry. Each rat underwent 10 trials per day. The intertrial interval was 30 s, during which the maze was cleaned with 70% alcohol to remove odor cues. The number of days each rat took to achieve 80% correct choices (8 correct entries to the baited arm out of 10 trials) and the time taken to reach the goal area were noted. Then, a memory-retention test was carried out two days after the last training session. Rats underwent 10 trials continuously, with an intertrial interval of 30 s. The number of errors, i.e., entry into the non-rewarded arm recorded in ten trials, was noted. The movement of each rat was recorded using a digital camera. Each record was converted to image sequences to track rat movement using the Image J (V1.53c) plugin, Animal Tracker 14. The latency, correct choices, and the number of days taken for the correct choice, retention memory, latency and the correct choice of each rat were measured [39].
4.6.2. Open-Field Behavior
Open-field behavior is a simple test to evaluate the locomotion and emotional status of the animals. The apparatus used was a large rectangular box (100 cm × l00 cm) with 40-cm-high plywood walls. The floor consisted of a clean, dark plastic material with a grid painted in white, dividing the field into 25 (5 × 5) equal squares. A 60 W bulb placed 100 cm above the center of the area provided illumination. A weak cider vinegar solution (10%) was used to clean the apparatus before the introduction of each animal. The rat was placed in one corner of the apparatus, and its behavior was observed for 5 min, estimating the time spent in the central square and time spent in the peripheral square of the offspring [40].
4.6.3. Marble-Burying Behavior Test
The marble-burying behavior test assessed the obsessive–compulsive behavior of infant rats. The polycarbonate cage (26 cm × 48 cm × 20 cm) was filled with bedding material to the depth of 5 cm, and in it were kept 20 standard glass marbles (15 mm DM, 5.2 g in weight), in 5 rows of 4 marbles. Then, rats were allowed in that cage for 30 min. Water and food were withheld during the experiment. Scoring was given when the rat buried two-thirds of a marbles surface into the bedding material [41].
4.6.4. Nestlet-Shredding Behavior Test
The polycarbonate cage (26 cm × 48 cm × 20 cm) was filled with bedding material, and 1 g of nestlet (cotton bed) was placed on the bed material. Then, the rat was allowed in that cage for 30 min. Water and food were withheld during the experiment. Scoring was given by weighing the remaining unshredded nestlet and dividing this weight by the starting weight to calculate the percentage of the nestlet shredded [42].
4.7. Biochemical Assay
After the completion of behavioral analysis, rats were killed via cervical dislocation. A sample collection was conducted between 09:00 and 10:00 AM to avoid variations caused by circadian rhythms. Plasma and serum were separated via centrifugation at 3000 rpm at 4 °C for 15 min. The brain was immediately removed and washed with ice-cold saline. Various brain parts—like the cerebral cortex, cerebellum, hippocampus, and hypothalamus—were dissected in ice-cold (4 °C) conditions. A motor-driven Teflon-glass tissue homogenizer was used to prepare 10% homogenate using ice-cold Tris–HCl (100 mM, pH 7.4) buffer. The homogenate was centrifuged at 2000 rev min−1, 4 °C for 15 min, and the supernatant was used for the biochemical analysis. For all experimental groups, previously described methods were used to estimate the corticosterone (CORT) [43], lipid peroxidation (LPO), superoxide dismutase (SOD), and catalase levels [37].
4.8. Statistical Analysis
Data are presented as a bar diagram with mean ± standard deviation. The results were analyzed using Graph Pad Prism software (Version 8.0). For the multivariable analyses, we used ANOVA with Tukey’s multiple comparison test; p < 0.05 was considered statistically significant; “*” refers to “compared to control”, “#” to “compared to EGS group”, and “@” to “compared to LGS”.
5. Conclusions
This study has demonstrated that maternal noise exposure dysregulated the HPA axis, increased oxidative stress, and decreased antioxidant levels. This impaired cognition, increased fear, anxiety, repetitive behavior, accelerated oxidative stress, and caused recurrent abortions, stillbirths, and neurobehavioral changes in offspring. Future studies should investigate concurrent alternations in levels of oxidative and antioxidant markers, identifying the molecular mechanism behind maternal-noise-exposure-induced neurobehavioral changes in offspring.
