
Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe


Abstract
:1. Introduction
2. Results
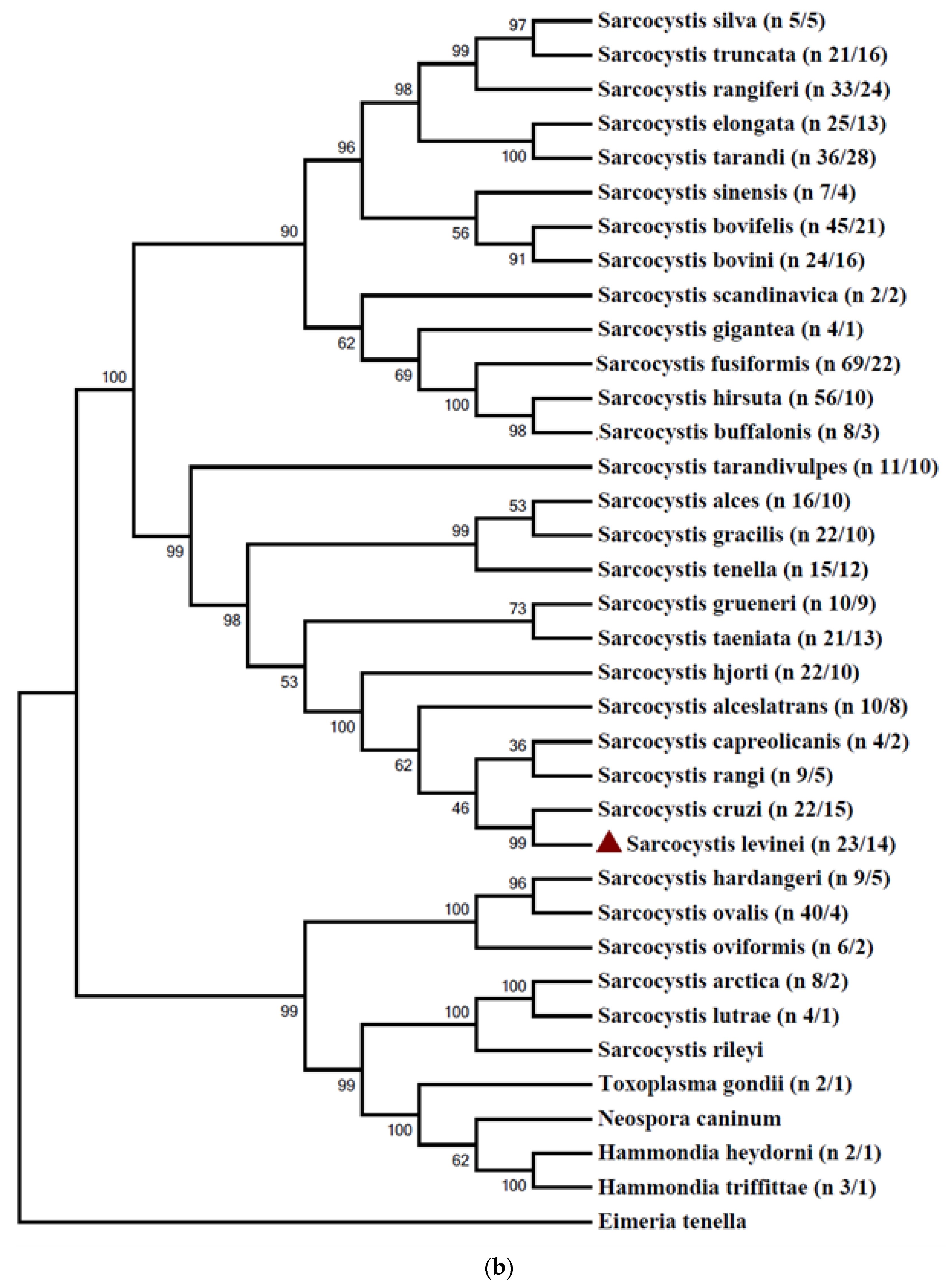
2.1. Phylogenetic Analysis of 18S rRNA and Cox 1 Gene Sequences of Various Sarcocystis spp.
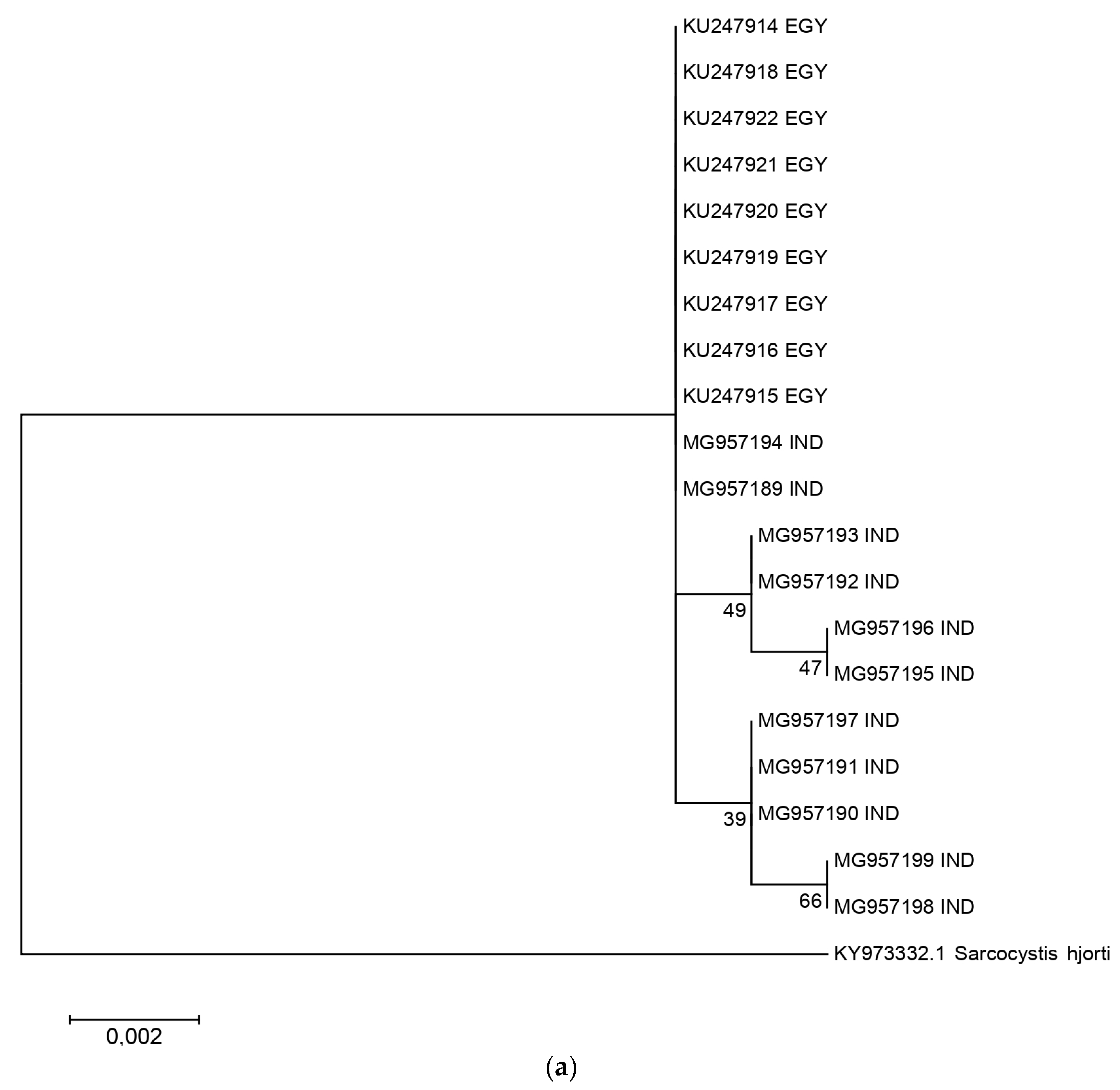
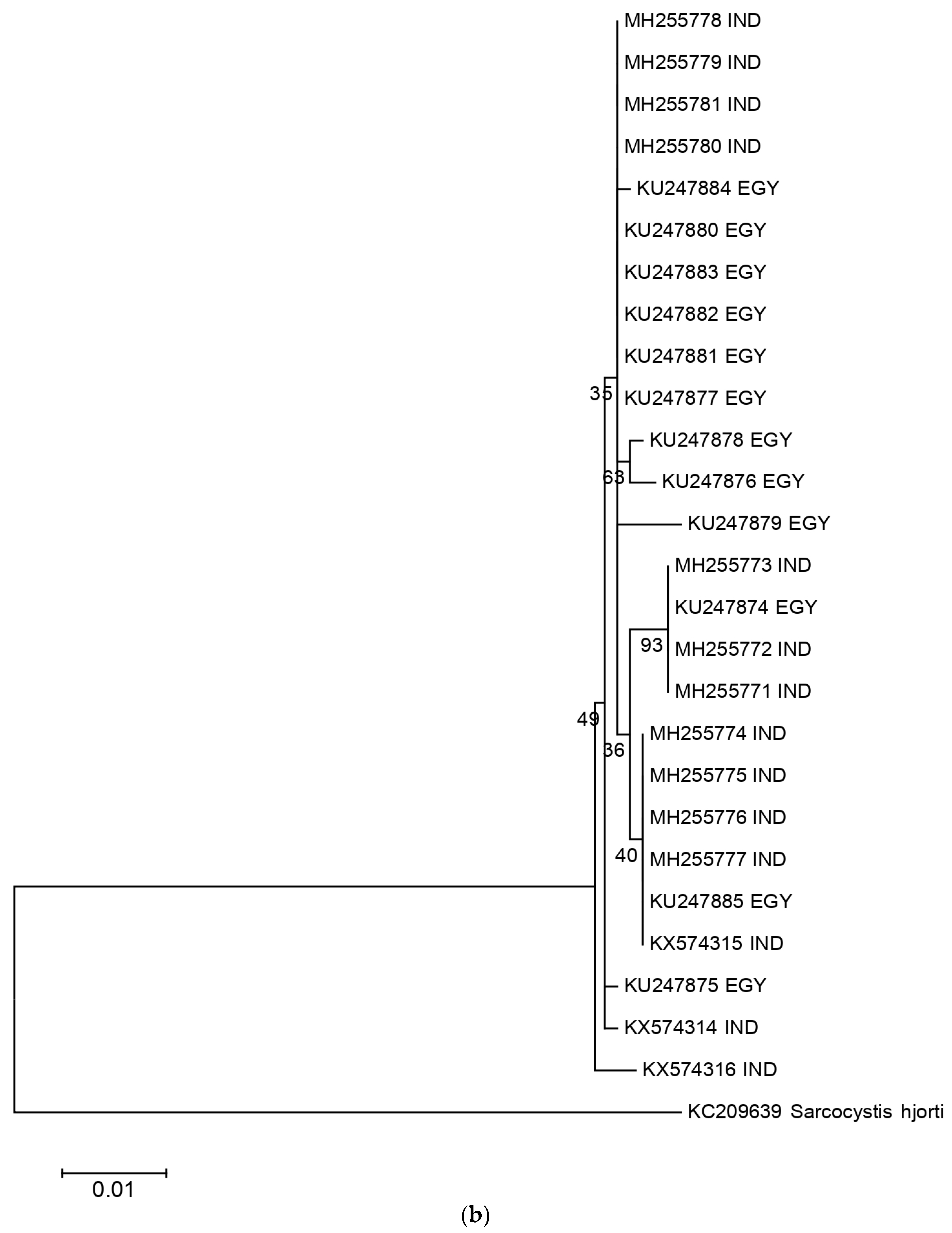
2.2. Phylogenetic Analysis and Nucleotide Identity of 18S rRNA and Cox 1 Gene Sequences of the Present Isolates with Other Isolates of S. levinei
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. DNA Isolation, Primer Selection, and Cloning of 18S rRNA and Cox 1 Genes of S. levinei
4.3. Sequence Analysis and Construction of Phylogenetic Trees
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 18S rRNA | 18S ribosomal RNA |
| 28S rRNA | 28S ribosomal RNA |
| ITS | Internal Transcribed Spacer |
| cox 1 | mitochondrial cytochrome 1 |
| DNA | Deoxyrobose Nucleic Acid |
| PCR | Polymerase Chain Reaction |
| NCBI | National Center for Biotechnology Information |
| MEGA | Molecular Evolutionary Genetics Analysis |
References
- Chen, X.; Zuo, Y.; Rosenthal, B.M.; He, Y.; Cui, L.; Yang, Z. Sarcocystis sinensis is an ultrastructurally distinct parasite of water buffalo that can cause foodborne illness but cannot complete its life-cycle in human beings. Vet. Parasitol. 2011, 178, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 243–248. [Google Scholar]
