
Abscisic Acid in Coordination with Nitrogen Alleviates Salinity-Inhibited Photosynthetic Potential in Mustard by Improving Proline Accumulation and Antioxidant Activity





Abstract
1. Introduction
2. Results
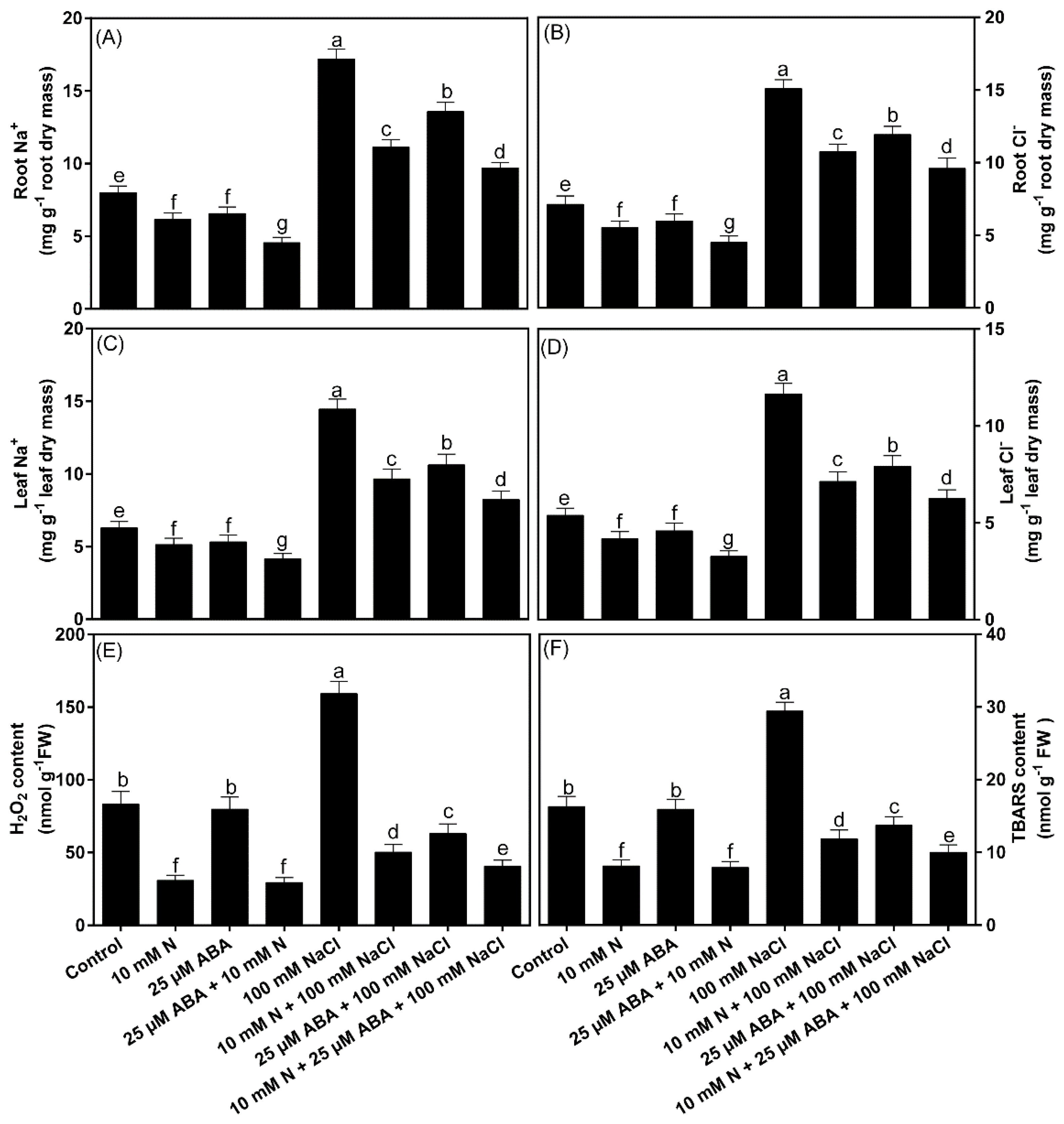
2.1. Impact of ABA and/or N on Accumulation of Na+ and Cl− Ions and Oxidative Stress
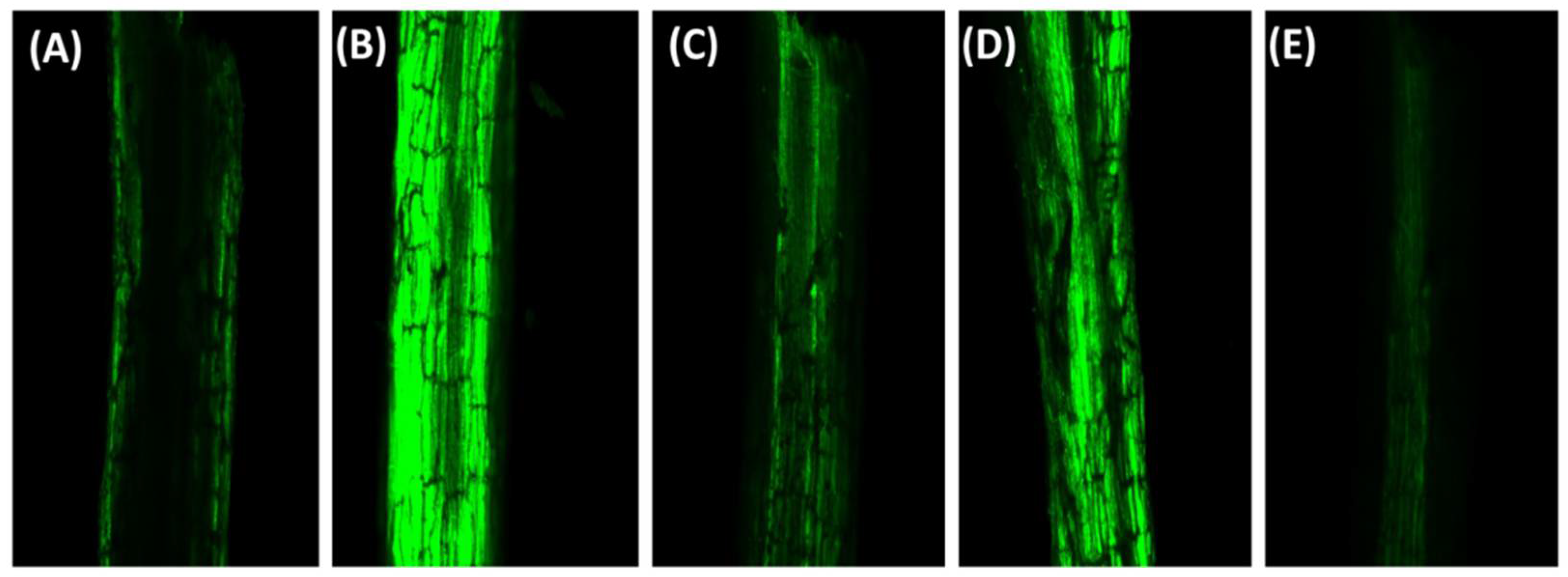
2.2. Visualization of the Status of O2− and H2O2 in Leaves Using Histochemical Staining and Confocal Laser Scanning Microscopy
2.3. Influence of ABA and/or N on Photosynthetic Characteristics under NaCl Stress
2.4. Impact of ABA and/or N on Growth under NaCl Stress
2.5. Impact of ABA and/or N on Proline Content under NaCl Stress
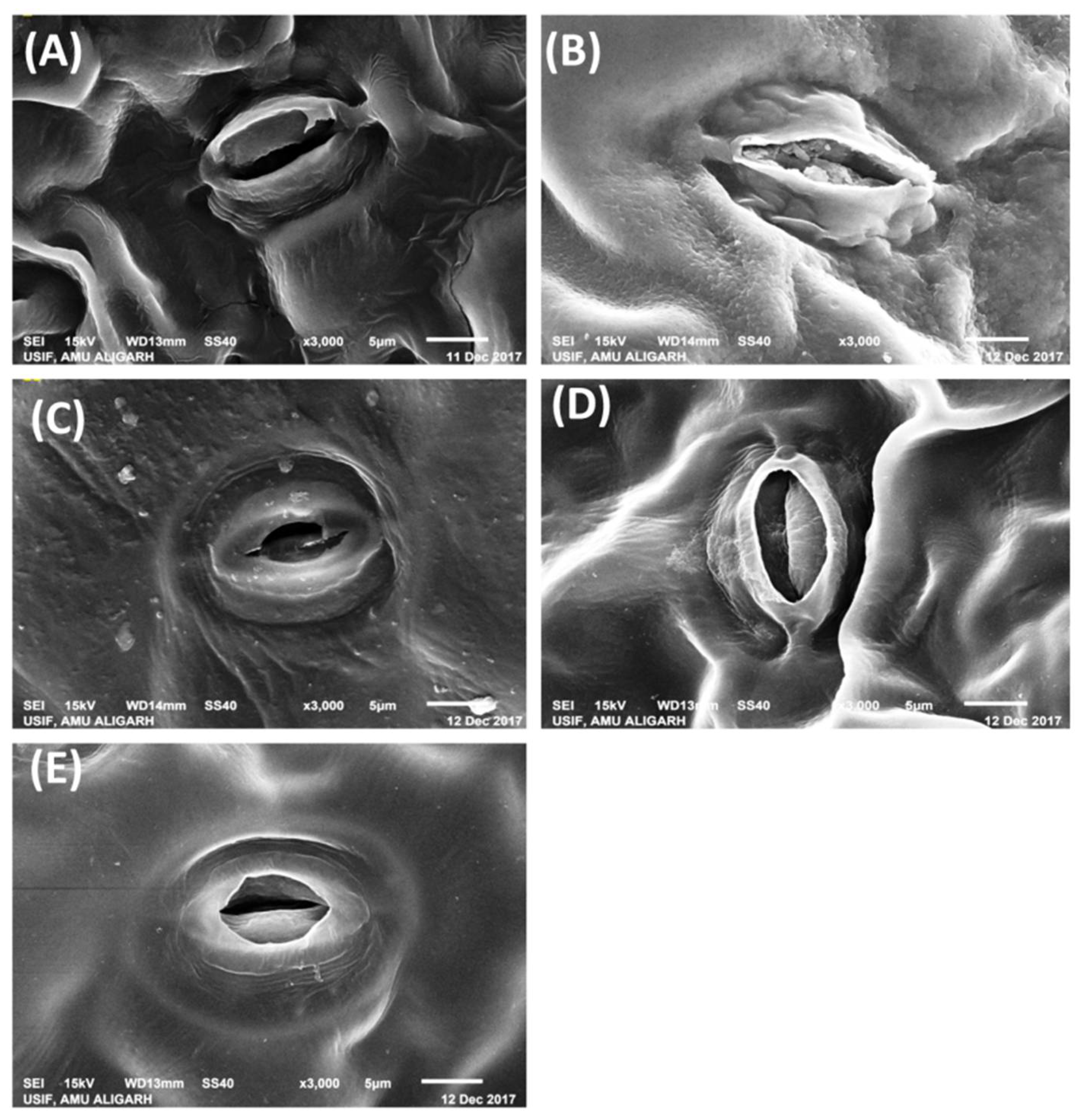