Author Contributions
Conceptualization: A.A. and R.R.; data curation: A.A. and D.K.S.; investigation: A.A. and R.R.; methodology: A.A. and R.R.; project administration: R.R.; software: D.K.S.; supervision: R.R.; writing—original draft: A.A. and D.K.S.; writing—review and editing: D.K.S. and R.R. All authors have read and agreed to the published version of the manuscript.
Funding
The Research Fellowship and Associateships (SRF) grant funded this study through the Government of India, funded by the Indian Council of Medical Research (ICMR), New Delhi, India (No. RBMH/FW/2020/31).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
All authors acknowledged Biorender.com in creating the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Noise exposure accelerates the risk of cognitive impairment and Alzheimer’s disease: Adulthood, gestational, and prenatal mechanistic evidence from animal studies. Neurosci. Biobehav. Rev. 2020, 117, 110–128. [Google Scholar] [CrossRef] [PubMed]
- Lipschutz, R.; Kulesz, P.A.; Elgbeili, G.; Biekman, B.; Laplante, D.P.; Olson, D.M.; King, S.; Bick, J. Maternal mental health mediates the effect of prenatal stress on infant temperament: The Harvey Mom Study. Dev. Psychopathol. 2023, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bergh, B.R.V.D.; Heuvel, M.I.V.D.; Lahti, M.; Braeken, M.; de Rooij, S.R.; Entringer, S.; Hoyer, D.; Roseboom, T.; Räikkönen, K.; King, S.; et al. Prenatal developmental origins of behavior and mental health: The influence of maternal stress in pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 26–64. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Q.; He, S.; Wu, K.; Ren, M.; Dong, H.; Di, J.; Yu, Z.; Huang, C. Ambient air pollution and gestational diabetes mellitus: A review of evidence from biological mechanisms to population epidemiology. Sci. Total Environ. 2020, 719, 137349. [Google Scholar] [CrossRef] [PubMed]
- Ju, L.; Hua, L.; Xu, H.; Li, C.; Sun, S.; Zhang, Q.; Cao, J.; Ding, R. Maternal atmospheric particulate matter exposure and risk of adverse pregnancy outcomes: A meta-analysis of cohort studies. Environ. Pollut. 2023, 317, 120704. [Google Scholar] [CrossRef]
- Hung, T.H.; Chen, P.H.; Tung, T.H.; Hsu, J.; Hsu, T.Y.; Wan, G.H. Risks of preterm birth and low birth weight and maternal exposure to NO2/PM2. 5 acquired by dichotomous evaluation: A systematic review and meta-analysis. Environ. Sci. Pollut. Res. 2022, 30, 9331–9349. [Google Scholar] [CrossRef]
- Thomson, E.M. Air Pollution, Stress, and Allostatic Load: Linking Systemic and Central Nervous System Impacts. J. Alzheimer’s Dis. 2019, 69, 597–614. [Google Scholar] [CrossRef]
- Arjunan, A.; Rajan, R. Noise and brain. Physiol. Behav. 2020, 227, 113136. [Google Scholar] [CrossRef]
- Basner, M.; Babisch, W.; Davis, A.; Brink, M.; Clark, C.; Janssen, S.; Stansfeld, S. Auditory and non-auditory effects of noise on health. Lancet 2014, 383, 1325–1332. [Google Scholar] [CrossRef]
- I Stirrat, L.; Sengers, B.G.; E Norman, J.; Homer, N.Z.M.; Andrew, R.; Lewis, R.M.; Reynolds, R.M. Transfer and Metabolism of Cortisol by the Isolated Perfused Human Placenta. J. Clin. Endocrinol. Metab. 2018, 103, 640–648. [Google Scholar] [CrossRef]
- Manikandan, S.; Devi, R.S. Antioxidant property of α-asarone against noise-stress-induced changes in different regions of rat brain. Pharmacol. Res. 2005, 52, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.; Devi, R.S.; Ravindran, R.; Senthilvelan, M. Effect of noise stress on free radical scavenging enzymes in brain. Environ. Toxicol. Pharmacol. 2005, 20, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar]
