
Amplification in Time and Dilution in Space: Partitioning Spatiotemporal Processes to Assess the Role of Avian-Host Phylodiversity in Shaping Eastern Equine Encephalitis Virus Distribution


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Domain and Virus Data
2.2. Climate Data
2.3. Avian-Host Occurrence and Phylodiversity
2.4. Statistical Analysis
2.5. Model Evaluation and Comparison
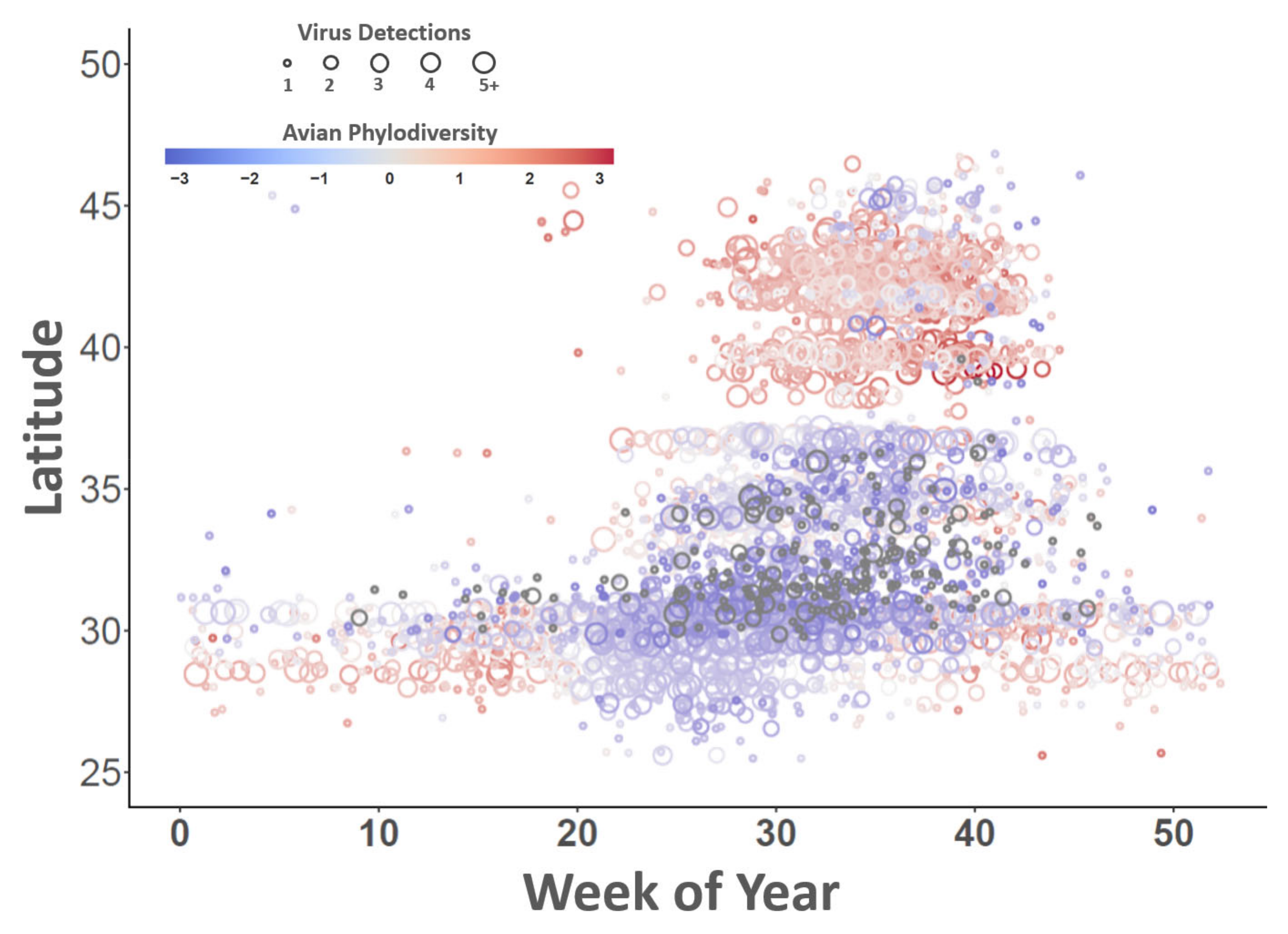
3. Results
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armstrong, P.M.; Andreadis, T.G. Eastern equine encephalitis virus—Old enemy, new threat. N. Engl. J. Med. 2013, 368, 1670–1673. [Google Scholar] [CrossRef] [PubMed]
- Burkett-Cadena, N.D.; Day, J.F.; Unnasch, T.R. Ecology of Eastern Equine Encephalitis Virus in the Southeastern United States: Incriminating Vector and Host Species Responsible for Virus Amplification, Persistence, and Dispersal. J. Med. Entomol. 2021, 59, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Sedda, L.; Brown, H.E. Predicting eastern equine encephalitis spread in North America: An ecological study. Curr. Res. Parasitol.-Vector-Borne Dis. 2021, 1, 100064. [Google Scholar] [CrossRef] [PubMed]
- Animal and Plant Health Inspection Service (APHIS). Equine Encephalitis. 2021. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/animal-disease-information/equine/eee-wee-vee (accessed on 14 December 2021).
- Villari, P.; Spielman, A.; Komar, N.; McDowell, M.; Timperi, R.J. The Economic Burden Imposed by a Residual Case of Eastern Encephalitis. Am. J. Trop. Med. Hyg. 1995, 52, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Andreadis, T.G. Ecology and Epidemiology of Eastern Equine Encephalitis Virus in the Northeastern United States: An Historical Perspective. J. Med. Entomol. 2021, 59, 1–13. [Google Scholar] [CrossRef]
- Davis, W.A. A study of birds and mosquitoes as hosts for the virus of eastern equine encephalomyelitis. Am. J. Hyg. 1940, 32, 45–59. [Google Scholar]
- Scott, T.W.; Weaver, S.C. Eastern equine encephalomyelitis virus: Epidemiology and evolution of mosquito transmission. Adv. Virus Res. 1989, 37, 277–328. [Google Scholar]
- Komar, N.; Dohm, D.J.; Turell, M.J.; Spielman, A. Eastern equine encephalitis virus in birds: Relative competence of European starlings (Sturnus vulgaris). Am. J. Trop. Med. Hyg. 1999, 60, 387–391. [Google Scholar] [CrossRef]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef]
- Alatoom, A.; Payne, D. An Overview of Arboviruses and Bunyaviruses. Lab. Med. 2009, 40, 237–240. [Google Scholar] [CrossRef]
