

Antioxidant Enzyme and Cytochrome P450 Activities Are Involved in Horseweed (Conyza Sumatrensis) Resistance to Glyphosate

 ,
,  and
and
Abstract
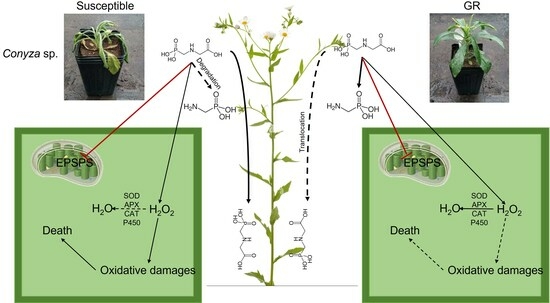
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
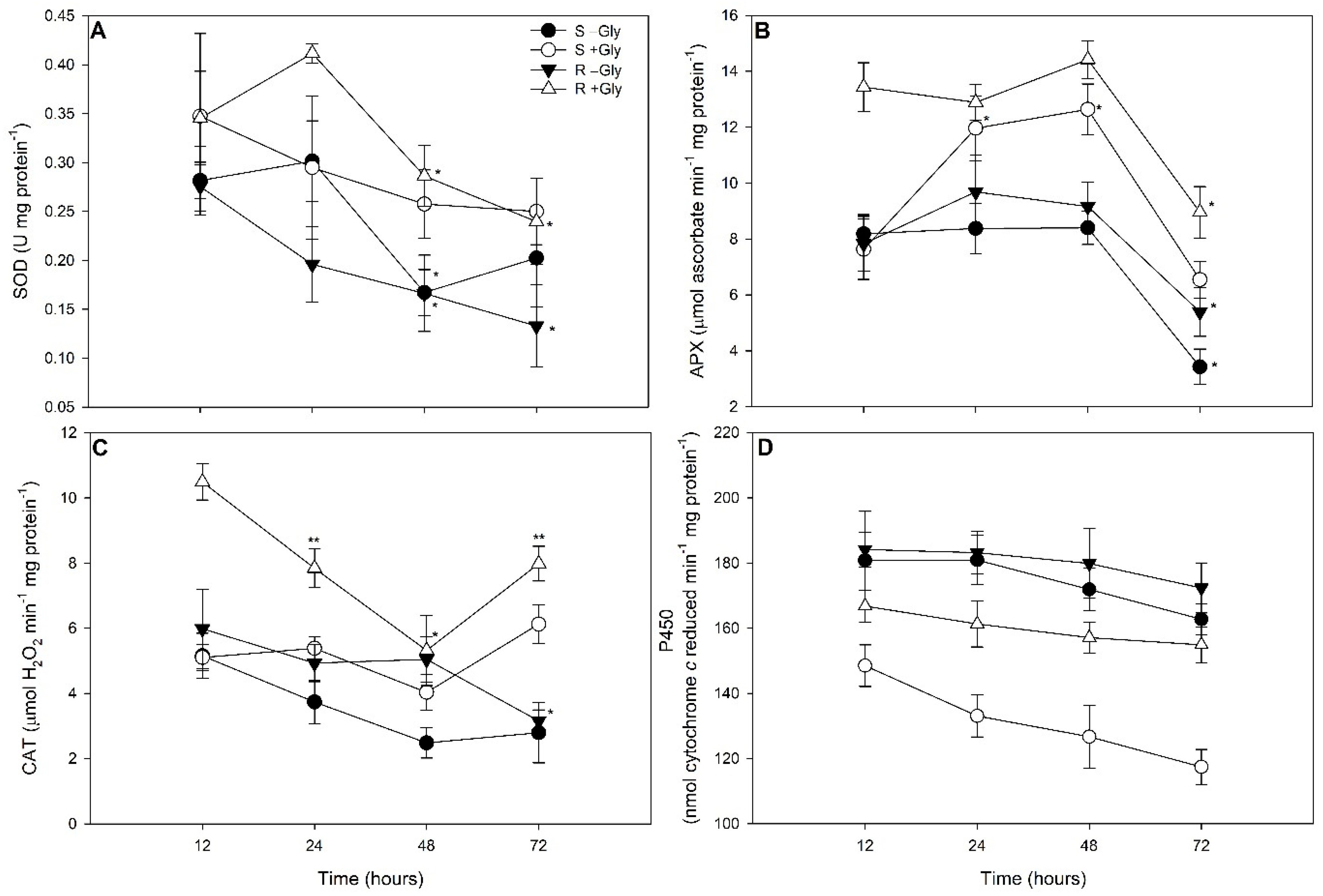
2.1. Oxidative Stress Markers
2.2. NADPH-Cytochrome P450 Reductase Evaluations
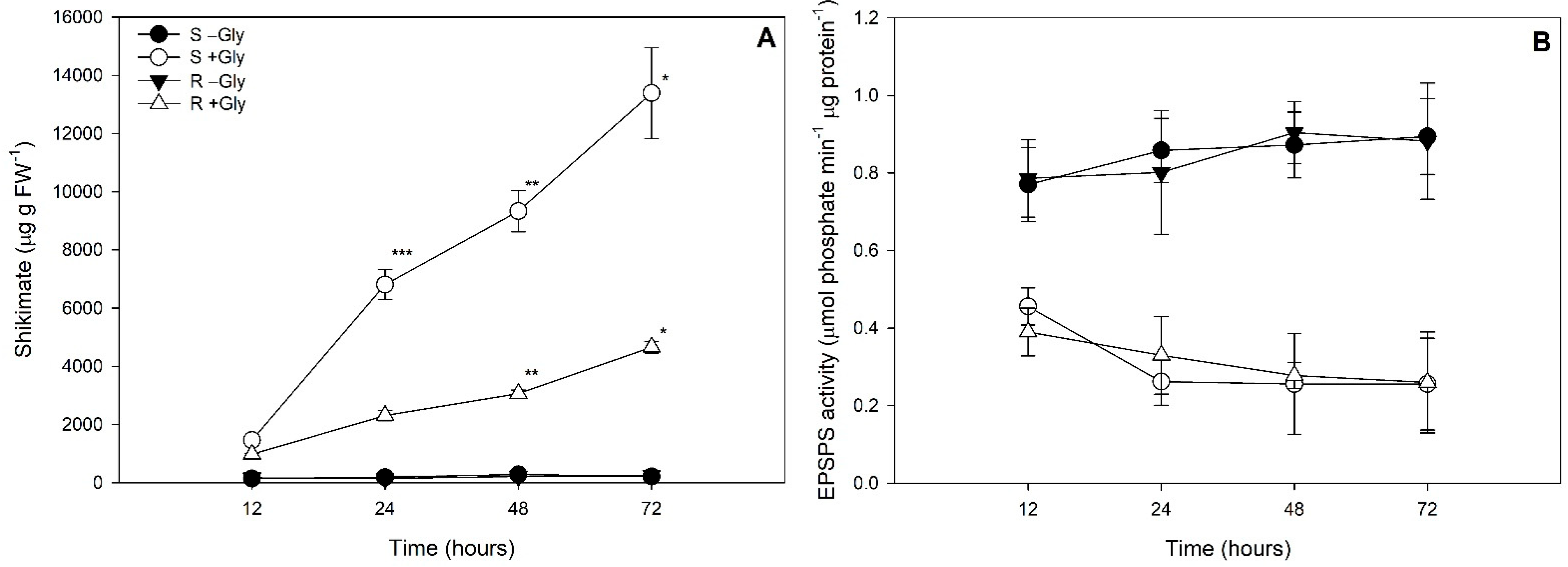
2.3. Shikimate Concentrations and EPSPS Activities
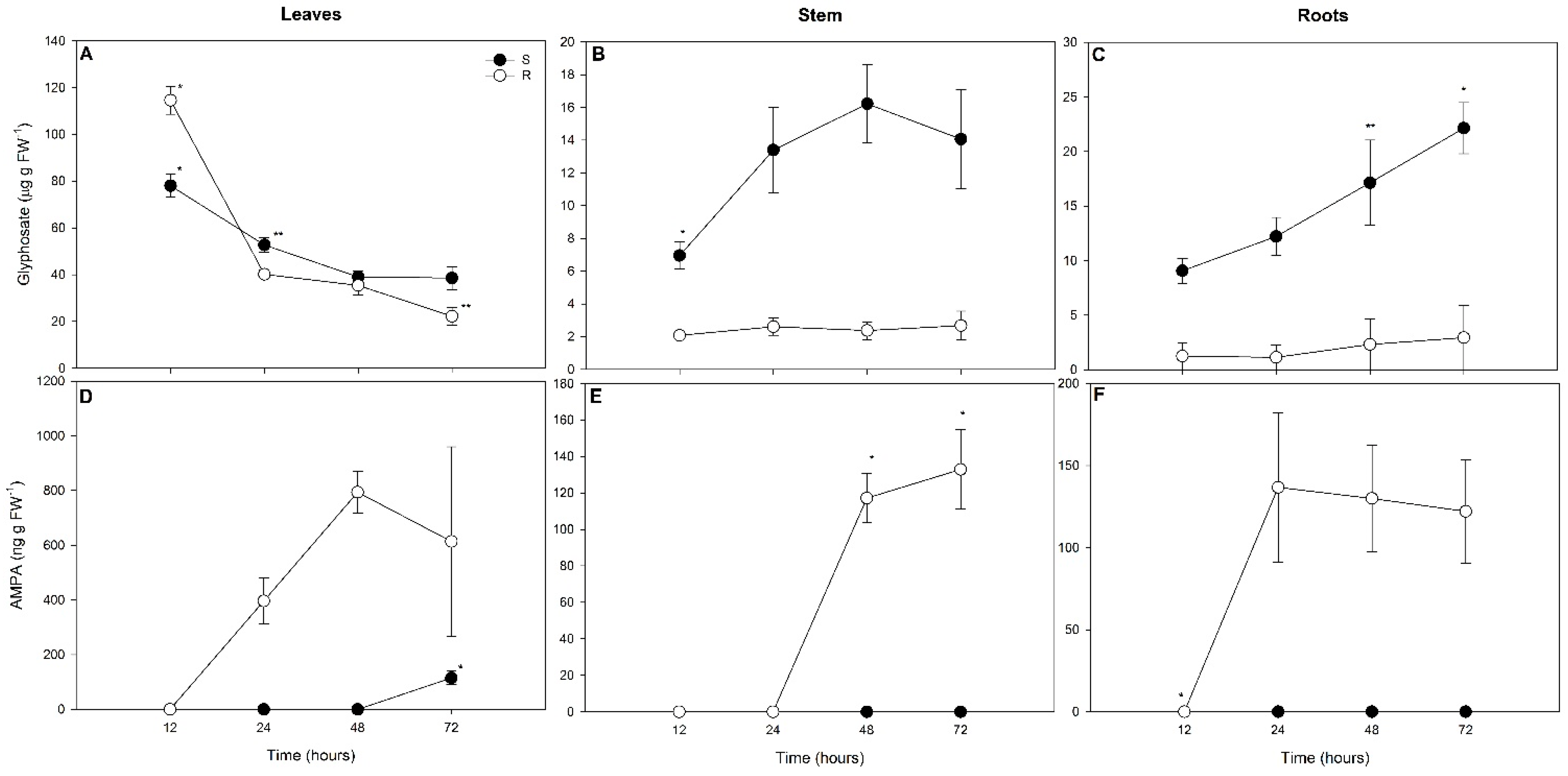
2.4. Glyphosate and AMPA Concentrations
3. Discussion
4. Material and Methods
4.1. Greenhouse Experiments
4.2. Enzymatic Evaluations of the Leaves
4.3. Glyphosate, AMPA and Shikimate Evaluations
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid (AMPA), an overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Siehl, D. Inhibitors of EPSPS synthase, glutamine synthetase and histidine synthesis. In Herbicide Activity: Toxicology, Biochemistry and Molecular Biology; Roe, R., Burton, J., Kuhr, R., Eds.; IOS Press: Amsterdam, The Netherlands, 1997; pp. 37–67. [Google Scholar]
- Ferreira Mendes, K.; Nogueira de Sousa, R.; Flávia Souza Laube, A.; Current Approaches to Pesticide Use and Glyphosate-Resistant Weeds in Brazilian Agriculture. Multifunctionality and Impacts of Organic and Conventional Agriculture IntechOpen. 2020. Available online: https://www.intechopen.com/books/multifunctionality-and-impacts-of-organic-and-conventional-agriculture/current-approaches-to-pesticide-use-and-glyphosate-resistant-weeds-in-brazilian-agriculture (accessed on 3 September 2022).