- Deacon, R.M.J. Digging and marble burying in mice: Simple methods for in vivo identification of biological impacts. Nat. Protoc. 2006, 1, 122–124. [Google Scholar] [CrossRef]
- Thompson, S.L.; Welch, A.C.; Ho, E.V.; Bessa, J.M.; Portugal-Nunes, C.; Morais, M.; Young, J.W.; Knowles, J.A.; Dulawa, S.C. Btbd3 expression regulates compulsive-like and exploratory behaviors in mice. Transl. Psychiatry 2019, 9, 222. [Google Scholar] [CrossRef]
- Sundareswaran, L.; Srinivasan, S.; Wankhar, W.; Sheeladevi, R. Effect of Scoparia dulcis on noise stress induced adaptive immunity and cytokine response in immunized Wistar rats. J. Ayurveda Integr. Med. 2017, 8, 13–19. [Google Scholar] [CrossRef]
- Gedzun, V.R.; Khukhareva, D.D.; Sarycheva, N.Y.; Kotova, M.M.; Kabiolsky, I.A.; Dubynin, V.A. Perinatal Stressors as a Factor in Impairments to Nervous System Development and Functions: Review of In Vivo Models. Neurosci. Behav. Physiol. 2023, 53, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, T.; Patro, N.; Patro, I.K. Cumulative multiple early life hits- a potent threat leading to neurological disorders. Brain Res. Bull. 2019, 147, 58–68. [Google Scholar] [CrossRef]
- Sivakumaran, K.; Ritonja, J.A.; Waseem, H.; AlShenaibar, L.; Morgan, E.; Ahmadi, S.A.; Denning, A.; Michaud, D.; Morgan, R.L. Impact of Noise Exposure on Risk of Developing Stress-Related Obstetric Health Effects: A Systematic Review and Meta-Analysis. Noise Health 2022, 24, 137. [Google Scholar]
- Liu, Y.; Zhang, C.; Liu, Y.; Zhu, J.; Qu, H.; Zhou, S.; Chen, M.; Xu, D.; Chen, L.; Wang, H. Paternal Nicotine/Ethanol/Caffeine Mixed Exposure Induces Offspring Rat Dysplasia and Its Potential “GC-IGF1” Programming Mechanism. Int. J. Mol. Sci. 2022, 23, 15081. [Google Scholar] [CrossRef]
- Nyadanu, S.D.; Tessema, G.A.; Mullins, B.; Kumi-Boateng, B.; Ofosu, A.A.; Pereira, G. Prenatal exposure to long-term heat stress and stillbirth in Ghana: A within-space time-series analysis. Environ. Res. 2023, 222, 115385. [Google Scholar] [CrossRef]
- Garcia-Flores, V.; Romero, R.; Furcron, A.-E.; Levenson, D.; Galaz, J.; Zou, C.; Hassan, S.S.; Hsu, C.-D.; Olson, D.; Metz, G.A.S.; et al. Prenatal Maternal Stress Causes Preterm Birth and Affects Neonatal Adaptive Immunity in Mice. Front. Immunol. 2020, 11, 254. [Google Scholar] [CrossRef]
- Yu, F.; Xu, X.; Ren, Z.; Yang, J.; Kong, F. Maternal high-decibel acoustic exposure elevates prenatal stress, impairing postnatal hearing thresholds associated with decreasing ribbon synapses in young rats. Reprod. Toxicol. 2019, 89, 21–27. [Google Scholar] [CrossRef]
- Oinam, B.; Anand, V.; Kajal, R.K. A geospatial based hotspot and regression analysis of abortion and stillbirth prevalence in Manipur, India. Women Health 2021, 61, 599–608. [Google Scholar] [CrossRef]
- Dzhambov, A.M.; Dimitrova, D.D.; Dimitrakova, E.D. Noise Exposure During Pregnancy, Birth Outcomes And Fetal Development: Meta-Analyses Using Quality Effects Model. Folia Med. 2014, 56, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 21, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Nepomnaschy, P.A.; Sheiner, E.; Mastorakos, G.; Arck, P.C. Stress, immune function and reproduction. Ann. NY Acad. Sci. 2007, 1113, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Fe’Li, S.N.; Esmaielpour, M.R.M.; Hokmabadi, R.; Rezaei-Hachesu, V. Effect of occupational noise exposure on cortisol hormone level: A systematic review. Noise Vib. Worldw. 2022, 53, 579–588. [Google Scholar] [CrossRef]