- Go, Y.Y.; Balasuriya, U.B.R.; Lee, C.-k. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58–77. [Google Scholar] [CrossRef]
- Wong, K.T. Alphaviral Equine Encephalomyelitis (Eastern, Western, and Venezuelan). In Infections of the Central Nervous System; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Stenkamp-Strahm, C.; Patyk, K.; McCool-Eye, M.J.; Fox, A.; Humphreys, J.; James, A.; South, D.; Magzamen, S. Using geospatial methods to measure the risk of environmental persistence of avian influenza virus in South Carolina. Spat. Spatio Temporal Epidemiol. 2020, 34, 100342. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Ramey, A.M.; Douglas, D.C.; Mullinax, J.M.; Soos, C.; Link, P.; Walther, P.; Prosser, D.J. Waterfowl occurrence and residence time as indicators of H5 and H7 avian influenza in North American Poultry. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Ramey, A.M.; Reeves, A.B.; Drexler, J.Z.; Ackerman, J.T.; De La Cruz, S.; Lang, A.S.; Leyson, C.; Link, P.; Prosser, D.J.; Robertson, G.J.; et al. Influenza A viruses remain infectious for more than seven months in northern wetlands of North America. Proc. R. Soc. Biol. Sci. 2020, 287, 20201680. [Google Scholar] [CrossRef]
- Tolsá, M.J.; García-Peña, G.E.; Rico-Chávez, O.; Roche, B.; Suzán, G. Macroecology of birds potentially susceptible to West Nile virus. Proc. R. Soc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Pelzel-McCluskey, A.M.; Cohnstaedt, L.W.; McGregor, B.L.; Hanley, K.A.; Hudson, A.R.; Young, K.I.; Peck, D.; Rodriguez, L.L.; Peters, D.P.C. Integrating Spatiotemporal Epidemiology, Eco-Phylogenetics, and Distributional Ecology to Assess West Nile Disease Risk in Horses. Viruses 2021, 13, 1811. [Google Scholar] [CrossRef]
- Suzán, G.; García-Peña, G.E.; Castro-Arellano, I.; Rico, O.; Rubio, A.V.; Tolsá, M.J.; Roche, B.; Hosseini, P.R.; Rizzoli, A.; Murray, K.A.; et al. Metacommunity and phylogenetic structure determine wildlife and zoonotic infectious disease patterns in time and space. Ecol. Evol. 2015, 5, 865–873. [Google Scholar] [CrossRef]
- Huang, Z.Y.X.; de Boer, W.F.; van Langevelde, F.; Olson, V.; Blackburn, T.M.; Prins, H.H.T. Species’ Life-History Traits Explain Interspecific Variation in Reservoir Competence: A Possible Mechanism Underlying the Dilution Effect. PLoS ONE 2013, 8, e54341. [Google Scholar] [CrossRef]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T.; Marm Kilpatrick, A. Introduction, Spread, and Establishment of West Nile Virus in the Americas. J. Med. Entomol. 2019, 56, 1448–1455. [Google Scholar] [CrossRef]