2.6. Impact ofABA and/or N on Stomatal Behavior under NaCl Stress
2.7. Impact of ABA and/or N on Antioxidant Metabolism under NaCl Stress
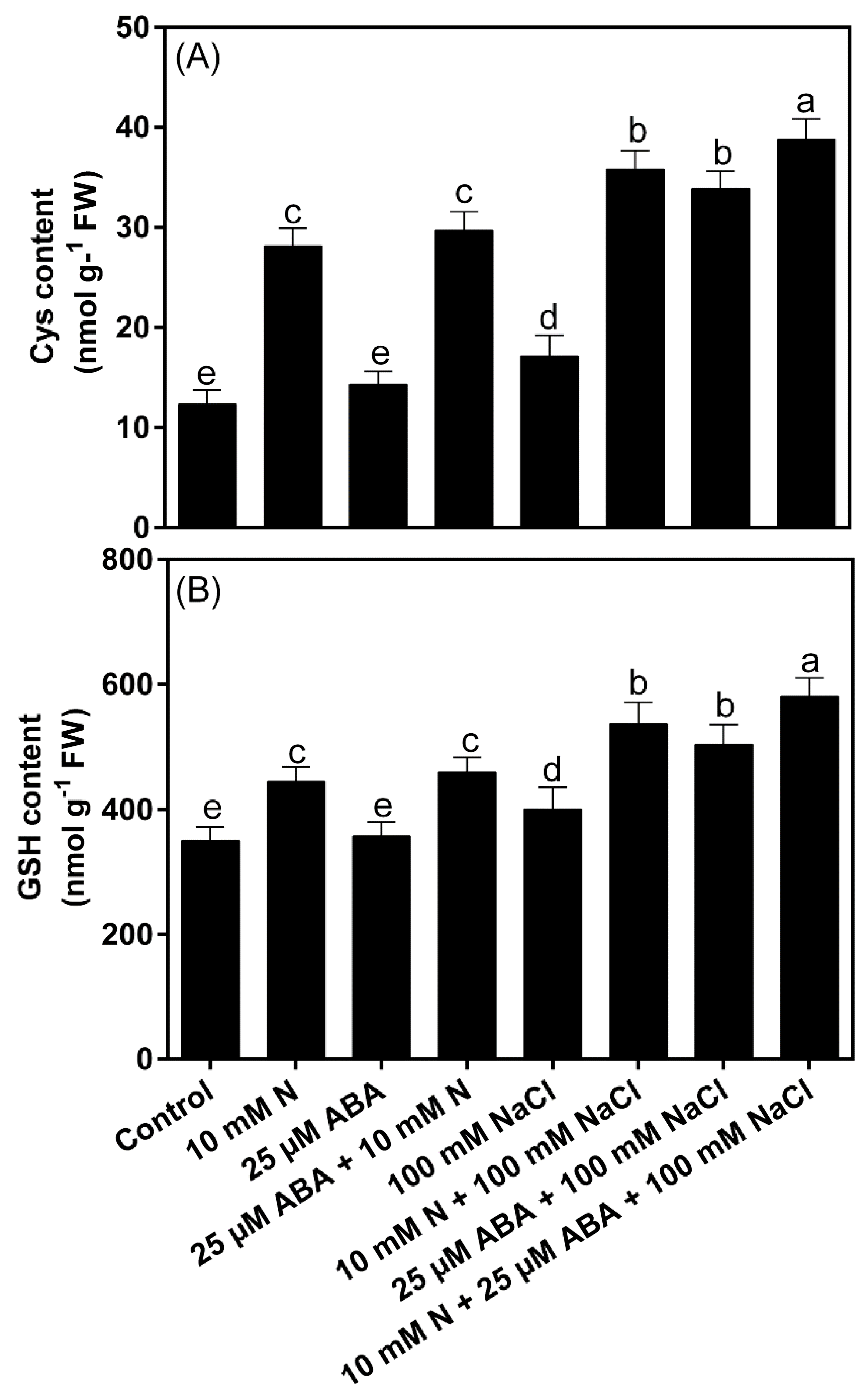
2.8. Impact of ABA and/or N on Cysteine and GSH Content under NaCl Stress
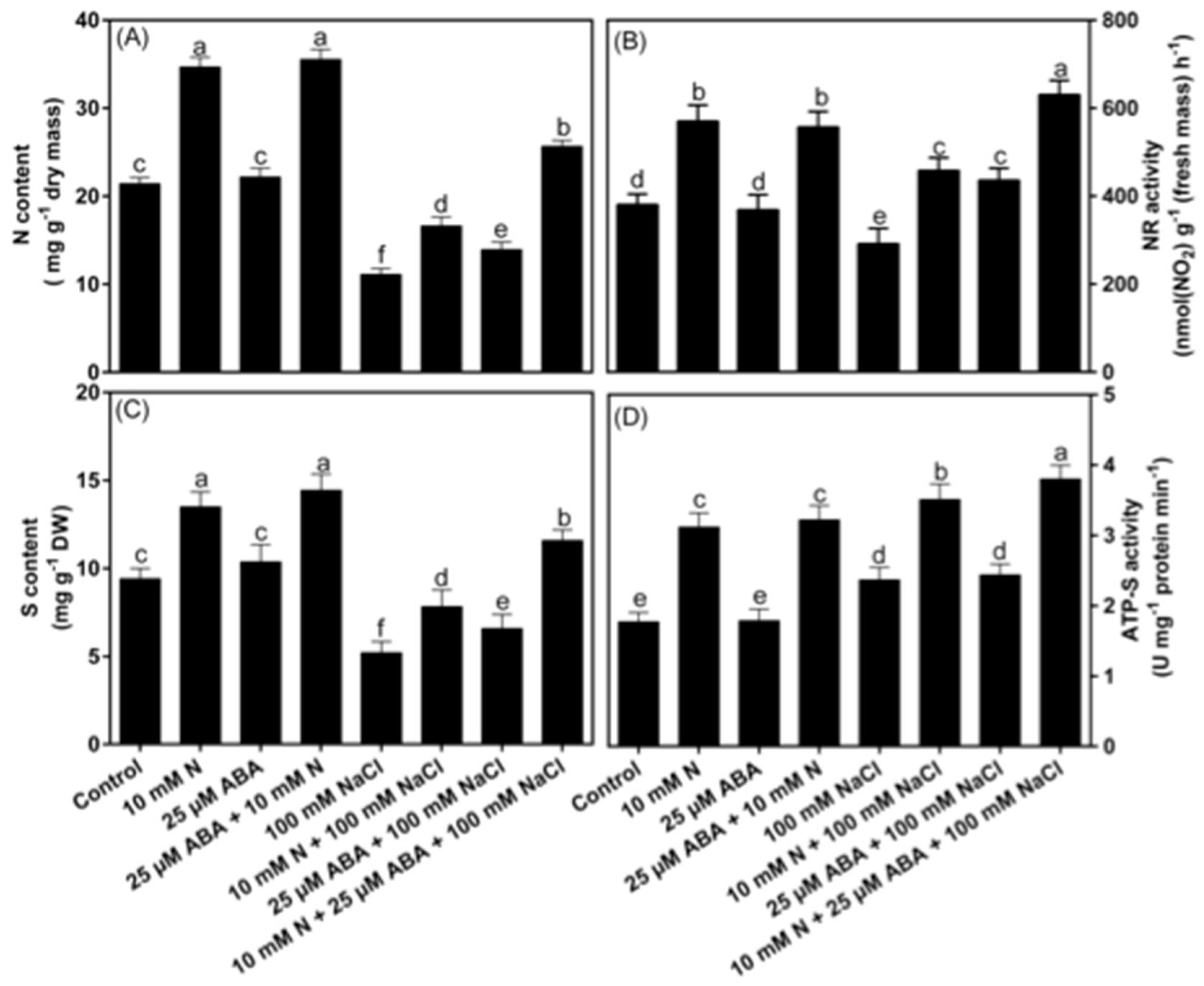
2.9. Effect of ABA and/or N on Contents of N, S and Activity of NR and ATP-S under NaCl
3. Discussion
3.1. Modulation of Ionic Toxicity (Na+ and Cl−) and Mineral Elements (N and S)-Accumulation, and the Status of Oxidative Stress and Antioxidant Metabolism
3.2. Control of the Status of N-Containing Osmolyte Proline and S-Containing Compounds (Cys and GSH), and Activity of NR and ATP-S
3.3. Overall Improvement in the Photosynthetic and Growth Parameters
4. Methodology