- Ashmawy, K.L.; Somaia, S.; Akkada, A.; Gashir, M.B. Prevalence and molecular characterization of Sarcocystis spp. in water buffaloes (Bubalis bubalis) in Egypt. Trop. Anim. Health Prod. 2014, 46, 1351–1356. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Sharifiyazdi, H.; Khordadmehr, M.; Larki, S. Characterization of Sarcocystis fusiformis based on sequencing and PCR-RFLP in water buffalo (Bubalus bubalis) in Iran. Parasitol. Res. 2011, 109, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B.; Hilali, M.; Abbas, I.E. Molecular differentiation of Sarcocystis buffalonis and Sarcocystis levinei in water buffaloes (Bubalus bubalis) from Sarcocystis hirsuta and Sarcocystis cruzi in cattle (Bos taurus). Parasitol. Res. 2016, 115, 2459–2471. [Google Scholar] [CrossRef] [PubMed]
- Sudan, V.; Kumar, R.; Shanker, D.; Paliwal, S. First report of molecular characterization and phylogenetic analysis of Sarcocystis tenella from India. Parasitol. Res. 2019, 118, 1429–1434. [Google Scholar] [CrossRef]
- Sudan, V.; Shanker, D.; Paliwal, S.; Kumar, R.; Singh, A. Phylogenetics of Sarcocystis fusiformis isolates based on 18S rRNA and cox 1 genes. Microb. Pathog. 2021, 159, 105144. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- Jehle, C.; Dinkel, A.; Sander, A.; Morent, M.; Romig, T.; Luc, P.; De, T.; Thai, V.; Mackenstedt, U. Diagnosis of Sarcocystis spp. in cattle (Bos taurus) and water buffalo (Bubalus bubalis) in Northern Vietnam. Vet. Parasitol. 2009, 166, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Z.; He, Y.; Zhao, H.; Rosenthal, B.M.; Dunams, D.B.; Li, X.; Zuo, Y.; Feng, G.; Cui, L.; Yang, Z. Sarcocystis cruzi: Comparative studies confirm natural infections of buffaloes. Exp. Parasitol. 2011, 127, 460–466. [Google Scholar] [CrossRef]