- Ogden, N.; Tsao, J. Biodiversity and Lyme disease: Dilution or amplification? Epidemics 2009, 1, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Dobson, A.D. Pangloss revisited: A critique of the dilution effect and the biodiversity-buffers-disease paradigm. Parasitology 2012, 139, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Mihaljevic, J.R.; Joseph, M.B.; Orlofske, S.A.; Paull, S.H. The Scaling of Host Density with Richness Affects the Direction, Shape, and Detectability of Diversity-Disease Relationships. PLoS ONE 2014, 9, e97812. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Van Langevelde, F.; Estrada-Peña, A.; Suzán, G.; De Boer, W. The diversity–disease relationship: Evidence for and criticisms of the dilution effect. Parasitology 2016, 143, 1075–1086. [Google Scholar] [CrossRef]
- Norman, R.; Bowers, R.; Begon, M.; Hudson, P.J. Persistence of tick-borne virus in the presence of multiple host species: Tick reservoirs and parasite mediated competition. J. Theor. Biol. 1999, 200, 111–118. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Keesing, F. Biodiversity and Disease Risk: The Case of Lyme Disease. Conservation Biology 2000, 14, 722–728. [Google Scholar] [CrossRef]
- Estrada-Peña, A. The relationships between habitat topology, critical scales of connectivity and tick abundance Ixodes ricinus in a heterogeneous landscape in northern Spain. Ecography 2003, 26, 661–671. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Young, K.I.; Cohnstaedt, L.W.; Hanley, K.A.; Peters, D.P.C. Vector Surveillance, Host Species Richness, and Demographic Factors as West Nile Disease Risk Indicators. Viruses 2021, 13, 934. [Google Scholar] [CrossRef]
- Abellan, J.J.; Richardson, S.; Best, N. Use of Space & Time Models to Investigate the Stability of Patterns of Disease. Environ. Health Perspect. 2008, 116, 1111–1119. [Google Scholar] [CrossRef]
- Knorr-Held, L. Bayesian modelling of inseparable space-time variation in disease risk. Stat. Med. 2000, 19, 2555–2567. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Mahjoor, A.; Reiss, K.C.; Uribe, A.A.; Brown, M.T. A geostatistical model for estimating edge effects and cumulative human disturbance in wetlands and coastal waters. Int. J. Geogr. Inf. Sci. 2020, 34, 1508–1529. [Google Scholar] [CrossRef]
- Briz-Redón, Á. The impact of modelling choices on modelling outcomes: A spatio-temporal study of the association between COVID-19 spread and environmental conditions in Catalonia (Spain). Stoch. Environ. Res. Risk Assess. 2021, 35, 1701–1713. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). ArboNET. 2021. Available online: https://wwwn.cdc.gov/arbonet (accessed on 14 December 2021).