4.1. Experimental Design and Growth Conditions
4.2. Analyses of Na+ and Cl− Content
4.3. Content of H2O2 and TBARS
4.4. Superoxide Ion (O2−) and H2O2 by a Histochemical Staining Method
4.5. Analysis of H2O2 in Roots by Confocal Laser Scanning Microscopy
4.6. Photosynthetic Parameters and Rubisco Activity
4.7. Growth Parameters
4.8. Assay of Antioxidant Enzymes
4.9. Determination of Nitrate Reductase (NR) Activity and N Content
4.10. Determination of Proline Content
4.11. Analyses of Content of S, Cysteine, GSH, and ATP-S Activity
4.12. Analysis of Stomatal Behavior
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrol, D.P.; Shankar, U. Integrated pest management. In Breeding Oilseed Crops for Sustainable Production; Academic Press: Cambridge, MA, USA, 2016; pp. 523–549. [Google Scholar]
- Shekhawat, K.; Rathore, S.S.; Premi, O.P.; Kandpal, B.K.; Chauhan, J.S. Advances in agronomic management of Indian mustard (Brassicajuncea (L.) Czern & Coss.): An overview. Int. J. Agron. 2012, 2012, 408284. [Google Scholar] [CrossRef]
- Singh, K.H.; Shakya, R.; Mahawar, R.K. Genetic diversity and patterns of variation among Indian mustard (Brassica juncea (L.) Czern. & Coss.) varieties. J. Breed. Genet. 2014, 46, 329–339. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Management of Salt Affected Soils. FAO Soil Portal. Food and Agriculture Organization of the United Nations. 2021. Available online: http://www.fao.org/soils-portal/soil-management/management-of-some-problem-soils/salt-affected-soils/en/ (accessed on 16 August 2021).
