
Kiluluma ceratotherii (Nematoda: Strongylida) in a White Rhinoceros (Ceratotherium simum) from the United States: Case Report

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
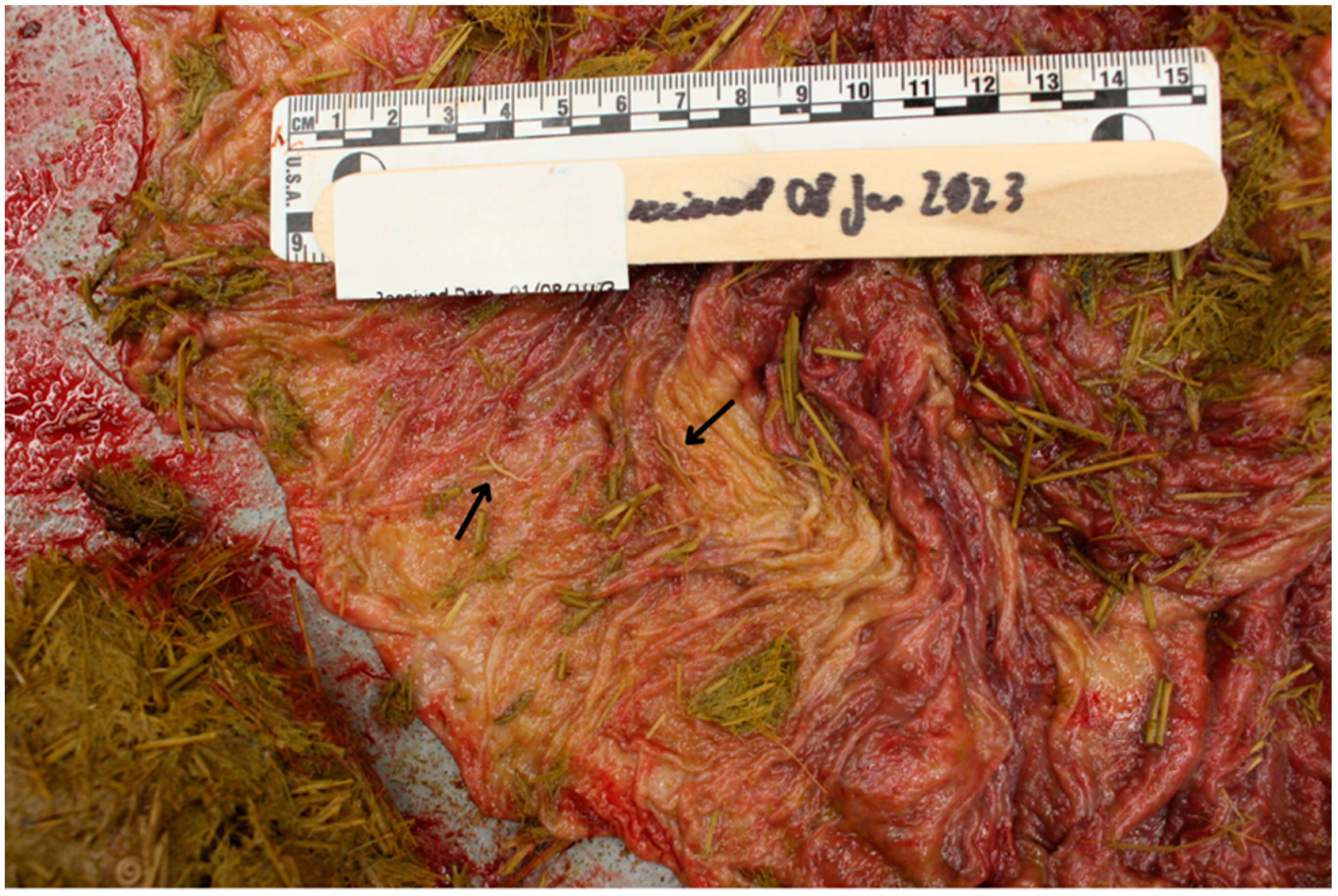
2.1. Case Presentation
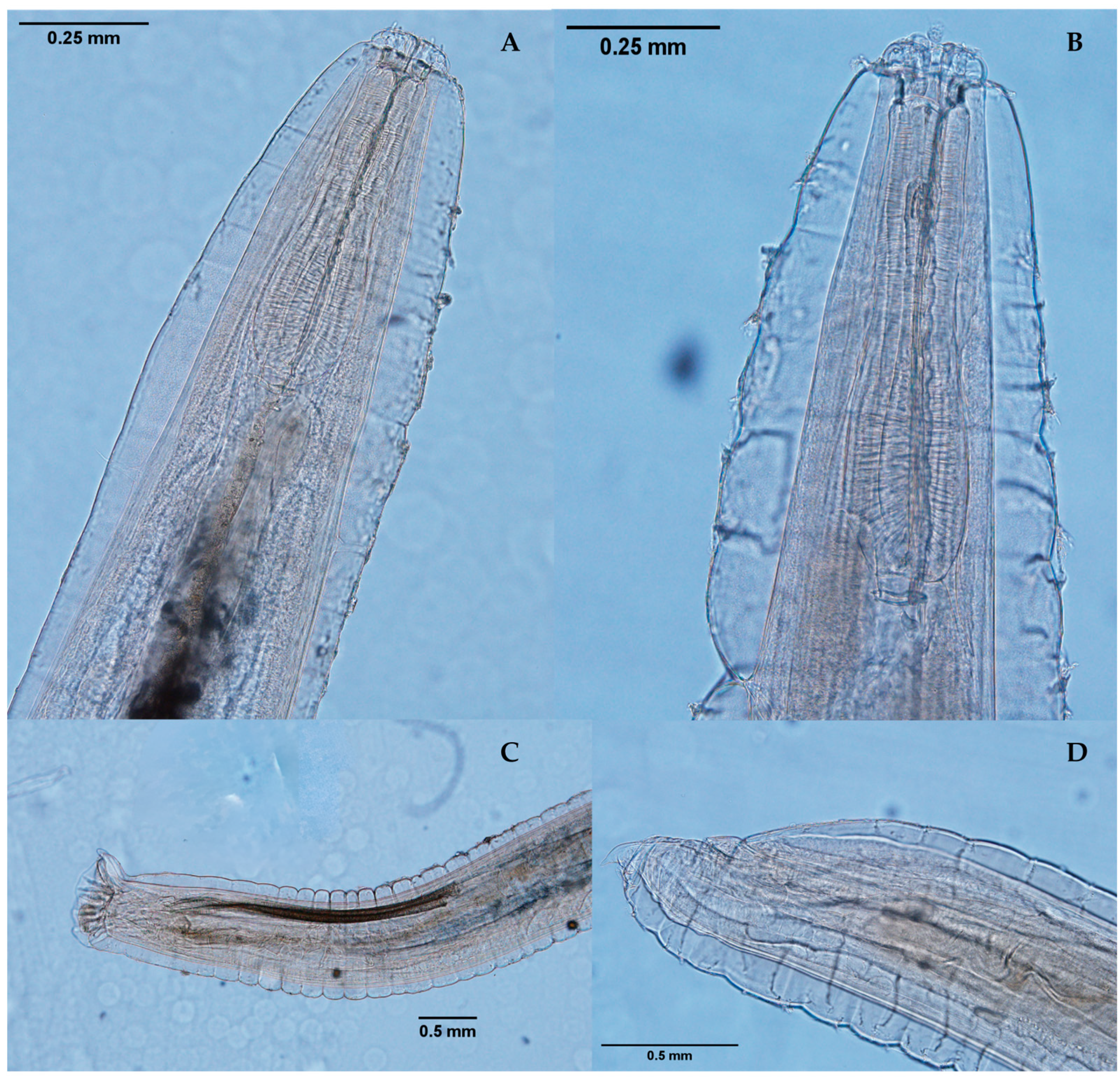
2.2. Morphological Diagnosis
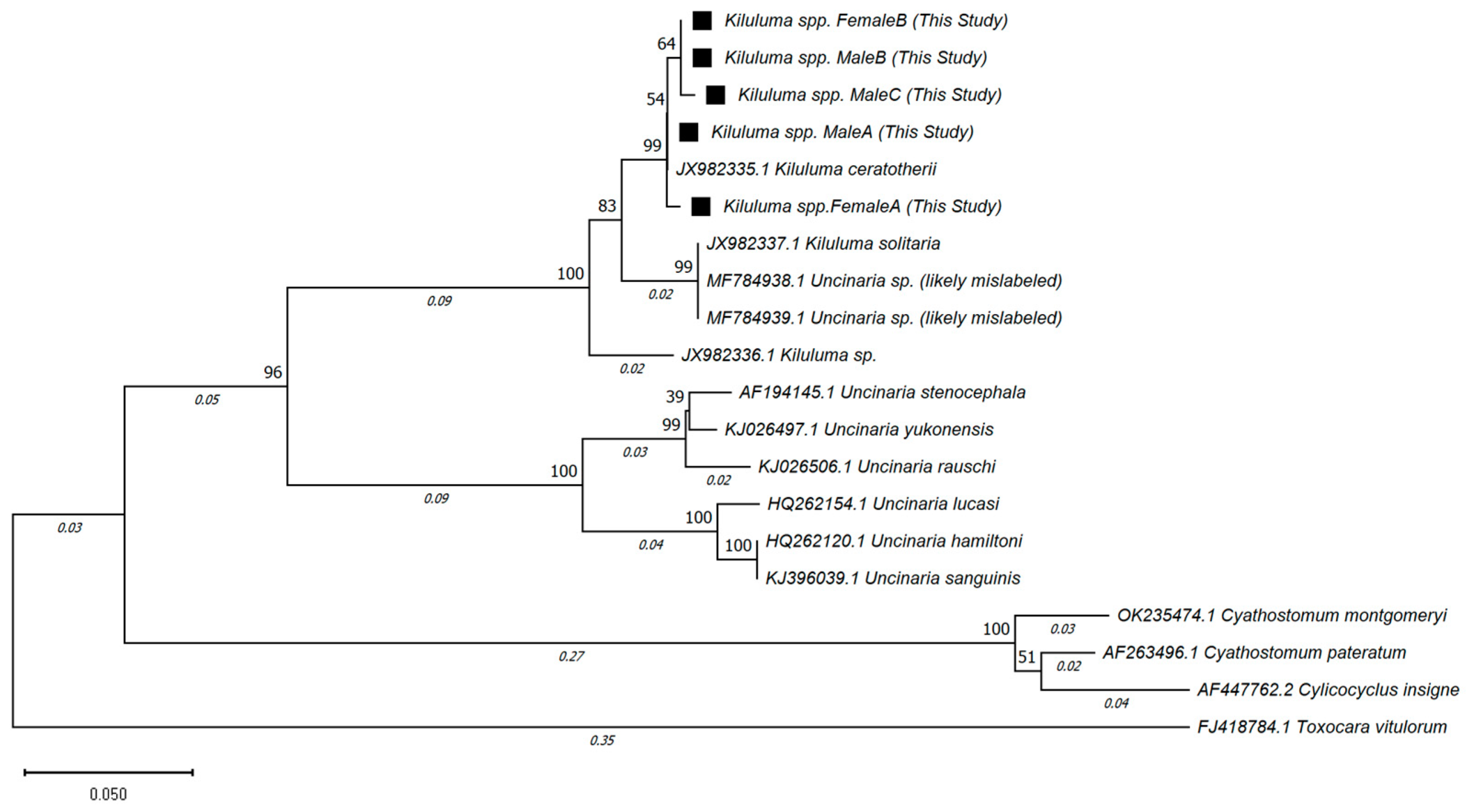
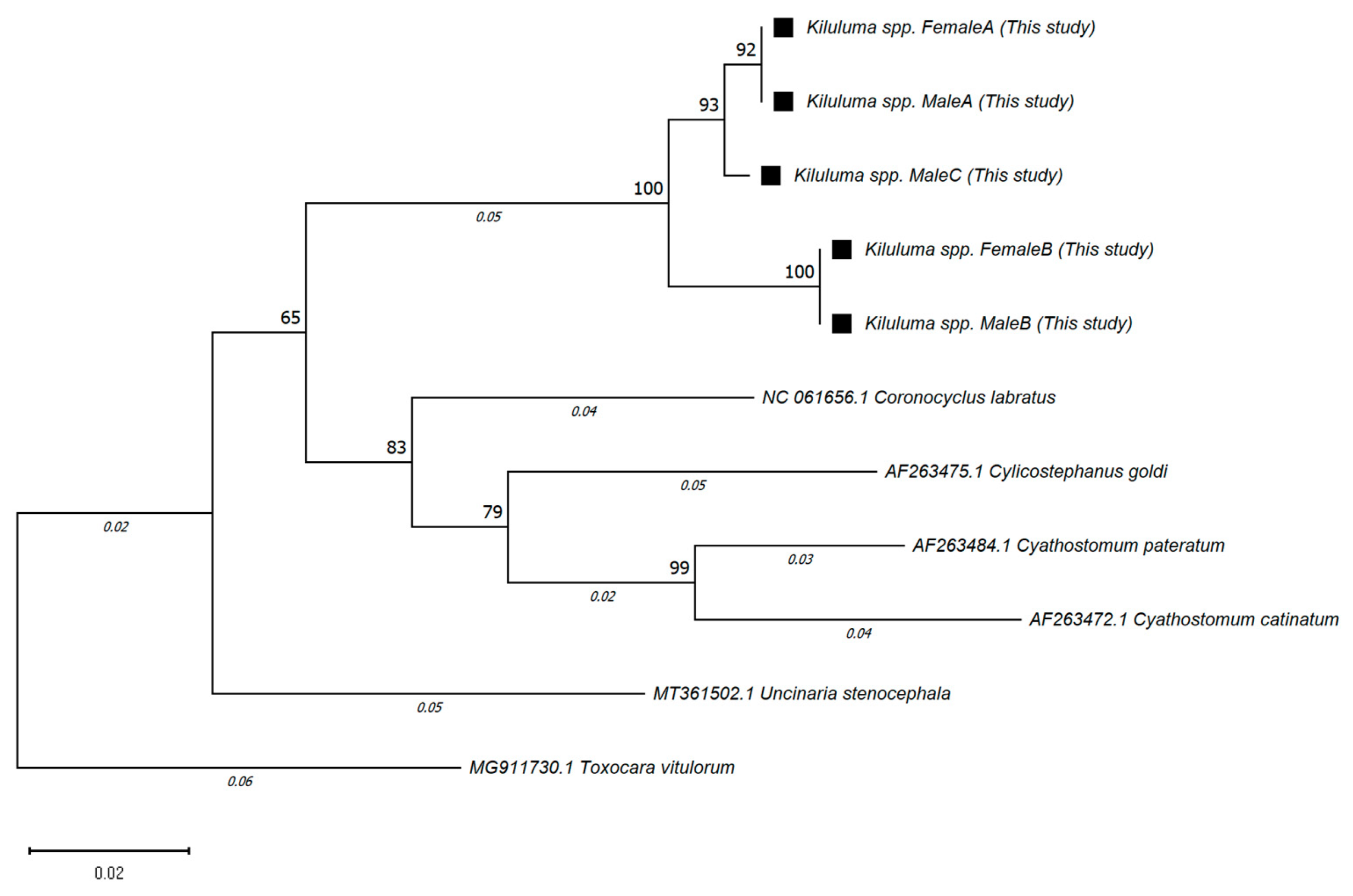
2.3. Results of Molecular Analysis
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Titcomb, G.C.; Pansu, J.; Hutchinson, M.C.; Tombak, K.J.; Hansen, C.B.; Baker, C.C.; Kartzinel, T.R.; Young, H.S.; Pringle, R.M. Large-herbivore nemabiomes: Patterns of parasite diversity and sharing. Proc. R. Soc. B 2022, 289, 20212702. [Google Scholar] [CrossRef] [PubMed]