- PRISM. Total Annual Precipitation and Mean Maximum Temperature Climate Data. 2020. Available online: http://prism.oregonstate.edu (accessed on 12 June 2020).
- Sullivan, B.L.; Aycrigg, J.L.; Barry, J.H.; Bonney, R.E.; Bruns, N.; Cooper, C.B.; Damoulas, T.; Dhondt, A.A.; Dietterich, T.; Farnsworth, A.; et al. The eBird enterprise: An integrated approach to development and application of citizen science. Biol. Conserv. 2014, 169, 31–40. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Kembel, S.; Cowan, P.; Helmus, M.; Cornwell, W.; Morlon, H.; Ackerly, D.; Blomberg, S.; Webb, C. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Larsen, A.E.; Meng, K.; Kendall, B.E. Causal analysis in control–impact ecological studies with observational data. Methods Ecol. Evol. 2019, 10, 924–934. [Google Scholar] [CrossRef]
- Laubach, Z.M.; Murray, E.J.; Hoke, K.L.; Safran, R.J.; Perng, W. A biologist’s guide to model selection and causal inference. Proc. R. Soc. Biol. Sci. 2021, 288, 20202815. [Google Scholar] [CrossRef]
- Meisner, J.; Kato, A.; Lemerani, M.; Miaka, E.M.; Taban, A.I.; Wakefield, J.; Rowhani-Rahbar, A.; Pigott, D.; Mayer, J.; Rabinowitz, P. Livestock, pathogens, vectors, and their environment: A causal inference-based approach to estimating the pathway-specific effect of livestock on human African trypanosomiasis risk. medRxiv 2022, 1, 1–17. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Srygley, R.B.; Lawton, D.; Hudson, A.R.; Branson, D.H. Grasshoppers exhibit asynchrony and spatial non-stationarity in response to the El Niño/Southern and Pacific Decadal Oscillations. Ecol. Model. 2022, 471, 110043. [Google Scholar] [CrossRef]
- Bivand, R.; Piras, G. Comparing Implementations of Estimation Methods for Spatial Econometrics. J. Stat. Softw. 2015, 63, 1–36. [Google Scholar] [CrossRef]
- Simpson, D.; Rue, H.; Riebler, A.; Martins, T.G.; Sørbye, S.H. Penalising Model Component Complexity: A Principled, Practical Approach to Constructing Priors. Stat. Sci. 2017, 32, 1–28. [Google Scholar] [CrossRef]
- Riebler, A.; Sørbye, S.H.; Simpson, D.; Rue, H.; Lawson, A.B.; Lee, D.; MacNab, Y. An intuitive Bayesian spatial model for disease mapping that accounts for scaling. Stat. Methods Med. Res. 2016, 25, 1145–1165. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Douglas, D.C.; Ramey, A.M.; Mullinax, J.M.; Soos, C.; Link, P.; Walther, P.; Prosser, D.J. The spatial-temporal relationship of blue-winged teal to domestic poultry: Movement state modeling of a highly mobile avian influenza host. J. Appl. Ecol. 2021, 58, 2040–2052. [Google Scholar] [CrossRef]
- Blangiardo, M.; Cameletti, M. Spatial and Spatio-Temporal Bayesian Models with R-INLA; John Wiley & Sons, Inc.: Chichester, UK, 2015. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. Soc. Ser. Stat. Methodol. 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Martins, T.G.; Simpson, D.; Lindgren, F.; Rue, H. Bayesian computing with INLA: New features. Comput. Stat. Data Anal. 2013, 67, 68–83. [Google Scholar] [CrossRef]