- Sehgal, J.L.; Abrol, I.P. Soil Degradation in India: Status and Impact; Oxford & IBH Publishing Co.: Delhi, India, 1994. [Google Scholar]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycine betaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Agarwal, R.M. Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L) as influenced by potassium supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef]
- Rasheed, F.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. The key roles of salicylic acid and sulfur in plant salinity stress tolerance. J. Plant Growth Regul. 2020, 1–14. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Sofo, A.; Scopa, A.; Roychoudhury, A.; Gill, S.S.; Iqbal, M.; Lukatkin, A.S.; Pereira, E.; Duarte, A.C.; Ahmad, I. Lipids and proteins—Major targets of oxidative modifications in abiotic stressed plants. Environ. Sci. Pollut. Res. 2015, 22, 4099–4121. [Google Scholar] [CrossRef]
- Tuteja, N. Mechansims of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gill, S.S.; Gill, R. Cysteine—Jack of all glutathione-based plant stress defense trades. In Plant Adaptation to Environmental Change: Significance of Amino Acids and Their Derivatives; Anjum, N.A., Gill, S.S., Gill, R., Eds.; CABI: Wallingford, UK, 2014; pp. 35–52. [Google Scholar]
- Iqbal, N.; Umar, S.; Khan, N.A.; Khan, M.I.R. A new perspective of phytohormones in salinity tolerance: Regulation of proline metabolism. Environ. Exp. Bot. 2014, 100, 34–42. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- Gurmani, A.R.; Bano, A.; Ullah, N.; Khan, H.; Jahangir, M.; Flowers, T.J. Exogenous abscisic acid (ABA) and silicon (Si) promote salinity tolerance by reducing sodium (Na+) transport and bypass flow in rice (Oryza sativa indica). Aust. J. Crop Sci. 2013, 7, 1219–1226. [Google Scholar]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availability regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physiol. 2015, 178, 84–89. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gill, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, P.; Sharmila, P.; Saradhi, P.P. Proline suppresses rubisco activity by dissociating small subunits from holoenzyme. Biochem. Biophys. Res. Commun. 2001, 282, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Son, J.A.; Narayanankutty, D.P.; Roh, K.S. Influence of exogenous application of glutathione on rubisco and Rubiscoactivase in heavy metal-stressed tobacco plant grown in vitro. Saudi J. Biol. Sci. 2014, 21, 89–97. [Google Scholar] [CrossRef]
- Anjum, N.A.; Aref, I.M.; Duarte, A.C.; Pereira, E.; Ahmad, I.; Iqbal, M. Glutathione and proline can coordinately make plants withstand the joint attack of metal(loid) and salinity stresses. Front. Plant Sci. 2014, 5, 662. [Google Scholar] [CrossRef]
- Sripinyowanich, S.; Klomsakul, P.; Boonburapong, B.; Bangyeekhun, T.; Asami, T.; Gu, H.; Chadchawan, S. Exogenous ABA induces salt tolerance in Indica rice (Oryza sativa L.): The role of OsP5CS1 and OsP5CR gene expression during salt stress. Environ. Exp. Bot. 2013, 86, 94–105. [Google Scholar] [CrossRef]
- Gao, S.; Sun, W.; Li, Y.; Shi, Y.; Qi, X. Physiological and biochemical effects of exogenous salicylic acid (SA) and abscisic acid (ABA) on maize seedlings under salt stress. Mol. Breed. 2017, 15, 4159–4164. [Google Scholar]
- Sikder, R.K.; Wang, X.; Zhang, H.; Gui, H.; Dong, Q.; Jin, D.; Song, M. Nitrogen enhances salt tolerance by modulating the antioxidant defense system and osmoregulation substance content in Gossypiumhirsutum. Plants 2020, 9, 450. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M.; El-Sheikh, M.A.; Alatar, A.A.; Zhang, L. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef]
- Mandal, S.; Raju, R.; Kumar, A.; Kumar, P.; Sharma, P.C. Current status of research, technology response and policy needs of salt-affected soils in India–A Review. J. Indian Soc. Coast. Agric. Res. 2018, 36, 40–53. [Google Scholar]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.M.; Hafeez Laghari, A.; Mustafa Bhabhan, G.; Hussain Talpur, K.; Bhutto, T.A.; Wahocho, S.A.; Lashari, A.A. Role of nitrogen for plant growth and development: A review. Adv. Environ. Biol. 2016, 10, 209–219. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Nazar, R.; Iqbal, N.; Syeed, S.; Khan, N.A. Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J. Plant Physiol. 2011, 168, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Abdolzadeh, A.; Shima, K.; Lambers, H.; Chiba, K. Change in uptake, transport and accumulation of ions in Nerium oleander (Rosebay) as affected by different nitrogen sources and salinity. Ann. Bot. 2008, 102, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, J.; Wang, S.; Hüttermann, A.; Altman, A. Salt, nutrient uptake and transport, and ABA of Populuseuphratica; a hybrid in response to increasing soil NaCl. Trees 2001, 15, 186–194. [Google Scholar] [CrossRef]