- Sudan, V.; Shanker, D. Redescription of Sarcocystis Species Affecting Buffaloes in the Wake of Advancement in Molecular Biology in Parasitology: Camera Lucida to OMICS; Himanshu Publishers: Udaipur, India, 2018; pp. 133–141. [Google Scholar]
- Sudan, V.; Shanker, D.; Kumar, R.; Sachan, A. Pathological studies on bubaline tissues naturally infected with Sarcocystis spp. J. Vet. Parasitol. 2019, 33, 8–11. [Google Scholar] [CrossRef]
- Ghoshal, S.B.; Joshi, S.C.; Shah, H.L. A note on the natural occurrence of Sarcocystis in buffaloes (Bubalus bubalis) in Jabalpur region, M.P. Indian Vet. J. 1986, 63, 165–166. [Google Scholar]
- Chhabra, M.B.; Samantaray, S. Sarcocystis and sarcocystosis in India: Status and emerging perspectives. J. Parasit. Dis. 2012, 37, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudan, V.; Kumar, R.; Sachan, D. Molecular identification of Sarcocystis cruzi and S. hirsuta sarcocysts in Mathura, Uttar Pradesh. J. Vet. Parasitol. 2020, 32, 8–11. [Google Scholar]
- Huong, L.T.T.; Dubey, J.P.; Uggla, A. Redescription of Sarcocystis levinei Dissanaike and Kan, 1978 (Protozoa: Sarcocystidae) of the water buffalo (Bubalus bubalis). J. Parasitol. 1997, 83, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B.; Hilali, M.; Mawgood, S.A. Molecular characterization of three regions of the nuclear ribosomal DNA unit and the mitochondrial cox 1 gene of Sarcocystis fusiformis from water buffaloes (Bubalus bubalis) in Egypt. Parasitol. Res. 2015, 114, 3401–3413. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Q.; Zuo, Y.X.; Ding, B.; Chen, X.W.; Luo, J.; Zhang, Y.P. Identification of Sarcocystis hominis-like (Protozoa:Sarcocystidae) cyst in water buffalo (Bubalus bubalis) based on 18S rRNA gene sequences. J. Parasitol. 2001, 87, 934–937. [Google Scholar] [CrossRef]