- Lindgren, F.; Rue, H. Bayesian Spatial Modelling with R-INLA. J. Stat. Softw. 2015, 63, 1–25. [Google Scholar] [CrossRef]
- Verbosio, F.; De Coninck, A.; Kourounis, D.; Schenk, O. Enhancing the scalability of selected inversion factorization algorithms in genomic prediction. J. Comput. Sci. 2017, 22, 99–108. [Google Scholar] [CrossRef]
- Kourounis, D.; Fuchs, A.; Schenk, O. Towards the Next Generation of Multiperiod Optimal Power Flow Solvers. IEEE Trans. Power Syst. 2018, 33, 4005–4014. [Google Scholar] [CrossRef]
- Van Niekerk, J.; Bakka, H.; Rue, H.; Schenk, O. New frontiers in Bayesian modeling using the INLA package in R. arXiv 2019, arXiv:1907.10426. [Google Scholar] [CrossRef]
- Gelman, A.; Hwang, J.; Vehtari, A. Understanding predictive information criteria for Bayesian models. Stat. Comput. 2014, 24, 997–1016. [Google Scholar] [CrossRef]
- Gneiting, T.; Raftery, A.E. Strictly Proper Scoring Rules, Prediction, and Estimation. J. Am. Stat. Assoc. 2007, 102, 359–378. [Google Scholar] [CrossRef]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; Van Der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. Stat. Methodol. 2002, 64, 583–639. [Google Scholar] [CrossRef]
- Hooten, M.B.; Hobbs, N.T. A guide to Bayesian model selection for ecologists. Ecol. Monogr. 2015, 85, 3–28. [Google Scholar] [CrossRef]
- Dawson, W.R.; Marsh, R.L.; Yacoe, M.E. Metabolic adjustments of small passerine birds for migration and cold. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1983, 245, R755–R767. [Google Scholar] [CrossRef]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Elias, S.P.; Keenan, P.; Kenney, J.L.; Morris, S.R.; Covino, K.M.; Robinson, S.; Foss, K.A.; Rand, P.W.; Lubelczyk, C.; Lacombe, E.H.; et al. Seasonal patterns in eastern equine encephalitis virus antibody in songbirds in southern Maine. Vector-Borne Zoonotic Dis. 2017, 17, 325–330. [Google Scholar] [CrossRef]
- Edman, J.; Timperi, R.; Werner, B. Epidemiology of eastern equine encephalitis in Massachusetts. J. Fla. Mosq. Control. Assoc. 1993, 64, 84–96. [Google Scholar]
- Morris, C.D.; Zimmerman, R.H.; Edman, J.D. Epizootiology of Eastern Equine Encephalomyelitis Virus in Upstate New York, USA: II. Population dynamics and vector potential of adult Culiseta melanura (Diptera: Culicidae) in relation to distance from breeding site. J. Med. Entomol. 1980, 17, 453–465. [Google Scholar] [CrossRef]
- Morris, C.D.; Zimmerman, R.H. Epizootiology of Eastern Equine Encephalomyelitis Virus in Upstate New York, USA: III. Population dynamics and vector potential of adult Culiseta morsitans (Diptera: Culicidae). J. Med. Entomol. 1981, 18, 313–316. [Google Scholar] [CrossRef]
- Armstrong, P.M.; Andreadis, T.G.; Anderson, J.F.; Stull, J.W.; Mores, C.N. Tracking eastern equine encephalitis virus perpetuation in the northeastern United States by phylogenetic analysis. Am. J. Trop. Med. Hyg. 2008, 79, 291–296. [Google Scholar] [CrossRef][Green Version]