- Hussain, S.J.; Khan, N.A.; Anjum, N.A.; Masood, A.; Khan, M.I.R. Mechanistic elucidation of salicylic acid and sulphur-induced defence systems, nitrogen metabolism, photosynthetic, and growth potential of mung-bean (Vignaradiata) under salt stress. J. Plant Growth Regul. 2021, 40, 1000–1016. [Google Scholar] [CrossRef]
- Syeed, S.; Sehar, Z.; Masood, A.; Anjum, N.A.; Khan, N.A. Control of Elevated Ion Accumulation, Oxidative Stress, and Lipid Peroxidation with Salicylic Acid-Induced Accumulation of Glycine Betaine in Salinity Exposed Vignaradiata L. Appl. Biochem. Biotechnol. 2021, 1–20. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Bor, M.; Özdemir, F.; Türkan, I. The effect of salt stress on lipid peroxidation and antioxidants in leaves of sugar beet Beta vulgaris L. and wild beet Beta maritima L. Plant Sci. 2003, 164, 77–84. [Google Scholar] [CrossRef]
- Rather, B.A.; Mir, I.R.; Sehar, Z.; Anjum, N.A.; Masood, A.; Khan, N.A. The outcomes of the functional interplay of nitric oxide and hydrogen sulfide in metal stress tolerance in plants. Plant Physiol. Biochem. 2020, 155, 523–534. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. The role of endogenous nitric oxide in salicylic acid-induced up-regulation of ascorbate-glutathione cycle involved in salinity tolerance of pepper (Capsicum annuum L.) plants. Plant Physiol. Biochem. 2020, 147, 10–20. [Google Scholar] [CrossRef]
- Bray, E.A. Response to abiotic stress. Physiol. Mol. Biol. Plants 2000, 1158–1203. [Google Scholar]
- Borella, J.; Becker, R.; Lima, M.C.; Oliveira, D.D.S.C.D.; Braga, E.J.B.; Oliveira, A.C.B.D.; Amarante, L.D. Nitrogen source influences the antioxidative system of soybean plants under hypoxia and re-oxygenation. Sci. Agric. 2019, 76, 51–62. [Google Scholar] [CrossRef]
- Giraudat, J.; Parcy, F.; Bertauche, N.; Gosti, F.; Leung, J.; Morris, P.C.; Vartanian, N. Current advances in abscisic acid action and signalling. Plant Mol. Biol. 1994, 26, 1557–1577. [Google Scholar] [CrossRef]
- Merlot, S.; Giraudat, J. Genetic analysis of abscisic acid signal transduction. Plant Physiol. 1997, 114, 751. [Google Scholar] [CrossRef]
- Leung, J.; Giraudat, J. Abscisic acid signal transduction. Annu. Rev. Plant Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Gill, R.; Yadav, S.; Hassanuzzaman, M.; Fujita, M.; Mishra, P.; Sabat, S.C.; Tuteja, N. Superoxide dismutase–mentor of abiotic stress olerace in crop plants. Environ. Sci. Pollut. Res. 2015, 22, 10375–10394. [Google Scholar]
- Wangeline, A.L.; Burkhead, J.L.; Hale, K.L.; Lindblom, S.D.; Terry, N.; Pilon, M.; Pilon-Smits, E.A. Overexpression of ATP sulfurylase in Indian mustard: Effects on tolerance and accumulation of twelve metals. J. Environ. Qual. 2004, 33, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Alam, M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (Oryza sativa L.) varieties. BioMed Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [PubMed]
- Aleksza, D.; Horváth, G.V.; Sándor, G.; Szabados, L. Proline accumulation is regulated by transcription factors associated with phosphate starvation. Plant Physiol. 2017, 175, 555–567. [Google Scholar] [CrossRef]
- Hawkesford, M.; Hoefgen, R.; Galili, G.; Amir, R.; Angenon, G.; Hesse, H.; Rentsch, D.; Schaller, J.; Van der Meer, I.; Rouster, J.; et al. Optimising nutritional quality of crops. In Plant Genetic Engineering; Jaiwal, P.K., Ed.; Plant Metabolic Engineering and Molecular Farming; Studium Press LLC: Houston, TX, USA, 2006; Volume 7, pp. 85–116. [Google Scholar]
- Brunold, C.; Suter, M. Regulation of sulfate assimilation by nitrogen nutrition in the duckweed Lemna minor L. Plant Physiol. 1984, 76, 579–583. [Google Scholar] [CrossRef]
- Jamal, A.; Fazli, I.S.; Ahmad, S.; Kim, K.T.; Oh, D.G.; Abdin, M.Z. Effect of sulfur on nitrate reductase and ATP sulfurylase activities in groundnut (Arachis hypogea L.). J. Plant Biol. 2006, 49, 513–517. [Google Scholar] [CrossRef]
- Akram, S.; Siddiqui, M.N.; Hussain, B.N.; Al Bari, M.A.; Mostofa, M.G.; Hossain, M.A.; Tran, L.S.P. Exogenous glutathione modulates salinity tolerance of soybean [Glycine max (L.) Merrill] at reproductive stage. J. Plant Growth Regul. 2017, 36, 877–888. [Google Scholar] [CrossRef]
- Cao, F.; Cai, Y.; Liu, L.; Zhang, M.; He, X.; Zhang, G.; Wu, F. Differences in photosynthesis, yield and grain cadmium accumulation as affected by exogenous cadmium and glutathione in the two rice genotypes. Plant Growth Regul. 2015, 75, 715–723. [Google Scholar] [CrossRef]