- Gjerde, B. Molecular characterisation of Sarcocystis bovifelis, Sarcocystis bovini n. sp., Sarcocystis hirsuta and Sarcocystis cruzi from cattle (Bos taurus) and Sarcocystis sinensis from water buffaloes (Bubalus bubalis). Parasitol. Res. 2015, 115, 1473–1492. [Google Scholar] [CrossRef]
- Sudan, V.; Kumar, R.; Shanker, D.; Singh, A. Sequence phylogenetic analysis and associative genetic diversity of Sarcocystis hirsuta based on 18S rRNA gene. Beni Suef Univ. J. Basic Appl. Sci. 2021, 10, 22. [Google Scholar] [CrossRef]
- Rosenthal, B.M.; Dunams, D.B.; Pritt, B. Restricted genetic diversity in the ubiquitous cattle parasite, Sarcocystis cruzi. Infect. Genet. Evol. 2008, 8, 588–592. [Google Scholar] [CrossRef]
- Bhoora, R.; Franssen, L.; Oosthuizen, M.C.; Guthrie, A.J.; Zweygarth, E.; Penzhorn, B.L.; Jongejan, F.; Collins, N.E. Sequence heterogeneity in the 18S rRNA gene within Theileria equi and Babesia caballi from horses in South Africa. Vet. Parasitol. 2009, 159, 112–120. [Google Scholar] [CrossRef] [Green Version]
- El-Sherry, S.; Ogedengbe, M.E.; Hafeez, M.A.; Barta, J.R. Divergent nuclear 18S rDNA paralogs in a turkey coccidium, Eimeria meleagrimitis, complicate molecular systematics and identification. Int. J. Parasitol. 2013, 43, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Vrba, V.; Poplstein, M.; Pakandl, M. The discovery of the two types of small subunit ribosomal RNA gene in Eimeria mitis contests the existence of E. mivati as an independent species. Vet. Parasitol. 2011, 183, 47–53. [Google Scholar] [CrossRef] [PubMed]