- Alcántara-de la Cruz, R.; Domínguez-Martínez, P.A.; da Silveira, H.M.; Cruz-Hipólito, H.E.; Palma-Bautista, C.; Vázquez-García, J.G.; Dominguez-Valenzuela, J.A.; de Prado, R. Management of glyphosate-resistant weeds in Mexican citrus groves: Chemical alternatives and economic viability. Plants 2019, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Smedbol, É.; Lucotte, M.; Labrecque, M.; Lepage, L.; Juneau, P. Phytoplankton growth and PSII efficiency sensitivity to a glyphosate-based herbicide (Factor 540®). Ecotoxicol. Environ. Saf. 2017, 192, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Smedbol, E.; Gomes, M.P.; Paquet, S.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Effects of low concentrations of glyphosate-based herbicide Factor 540® on an agricultural stream freshwater phytoplankton community. Chemosphere 2018, 192, 133–141. [Google Scholar] [CrossRef]
- Gomes, M.P.; dos Santos, M.P.; de Freitas, P.L.; Schafaschek, A.M.; de Barros, E.N.; Kitamura, R.S.A.; Paulete, V.; Navarro-Silva, M.A. The aquatic macrophyte Salvinia molesta mitigates herbicides (glyphosate and aminomethylphosphonic acid) effects to aquatic invertebrates. Environ. Sci. Pollut. Res. 2022, 1–14. Available online: https://link.springer.com/10.1007/s11356-022-23012-w (accessed on 16 September 2022). [CrossRef]
- Mendes, E.J.; Malage, L.; Rocha, D.C.; Kitamura, R.S.A.; Gomes, S.M.A.; Navarro-Silva, M.A.; Gomes, M.P. Isolated and combined effects of glyphosate and its by-product aminomethylphosphonic acid on the physiology and water remediation capacity of Salvinia molesta. J. Hazard. Mater. 2021, 417, 125694. [Google Scholar] [CrossRef]
- EMBRAPA. Resistência de Plantas Daninhas a Herbicidas Preocupa Agricultores. 2018. Available online: https://www.embrapa.br/busca-de-noticias/-/noticia/37661812/resistencia-de-plantas-daninhas-a-herbicidas-preocupa-agricultores (accessed on 2 May 2019).
- Powles, S.B. Evolved glyphosate-resistant weeds around the world: Lessons to be learnt. Am. J. Manag. Care. 2013, 19, 871–880. [Google Scholar] [CrossRef]
- Anagnostopoulos, C.; Stasinopoulou, P.; Kanatas, P.; Travlos, I. Differences in metabolism of three Conyza species to herbicides glyphosate and triclopyr revealed by LC-MS/MS. Child. J. Agric. Res. 2020, 80, 100–107. [Google Scholar] [CrossRef]
- Agostinetto, D.; Silva DRO da Vargas, L. Soybean yield loss and economic thresholds due to glyphosate resistant hairy fleabane interference. Arq. Inst. Biológico 2018, 84. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1808-16572017000100230&lng=en&tlng=en (accessed on 2 September 2022). [CrossRef]
- Gazziero, D.L.P.; Adegas, F.S.; Voll, E.; Vargas, L.; Karam, D.; Matallo, M.B. Interferência da buva em áreas cultivadas com soja. In Proceedings of the XXVII Congresso Brasileiro da Ciência das Plantas Daninhas, Ribeirão Preto, Brazil, 19–23 July 2010; Sociedade Brasileira de Plantas Daninhas: São Paulo, SP, Brazil; pp. 1555–1558. [Google Scholar]
- Ferreira, E.; Galon, L.; Aspiazú, I.; Silva, A.; Concenço, G.; Silva, A.; Oliveira, J.; Vargas, L. Glyphosate translocation in hairy fleabane (Conyza bonariensis) biotypes. Planta Daninha 2008, 26, 637–643. [Google Scholar] [CrossRef][Green Version]
- Vargas, L.; Silva, D.; Agostinetto, D.; Matallo, M.; Santos, F.; Almeida, S.; Chavarria, G. Glyphosate influence on the physiological parameters of Conyza bonariensis biotypes. Planta Daninha 2014, 32, 151–159. [Google Scholar] [CrossRef][Green Version]