- Ding, X.; Jiang, Y.; He, L.; Zhou, Q.; Yu, J.; Hui, D.; Huang, D. Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci. Rep. 2016, 6, 35424. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Hwang, S.; Mel Lytle, C.; Zhu, Y.; Tai, J.C.; Bravo, R.C.; Chen, Y.; Leustek, T.; Terry, N. Overexpression of ATP sulfurylase in Indian mustard leads to increased selenate uptake, reduction, and tolerance. Plant Physiol. 1999, 119, 123–132. [Google Scholar] [CrossRef]
- Bashir, H.; Ahmad, J.; Bagheri, R.; Nauman, M.; Qureshi, M.I. Limited sulfur resource forces Arabidopsis thaliana to shift towards non-sulfur tolerance under cadmium stress. Environ. Exp. Bot. 2013, 94, 19–32. [Google Scholar] [CrossRef]
- Turan, M.A.; Katkat, V.; Taban, S. Salinity-Induced stomatal resistance, proline, chlorophyll and. Int. J. Agric. Res. 2007, 2, 483–488. [Google Scholar]
- Taffouo, V.D.; Kouamou, J.K.; Ngalangue, L.M.T.; Ndjeudji, B.A.N.; Akoa, A. Effects of salinity stress on growth, ions partitioning and yield of some cowpea (Vigna unguiculata L. Walp.) cultivars. Int. J. Bot. 2009, 5, 135–143. [Google Scholar]
- Memon, S.A.; Hou, X.; Wang, L.J. Morphlogical analysis of salt stress response of pakchoi. Electron. J. Environ. Agric. Food Chem. 2010, 9, 248–254. [Google Scholar]
- Kumar, V.; Shriram, V.; Kishor, P.K.; Jawali, N.; Shitole, M.G. Enhanced proline accumulation and salt stress tolerance of transgenic Indica rice by over-expressing P5CSF129A gene. Plant Biotechnol. Rep. 2010, 4, 37–48. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Sudmalis, D.; Millah, S.K.; Gagliano, M.C.; Butré, C.I.; Plugge, C.M.; Rijnaarts, H.H.M.; Temmink, H. The potential of osmolytes and their precursors to alleviate osmotic stress of anaerobic granular sludge. Water Res. 2018, 147, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Fatma, M.; Asgher, M.; Masood, A.; Khan, N.A. Excess sulfur supplementation improves photosynthesis and growth in mustard under salt stress through increased production of glutathione. Environ. Exp. Bot. 2014, 107, 55–63. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Zeevaart, J.A. Abscisic acid biosynthesis and metabolism. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 137–155. [Google Scholar]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition. In Commonwealth Agricultural Bureaux, 2nd ed.; Farnham Royal: Bucks, UK, 1966; p. 547. [Google Scholar]
- Okuda, T.; Matsuda, Y.; Yamanaka, A.; Sagisaka, S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991, 97, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B.; Biosciences, J.M.I. Histochemical detection of superoxide and H2O2 accumulation in Brassica juncea seedlings. Bio-Protocol 2014, 4, e1108. [Google Scholar] [CrossRef]
- Usuda, H. The activation state of ribulose 1, 5-bisphosphate carboxylase in maize leaves in dark and light. Plant Cell Physiol. 1985, 26, 1455–1463. [Google Scholar]
- Asada, K. Ascorbate peroxidase–a hydrogen peroxide-scavenging enzyme in plants. Physiol. Plant 1992, 85, 235–241. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Kuo, T.M.; Warner, R.L.; Kleinhofs, A. In vitro stability of nitrate reductase from barley leaves. Phytochemistry 1982, 21, 531–533. [Google Scholar] [CrossRef]
- Nakagawa, H.; Poulle, M.; Oaks, A. Characterization of nitrate reductase from corn leaves (Zea mays cv W64A × W182E): Two molecular forms of the enzyme. Plant Physiol. 1984, 75, 285–289. [Google Scholar] [CrossRef]
- Lindner, R.C. Rapid analytical methods for some of the more common inorganic constituents of plant tissues. Plant Physiol. 1944, 19, 76. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Gaitonde, M.K. A spectrophotometric method for the direct determination of cysteine in the presence of other naturally occurring amino acids. Biochem. J. 1967, 104, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol. 1985, 113, 548–555. [Google Scholar] [PubMed]
- Lappartient, A.G.; Touraine, B. Demand-driven control of root ATP sulfurylase activity and SO42− uptake in intact canola (the role of phloem-translocated glutathione). Plant Physiol. 1996, 111, 147–157. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments/Parameters | Control | NaCl (100 mM) | ABA Concentration (µM) | 5 µM ABA + 100 mM NaCl | 10 µM ABA + 100 mM NaCl | 25 µM ABA + 100 mM NaCl | 50 µM ABA + 100 mM NaCl | |||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 10 | 25 | 50 | |||||||
| Chlorophyllcontent | 36.2 ± 1.15 a | 18.2 ± 0.58 f | 34.2 ± 1.08 a | 31.1 ± 0.98 b | 24.5 ± 0.78 d | 19.3 ± 0.61 f | 20.0 ± 0.63 ef | 22.0 ± 0.71 e | 28.5 ± 0.91 c | 25.0 ± 0.79 d |
| Net photosynthesis | 13.7 ± 0.44 a | 6.2 ± 0.19 f | 13.4 ± 0.43 a | 12.3 ± 0.39 b | 10.13 ± 0.33 c | 6.9 ± 0.22 f | 7.9 ± 0.25 e | 9.14 ± 0.29 d | 12.9 ± 0.41 ab | 9.1 ± 0.29 d |