- Knapp, S.E.; Krecek, R.C.; Horak, I.G.; Penzhorn, B.L. Helminths and arthropods of black and white rhinoceroses in southern Africa. J. Wildl. Dis. 1997, 33, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Zumpt, F. Parasites of the white and black rhinoceroses. Lammergeyer 1964, 3, 59–70. [Google Scholar]
- Beveridge, I.; Jabbar, A. New species of Kiluluma Skrjabin, 1916 (Nematoda: Strongylida) from the white rhinoceros Ceratotherium simium (Burchell), with a redescription of K. solitaria Thapar, 1924. Syst. Parasitol. 2013, 85, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Skrjabin, K. Parasitic trematodes and nematodes collected by the expedition of Prof. V. Dogiel and I. Sokolov in British East Africa. In Scientific Results of the Zoological Expedition to British East Africa and Uganda Made by Prof. V. Dogiel and I. Sokolov in the Year 1914; Forgotten Books: London, UK, 1916; Volume 1, pp. 99–157. [Google Scholar]
- Thapar, G.S. On Kiluluma Skriabin, a genus of strongylid nematodes parasitic in the African rhinoceros. J. Helminthol. 1924, 2, 209–238. [Google Scholar] [CrossRef]
- Thapar, G.S. On some new members of the genus Kiluluma from the African rhinoceros. J. Helminthol. 1925, 3, 63–80. [Google Scholar] [CrossRef]
- Taylor, E. The genus Kiluluma. Ann. Trop. Med. Parasitol. 1925, 19, 53–55. [Google Scholar] [CrossRef]
- Beveridge, I. A new species of Kiluluma Skrjabin, 1916 (Nematoda: Strongyloidea) from the white rhinoceros, Ceratotherium simum (Burchell) from South Africa. Syst. Parasitol. 2018, 95, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Chelladurai, J.J.; Bader, C.; Snobl, T.; Magstadt, D.; Cooper, V.; Brewer, M.T. Toxocara vitulorum infection in a cohort of beef calves in Iowa. Vet. Parasitol. 