- Hao, J.-H.; Qiang, S.; Liu, Q.-Q.; Cao, F. Reproductive traits associated with invasiveness in Conyza sumatrensis. J. Syst. Evol. 2009, 47, 245–254. [Google Scholar] [CrossRef]
- De Prado, J.L.; Osuna, M.D.; Heredia, A.A.; De Prado, R. Lolium rigidum, a Pool of Resistance Mechanisms to ACCase Inhibitor Herbicides. J. Agric. Food Chem. 2005, 53, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Huang, S.; Powles, S. Direct measurement of paraquat in leaf protoplasts indicates vacuolar paraquat sequestration as a resistance mechanism in Lolium rigidum. Pestic. Biochem. Physiol. 2010, 98, 104–109. [Google Scholar] [CrossRef]
- Busi, R.; Vila-Aiub, M.M.; Powles, S.B. Genetic control of a cytochrome P450 metabolism-based herbicide resistance mechanism in Lolium rigidum. Heredity 2011, 106, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef]
- Gomes, M.P.; Le Manac’H, S.G.; Moingt, M.; Smedbol, E.; Paquet, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Impact of phosphate on glyphosate uptake and toxicity in willow. J. Hazard. Mater. 2016, 304, 269–279. [Google Scholar] [CrossRef]
- Gomes, M.P.; Le Manac’h, S.G.; Maccario, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Differential effects of glyphosate and aminomethylphosphonic acid (AMPA) on photosynthesis and chlorophyll metabolism in willow plants. Pestic. Biochem. Physiol. 2016, 130, 65–70. [Google Scholar] [CrossRef]
- Lamb, D.C.; Kelly, D.E.; Hanley, S.Z.; Mehmood, Z.; Kelly, S.L. Glyphosate is an inhibitor of plant cytochrome P450: Functional expression of Thlaspi arvensae cytochrome P45071B1/reductase fusion protein in Escherichia coli. Biochem. Biophys. Res. Commun. 1998, 244, 110–114. [Google Scholar] [CrossRef]
- Gomes, M.P.; Tavares, D.S.; Richardi, V.S.; Marques, R.Z.; Wistuba, N.; Moreira de Brito, J.C.; Soffiatti, P.; Sant’Anne-Santos, B.F.; de Silva, M.A.N.; Juneau, P. Enrofloxacin and Roundup® interactive effects on the aquatic macrophyte Elodea canadensis physiology. Environ. Pollut. 2019, 249, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yu, Q.; Beffa, R.; González, S.; Maiwald, F.; Wang, J.; Powles, S.B. Cytochrome P450 CYP81A10v7 in Lolium rigidum confers metabolic resistance to herbicides across at least five modes of action. Plant J. 2020, 105, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu. Rev. Plant. Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Valenzuela, J.A.; Alcántara-de la Cruz, R.; Palma-Bautista, C.; Vázquez-García, J.G.; Cruz-Hipolito, H.E.; De Prado, R. Non-Target Site Mechanisms Endow Resistance to Glyphosate in Saltmarsh Aster (Aster squamatus). Plants 2021, 10, 1970. [Google Scholar] [CrossRef]
- Bracamonte, E.; Silveira HM da Alcántara-de la Cruz, R.; Domínguez-Valenzuela, J.A.; Cruz-Hipolito, H.E.; De Prado, R. From tolerance to resistance: Mechanisms governing the differential response to glyphosate in Chloris barbata. Pest. Manag. Sci. 2018, 74, 1118–1124. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of evolved herbicide resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Han, H.; Mao, L.; Nyporko, A.; Fan, L.; Bai, L.; Powles, S. Aldo-keto Reductase Metabolizes Glyphosate and Confers Glyphosate Resistance in Echinochloa colona. Plant Physiol. 2019, 181, 1519–1534. [Google Scholar] [CrossRef]