- El-Seify, M.; El-Morsey, A.; Hilali, M.A.; Zayed, A.; El-Dakhly, K.; Haridy, M.; Sakai, H.; Yanai, T.; Seify, E. Molecular characterization of Sarcocytis fusiformis and Sarcocystis buffalonis infecting water buffaloes (Bubalus bubalis) from Egypt. Am. J. Anim. Vet. Sci. 2014, 9, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Holmdahl, O.J.; Mattsson, J.G.; Uggla, A.; Johansson, K.E. The phylogeny of Neospora caninum and Toxoplasma gondii based on ribosomal RNA sequences. FEMS Microbiol. Lett. 1994, 119, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Nh | Hd | Pi | S | K |
|---|---|---|---|---|---|
| Cox 1 | 10 | 0.825 | 0.00336 | 19 | 2.76308 |
| 18S | 5 | 0.679 | 0.00118 | 3 | 1.00526 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudan, V.; Shanker, D.; Paliwal, S.; Kumar, R. Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe. Parasitologia 2023, 3, 231-240. https://doi.org/10.3390/parasitologia3030024
Sudan V, Shanker D, Paliwal S, Kumar R. Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe. Parasitologia. 2023; 3(3):231-240. https://doi.org/10.3390/parasitologia3030024
Chicago/Turabian StyleSudan, Vikrant, Daya Shanker, Sanjhi Paliwal, and Rahul Kumar. 2023. "Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe" Parasitologia 3, no. 3: 231-240. https://doi.org/10.3390/parasitologia3030024
APA StyleSudan, V., Shanker, D., Paliwal, S., & Kumar, R. (2023). Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe. Parasitologia, 3(3), 231-240. https://doi.org/10.3390/parasitologia3030024