- Young, D.S.; Kramer, L.D.; Maffei, J.G.; Dusek, R.J.; Backenson, P.B.; Mores, C.N.; Bernard, K.A.; Ebel, G.D. Molecular epidemiology of eastern equine encephalitis virus, New York. Emerg. Infect. Dis. 2008, 14, 454. [Google Scholar] [CrossRef]
- Tan, Y.; Lam, T.T.Y.; Heberlein-Larson, L.A.; Smole, S.C.; Auguste, A.J.; Hennigan, S.; Halpin, R.A.; Fedorova, N.; Puri, V.; Stockwell, T.B.; et al. Large-Scale Complete-Genome Sequencing and Phylodynamic Analysis of Eastern Equine Encephalitis Virus Reveals Source-Sink Transmission Dynamics in the United States. J. Virol. 2018, 92, e00074-18. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interaction | Parameters | R | Description |
|---|---|---|---|
| Type I | and | Unstructured space, unstructured time | |
| Type II | and | Unstructured space, structured time | |
| Type III | and | Structured space, unstructured time | |
| Type IV | and | Structured space, structured time |
| Model | DIC | WAIC | lCPO | Effects |
|---|---|---|---|---|
| Model 1 | 491,489 | 491,489 | 2.7 | |
| Model 2 | 476,292 | 476,317 | 2.6 | |
| Model 3 | 418,939 | 418,940 | 2.3 | |
| Model 4 | 418,937 | 418,939 | 2.3 | |
| Model 5 | 418,738 | 407,029 | 2.2 | |
| Model 6 | 401,792 | 401,816 | 2.2 | |
| Model 7 | 401,787 | 401,807 | 2.2 | |
| Model 8 | 401,600 | 401,141 | 2.2 | (Type I) |
| Model 9 | 395,877 | 397,125 | 1.9 | (Type II) |
| Model 10 | 391,159 | 392,244 | 1.9 | (Type III) |
| Model 11 | 205,194 | 214,075 | 1.2 | (Type IV) |
| Model 12 | 63,062 | 59,098 | 0.9 | (Type I) |
| Model 13 | 61,498 | 59,839 | 0.8 | (Type II) |
| Model 14 | 58,181 | 50,866 | 0.9 | (Type III) |
| Model 15 | 58,074 | 50,635 | 0.8 | (Type IV) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humphreys, J.M. Amplification in Time and Dilution in Space: Partitioning Spatiotemporal Processes to Assess the Role of Avian-Host Phylodiversity in Shaping Eastern Equine Encephalitis Virus Distribution. Geographies 2022, 2, 419-434. https://doi.org/10.3390/geographies2030026
Humphreys JM. Amplification in Time and Dilution in Space: Partitioning Spatiotemporal Processes to Assess the Role of Avian-Host Phylodiversity in Shaping Eastern Equine Encephalitis Virus Distribution. Geographies. 2022; 2(3):419-434. https://doi.org/10.3390/geographies2030026
Chicago/Turabian StyleHumphreys, John M. 2022. "Amplification in Time and Dilution in Space: Partitioning Spatiotemporal Processes to Assess the Role of Avian-Host Phylodiversity in Shaping Eastern Equine Encephalitis Virus Distribution" Geographies 2, no. 3: 419-434. https://doi.org/10.3390/geographies2030026
APA StyleHumphreys, J. M. (2022). Amplification in Time and Dilution in Space: Partitioning Spatiotemporal Processes to Assess the Role of Avian-Host Phylodiversity in Shaping Eastern Equine Encephalitis Virus Distribution. Geographies, 2(3), 419-434. https://doi.org/10.3390/geographies2030026