- Maroli, A.S.; Nandula, V.K.; Dayan, F.E.; Duke, S.O.; Gerard, P.; Tharayil, N. Metabolic Profiling and Enzyme Analyses Indicate a Potential Role of Antioxidant Systems in Complementing Glyphosate Resistance in an Amaranthus palmeri Biotype. J. Agric. Food Chem. 2015, 63, 9199–9209. [Google Scholar] [CrossRef]
- González-Torralva, F.; Rojano-Delgado, A.M.; Luque de Castro, M.D.; Mülleder, N.; De Prado, R. Two non-target mechanisms are involved in glyphosate-resistant horseweed (Conyza canadensis L. Cronq.) biotypes. J. Plant Physiol. 2012, 169, 1673–1679. [Google Scholar] [CrossRef]
- Smedbol, É.; Lucotte, M.; Maccario, S.; Gomes, M.P.M.P.; Paquet, S.; Moingt, M.; Mercier, L.L.C.; Sobarzo, M.R.P.; Blouin, M.-A. Glyphosate and aminomethylphosphonic acid content in glyphosate-resistant soybean leaves, stems and roots and associated phytotoxicity following a single glyphosate-based herbicide application. J. Agric. Food Chem. 2019, 67, 6133–6142. [Google Scholar] [CrossRef]
- Eceiza, M.V.; Gil-Monreal, M.; Barco-Antoñanzas, M.; Zabalza, A.; Royuela, M. The moderate oxidative stress induced by glyphosate is not detected in Amaranthus palmeri plants overexpressing EPSPS. J. Plant Physiol. 2022, 274, 153720. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Kitamura, R.S.A.; Marques, R.Z.; Barbato, M.L.; Zámocký, M. The Role of H2O2-Scavenging Enzymes (Ascorbate Peroxidase and Catalase) in the Tolerance of Lemna minor to Antibiotics: Implications for Phytoremediation. Antioxidants 2022, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef] [PubMed]
- Yanniccari, M.; Gigón, R.; Larsen, A. Cytochrome P450 Herbicide Metabolism as the Main Mechanism of Cross-Resistance to ACCase- and ALS-Inhibitors in Lolium spp. Populations from Argentina: A Molecular Approach in Characterization and Detection. Front. Plant Sci. 2020, 11, 600301. [Google Scholar] [CrossRef] [PubMed]
- Fathi, M.A.; Han, G.; Kang, R.; Shen, D.; Shen, J.; Li, C. Disruption of cytochrome P450 enzymes in the liver and small intestine in chicken embryos in ovo exposed to glyphosate. Environ. Sci. Pollut. Res. 2020, 27, 16865–16875. [Google Scholar] [CrossRef]
- Samsel, A.; Seneff, S. Glyphosate’s Suppression of Cytochrome P450 Enzymes and Amino Acid Biosynthesis by the Gut Microbiome: Pathways to Modern Diseases. Entropy 2013, 15, 1416–1463. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate degradation in glyphosate-resistant and -susceptible crops and weeds. J. Agric. Food Chem. 2011, 59, 5835–5841. [Google Scholar] [CrossRef]
- Coupland, D. The effect of temperature on the activity and metabolism of glyphosate applied to rhizome fragments of elymus repens (Agropyron repens). Pestic. Sci. 1984, 15, 226–234. [Google Scholar] [CrossRef]
- Eberbach, P.L.; Bowmer, K. Conversion of 14C-glyphosate to carbon dioxide by alligator weed. J. Aquat. Plant Manag. 1995, 33, 27–29. [Google Scholar]
- Xu, J.; Wang, X.; Guo, W. The cytochrome P450 superfamily: Key players in plant development and defense. J. Integr. Agric. 2015, 14, 1673–1686. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, W.; Li, Z.; Ma, L.; Yu, J.; Wang, H.; Liu, Z.; Xu, B. Identification and Characterization of Three New Cytochrome P450 Genes and the Use of RNA Interference to Evaluate Their Roles in Antioxidant Defense in Apis cerana cerana Fabricius. Front. Physiol. 2018, 9, 1608. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.; Ding, J.; Lee, D.; Lu, X.; Feng, Y.; Song, W. Overexpression of SoCYP85A1, a Spinach Cytochrome p450 Gene in Transgenic Tobacco Enhances Root Development and Drought Stress Tolerance. Front. Plant Sci. 2017, 8, 1909. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Maccario, S.; Lucotte, M.; Labrecque, M.; Juneau, P. Consequences of phosphate application on glyphosate uptake by roots: Impacts for environmental management practices. Sci. Total Environ. 2015, 537, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Richardi, V.S.; Bicalho, E.M.; da Rocha, D.C.; Navarro-Silva, M.A.; Soffiatti, P.; Gracia, Q.S.; Sant’Anna-Santos, B.F. Effects of Ciprofloxacin and Roundup on seed germination and root development of maize. Sci. Total Environ. 2019, 651, 2671–2678. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Vázquez-García, J.G.; Domínguez-Valenzuela, J.A.; Mendes, K.F.; de la Cruz, R.A.; Torra, J.; De Prado, R. Non-Target-Site Resistance Mechanisms Endow Multiple Herbicide Resistance to Five Mechanisms of Action in Conyza bonariensis. J. Agric. Food Chem. 2021, 69, 14792–14801. [Google Scholar] [CrossRef]
- Gomes, M.P.; Juneau, P. Temperature and light modulation of herbicide toxicity on algal and cyanobacterial physiology. Front. Environ. Sci. 2017, 5, 50. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Bio. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Guengerich, F.P.; Martin, M.V.; Sohl, C.D.; Cheng, Q.; Gasser, M.; Zhan, Q. Measurement of cytochrome P450 and NADPH-cytochrome P450 reductase. Nat. Protoc. 2013, 4, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest. Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Owens, D.K.; Corniani, N.; Silva, F.M.L.; Watson, S.B.; Howell, J.; Shaner, D.L. Biochemical Markers and Enzyme Assays for Herbicide Mode of Action and Resistance Studies. Weed Sci. 2015, 63, 23–63. [Google Scholar] [CrossRef]
- Gomes, G.L.G.C.; Carbonari, C.A.; Velini, E.D.; Trindade, M.L.B.; Silva, J.R.M. Extraction and Simultaneous Determination of Glyphosate, AMPA and Compounds of the Shikimic Acid Pathway in Plants. Planta Daninha 2015, 33, 295–304. [Google Scholar] [CrossRef]
- Matallo, M.B.; Almeida, S.D.B.; Franco, D.A.S.; Cerdeira, A.L.; Gazzeiro, D.L.P. Glyphosate as a tool to produce shikimic acid in plants. Planta Daninha 2014, 32, 601–608. [Google Scholar] [CrossRef][Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubis, G.C.; Marques, R.Z.; Kitamura, R.S.A.; Barroso, A.A.M.; Juneau, P.; Gomes, M.P. Antioxidant Enzyme and Cytochrome P450 Activities Are Involved in Horseweed (Conyza Sumatrensis) Resistance to Glyphosate. Stresses 2023, 3, 47-57. https://doi.org/10.3390/stresses3010005
Kubis GC, Marques RZ, Kitamura RSA, Barroso AAM, Juneau P, Gomes MP. Antioxidant Enzyme and Cytochrome P450 Activities Are Involved in Horseweed (Conyza Sumatrensis) Resistance to Glyphosate. Stresses. 2023; 3(1):47-57. https://doi.org/10.3390/stresses3010005
Chicago/Turabian StyleKubis, Gabrielly Cristina, Raizza Zorman Marques, Rafael Shinji Akiyama Kitamura, Arthur Arrobas Martins Barroso, Philippe Juneau, and Marcelo Pedrosa Gomes. 2023. "Antioxidant Enzyme and Cytochrome P450 Activities Are Involved in Horseweed (Conyza Sumatrensis) Resistance to Glyphosate" Stresses 3, no. 1: 47-57. https://doi.org/10.3390/stresses3010005
APA StyleKubis, G. C., Marques, R. Z., Kitamura, R. S. A., Barroso, A. A. M., Juneau, P., & Gomes, M. P. (2023). Antioxidant Enzyme and Cytochrome P450 Activities Are Involved in Horseweed (Conyza Sumatrensis) Resistance to Glyphosate. Stresses, 3(1), 47-57. https://doi.org/10.3390/stresses3010005