2015, 214, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Quintana, T.A.; Johnson, W.L.; Ritchie, D.; Smith, V.; Martin, K.A.; McMahan, K.; Brewer, M.T.; Jesudoss Chelladurai, J.R.J. Genetic characterization of the zoonotic parasite Ancylostoma caninum in the central and eastern United States. J. Helminthol. 2023, 97, e37. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintana, T.A.; Miller, K.R.; Hill-Thimmesch, K.J.; Schieferecke, G.; Johnson, W.L.; Herrin, B.H.; Jesudoss Chelladurai, J.R.J. Kiluluma ceratotherii (Nematoda: Strongylida) in a White Rhinoceros (Ceratotherium simum) from the United States: Case Report. Parasitologia 2023, 3, 241-247. https://doi.org/10.3390/parasitologia3030025
Quintana TA, Miller KR, Hill-Thimmesch KJ, Schieferecke G, Johnson WL, Herrin BH, Jesudoss Chelladurai JRJ. Kiluluma ceratotherii (Nematoda: Strongylida) in a White Rhinoceros (Ceratotherium simum) from the United States: Case Report. Parasitologia. 2023; 3(3):241-247. https://doi.org/10.3390/parasitologia3030025
Chicago/Turabian StyleQuintana, Theresa A., Kamilyah R. Miller, Kristen J. Hill-Thimmesch, Grace Schieferecke, William L. Johnson, Brian H. Herrin, and Jeba R. J. Jesudoss Chelladurai. 2023. "Kiluluma ceratotherii (Nematoda: Strongylida) in a White Rhinoceros (Ceratotherium simum) from the United States: Case Report" Parasitologia 3, no. 3: 241-247. https://doi.org/10.3390/parasitologia3030025
APA StyleQuintana, T. A., Miller, K. R., Hill-Thimmesch, K. J., Schieferecke, G., Johnson, W. L., Herrin, B. H., & Jesudoss Chelladurai, J. R. J. (2023). Kiluluma ceratotherii (Nematoda: Strongylida) in a White Rhinoceros (Ceratotherium simum) from the United States: Case Report. Parasitologia, 3(3), 241-247. https://doi.org/10.3390/parasitologia3030025