| Stomatal conductance | 238.5 ± 6.23 a | 139.4 ± 6.04 e | 230.2 ± 6.01 ab | 219.6 ± 5.74 b | 195 ± 5.09 b | 142.6 ± 6.18 c | 170 ± 4.44 e | 182 ± 4.75 cd | 217 ± 5.67 b | 193 ± 5.04 c |
| Intercellular CO2 concentration | 294.9 ± 9.40 a | 161.6 ± 7.01 f | 288.5 ± 9.19 a | 278.2 ± 8.87 b | 242.8 ± 6.34 c | 166 ± 7.20 f | 180.2 ± 7.82 e | 192.2 ± 8.34 d | 257 ± 6.72 ab | 201 ± 9.88 d |
| Leaf area | 116.64 ± 3.05 a | 65 ± 2.28 gh | 114.4 ± 2.99 a | 106.02 ± 2.77 b | 77.8 ± 2.48 ef | 61.4 ± 2.86 h | 82.6 ± 2.63 de | 88.8 ± 2.83 d | 97.64 ± 3.11 c | 72.14 ± 2.3 fg |
| Plant dry mass | 2.2 ± 0.07 a | 1.31 ± 0.05 e | 1.78 ± 0.06 cd | 1.8 ± 0.05 cd | 1.94 ± 0.05 bc | 1.39 ± 0.06 e | 1.63 ± 0.07 d | 1.71 ± 0.07 d | 2.101 ± 0.05 ab | 1.33 ± 0.06 e |
| H2O2 content | 89.92 ± 4.42 c | 157.8 ± 4.03 a | 87.2 ± 4.28 c | 86.4 ± 4.24 c | 85.2 ± 4.18 c | 84.7 ± 2.57 c | 119.6 ± 3.62 b | 110.2 ± 4.78 b | 64.23 ± 3.15 d | 87.32 ± 4.29 c |
| TBARS content | 17.5 ± 0.86 c | 28.2 ± 0.72 a | 16.2 ± 0.79 c | 15.9 ± 0.78 c | 16.9 ± 0.83 c | 15.7 ± 0.47 c | 21.43 ± 0.65 b | 20.35 ± 0.88 b | 13.0 ± 0.63 d | 17.2 ± 0.84 c |
| Treatments/ Parameters | Net Photosynthesis | Stomatal Conductance | Intercellular CO2 Concentration | Chlorophyll Content | Rubisco Activity | Proline Content | Leaf Area | Plant Dry Mass |
|---|---|---|---|---|---|---|---|---|
| Control | 13.72 ± 0.59 e | 241.32 ± 7.69 e | 278.23 ± 8.87 e | 28.8 ± 1.2498 e | 49.11 ± 1.56 e | 7.32 ± 0.32 e | 113.36 ± 5.27 e | 2.5 ± 0.08 e |
| 100 mM NaCl | 6.3 ± 0.27 h | 147.42 ± 6.39 h | 169.72 ± 7.36 h | 16.4 ± 0.7117 h | 16.14 ± 1.02 h | 9.56 ± 0.41 d | 61.7 ± 3.74 h | 1.12 ± 0.07 h |
| 10 mM N | 21.2 ± 0.67 a | 339.62 ± 8.87 a | 388.21 ± 10.14 a | 42.3 ± 1.3487 a | 79.20 ± 2.07 a | 12.85 ± 0.41 c | 182.24 ± 5.81 a | 4.15 ± 0.08 a |
| 25 μM ABA | 10.54 ± 0.33 g | 194.52 ± 6.20 g | 223.23 ± 7.12 g | 22.3 ± 0.7110 g | 33.94 ± 1.24 g | 13.60 ± 0.41 bc | 78.92 ± 3.67 g | 1.78 ± 0.08 g |
| 25 μM ABA + 10 mM N | 17.9 ± 0.57 c | 292.8 ± 7.65 c | 333.25 ± 8.71 c | 35.5 ± 1.1319 c | 62.60 ± 1.64 c | 13.09 ± 0.42 c | 147.8 ± 6.03 c | 3.32 ± 0.10 c |
| 10 mM N + 100 mM NaCl | 16.3 ± 0.51 d | 269.4 ± 8.58 d | 305.73 ± 9.74 d | 32.2 ± 1.0267 d | 55.96 ± 1.78 d | 15.74 ± 0.50 ab | 130.58 ± 6.07 d | 2.98 ± 0.09 d |
| 25 μM ABA + 100 mM NaCl | 12.13 ± 0.38 f | 217.92 ± 6.94 f | 250.77 ± 7.99 f | 25.5 ± 0.8130 f | 44.02 ± 1.40 f | 14.32 ± 0.46 b | 96.14 ± 4.47 f | 2.21 ± 0.10 f |
| 25 μM ABA + 10 mM N + 100 mM NaCl | 19.6 ± 0.62 b | 316.2 ± 8.26 b | 360.69 ± 9.42 b | 38.93 ± 1.2412 b | 71.21 ± 1.86 b | 17.21 ± 0.55 a | 165.02 ± 5.26 b | 3.61 ± 0.11 b |
| Treatments/ Parameters | APX | GR | SOD |
|---|---|---|---|
| (U mg−1 Protein min−1) | |||
| Control | 0.634 ± 0.018 e | 0.178 ± 0.005 e | 4.54 ± 0.108 e |
| 100 mM NaCl | 0.845 ± 0.024 d | 0.273 ± 0.008 d | 6.46 ± 0.153 d |
| 10 mM N | 1.146 ± 0.027 c | 0.345 ± 0.008 c | 7.06 ± 0.168 c |
| 25 μM ABA | 0.676 ± 0.019 e | 0.195 ± 0.005 e | 4.74 ± 0.113 e |
| 25 μM ABA + 10 mM N | 1.188 ± 0.028 c | 0.358 ± 0.008 c | 7.29 ± 0.173 c |
| 10 mM N + 100 mM NaCl | 1.435 ± 0.034 b | 0.405 ± 0.009 b | 8.12 ± 0.193 b |
| 25 μM ABA + 100 mM NaCl | 1.401 ± 0.033 b | 0.386 ± 0.009 b | 7.89 ± 0.188 b |
| 25 μM ABA + 10 mM N + 100 mM NaCl | 1.790 ± 0.036 a | 0.482 ± 0.009 a | 8.72 ± 0.207 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majid, A.; Rather, B.A.; Masood, A.; Sehar, Z.; Anjum, N.A.; Khan, N.A. Abscisic Acid in Coordination with Nitrogen Alleviates Salinity-Inhibited Photosynthetic Potential in Mustard by Improving Proline Accumulation and Antioxidant Activity. Stresses 2021, 1, 162-180. https://doi.org/10.3390/stresses1030013
Majid A, Rather BA, Masood A, Sehar Z, Anjum NA, Khan NA. Abscisic Acid in Coordination with Nitrogen Alleviates Salinity-Inhibited Photosynthetic Potential in Mustard by Improving Proline Accumulation and Antioxidant Activity. Stresses. 2021; 1(3):162-180. https://doi.org/10.3390/stresses1030013
Chicago/Turabian StyleMajid, Arif, Bilal A. Rather, Asim Masood, Zebus Sehar, Naser A. Anjum, and Nafees A. Khan. 2021. "Abscisic Acid in Coordination with Nitrogen Alleviates Salinity-Inhibited Photosynthetic Potential in Mustard by Improving Proline Accumulation and Antioxidant Activity" Stresses 1, no. 3: 162-180. https://doi.org/10.3390/stresses1030013
APA StyleMajid, A., Rather, B. A., Masood, A., Sehar, Z., Anjum, N. A., & Khan, N. A. (2021). Abscisic Acid in Coordination with Nitrogen Alleviates Salinity-Inhibited Photosynthetic Potential in Mustard by Improving Proline Accumulation and Antioxidant Activity. Stresses, 1(3), 162-180. https://doi.org/10.3390/stresses1030013