- Jafari, Z.; Okuma, M.; Karem, H.; Mehla, J.; Kolb, B.E.; Mohajerani, M.H. Prenatal noise stress aggravates cognitive decline and the onset and progression of beta amyloid pathology in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2019, 77, 66–86. [Google Scholar] [CrossRef]
- Hanafi, S.; Zulkifli, I.; Ramiah, S.; Chung, E.; Kamil, R.; Awad, E. Prenatal auditory stimulation induces physiological stress responses in developing embryos and newly hatched chicks. Poult. Sci. 2023, 102, 102390. [Google Scholar] [CrossRef]
- Jafari, Z.; Mehla, J.; Kolb, B.E.; Mohajerani, M.H. Prenatal noise stress impairs HPA axis and cognitive performance in mice. Sci. Rep. 2017, 7, 10560. [Google Scholar] [CrossRef] [PubMed]
- Barzegar, M.; Sajjadi, F.S.; Talaei, S.A.; Hamidi, G.; Salami, M. Prenatal exposure to noise stress: Anxiety, impaired spatial memory, and deteriorated hippocampal plasticity in postnatal life. Hippocampus 2015, 25, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-W.; Shin, M.-S.; Park, J.-K.; Shin, M.-A.; Lee, H.-H.; Lee, S.-J. Treadmill exercise alleviates prenatal noise stress-induced impairment of spatial learning ability through enhancing hippocampal neurogenesis in rat pups. J. Exerc. Rehabil. 2013, 9, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Hadizadeh, M.; Hamidi, G.; Salamim, M. Probiotic supplementation improves the cognitive function and the anxiety-like behaviors in the stressed rats. Iran. J. Basic Med. Sci. 2019, 22, 506–514. [Google Scholar] [CrossRef]
- Badache, S.; Bouslama, S.; Brahmia, O.; Baïri, A.M.; Tahraoui, A.K.; Ladjama, A. Prenatal noise and restraint stress interact to alter exploratory behavior and balance in juvenile rats, and mixed stress reverses these effects. Stress 2017, 20, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Dowell, J.; Elser, B.A.; Schroeder, R.E.; Stevens, H.E. Cellular stress mechanisms of prenatal maternal stress: Heat shock factors and oxidative stress. Neurosci. Lett. 2019, 709, 134368. [Google Scholar] [CrossRef]
- Bernhardt, L.K.; Madhyastha, S.; Bairy, L.; Kishore, A. Status of the brain antioxidant system at different growing periods after prenatal stress and N -acetyl cysteine administration. Folia Neuropathol. 2017, 1, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Scott, H.; Phillips, T.J.; Sze, Y.; Alfieri, A.; Rogers, M.F.; Volpato, V.; Case, C.P.; Brunton, P.J. Maternal antioxidant treatment prevents the adverse effects of prenatal stress on the offspring’s brain and behavior. Neurobiol. Stress 2020, 13, 100281. [Google Scholar] [CrossRef]
- Juarez, P.; Cerdeño, V.M. Parvalbumin and parvalbumin chandelier interneurons in autism and other psychiatric disorders. Front. Psychiatry 2022, 13, 913550. [Google Scholar] [CrossRef]
- Teimouri, M.; Heidari, M.H.; Amini, A.; Sadeghi, Y.; Abdollahifar, M.-A.; Aliaghaei, A.; Khavanin, A.; Nadri, F.; Danyali, S.; Sanchooli, T. Neuroanatomical changes of the medial prefrontal cortex of male pups of Wistar rat after prenatal and postnatal noise stress. Acta Histochem. 2020, 122, 151589. [Google Scholar] [CrossRef] [PubMed]
- Sajjadi, F.; Aghighi, F.; Vahidinia, Z.; Azami-Tameh, A.; Salami, M.; Talaei, S. Prenatal urban traffic noise exposure impairs spatial learning and memory and reduces glucocorticoid receptor expression in the hippocampus of male rat offspring. Physiol. Int. 2020, 107, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Soares-Cunha, C.; Coimbra, B.; Borges, S.; Domingues, A.V.; Silva, D.; Sousa, N.; Rodrigues, A.J. Mild Prenatal Stress Causes Emotional and Brain Structural Modifications in Rats of Both Sexes. Front. Behav. Neurosci. 2018, 12, 129. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).