Abstract
Selaginella iridescens (Selaginellaceae), a new species of spikemoss from Hainan Island, China, is described based on morphological and molecular evidence. The new species morphologically resembles S. pseudotamariscina from Vietnam which was recently recognized and segregated from S. tamariscina and S. pulvinata. Some characters of S. iridescens, including the iridescent leaves and long arista (0.35–1.20 mm long), the widely membranaceous margin of axillary leaves (ca. 2/3), dorsal leaves sulcate extending to the top, and slender main stem, dorsoventrally prostrate, distinguish it from S. pseudotamariscina. Phylogenetic results based on rbcL of 28 Selaginella species indicate that S. iridescens is sister to S. pseudotamariscina, and distant from the S. tamariscina-S. pulvinata clade. The ancestral character reconstruction result reveals that the rosette is apomorphic and has evolved independently at least six times in Selaginella.
1. Introduction
Selaginella P. Beauv., the single genus in Selaginellaceae, is the largest extant lycophyte genus that comprises approximately 700–800 taxonomically difficult species [1,2,3,4]. The monophyly of Selaginella was well-supported by recent phylogenetic studies, however, the infrageneric classification is still uncertain [1,5,6,7]. Based on the morphological characters, Jermy [1] divided the genus Selaginella into five subgenera, whose classification was the most widely accepted before molecular phylogenetic studies. Recently, Weststrand and Korall [3,4] divided the genus Selaginella into seven subgenera based on DNA sequences (rbcL, pgiC, and SQD1) and morphological data. This classification system was supported by the phylogenomic study based on plastome datasets, which further resolved the uncertainty of the S. sanguinolenta group as the most basal clade of S. subg. Stachygynandrum [7].
Species of Selaginella have various growth forms, including rosette, creeping, climbing, prostrate, erect, and suberect [2,8,9]. The rosette is a special morphological feature in Selaginella, but only few species are known to be rosette-forming [4,6,10]. In China, S. tamariscina (P. Beauv.) Spring and S. pulvinata (Hook. et Grev.) Maxim are two famous rosette-forming species of Selaginella [2,9]. In Hainan Island, about 14 Selaginella species are recorded in Flora of China (FOC), including two endemic species (S. hainanensis X. C. Zhang and Nooteboom and S. scarbrifolia Ching and Chu H. Wang) and one rosette-forming species (S. tamariscina) [2,11]. During a field trip in Hainan Island of China, a new rosette-forming Selaginella species was collected. A detailed examination on morphology of our newly collected materials and potential similar species revealed that these specimens were morphologically distinct from the two famous rosette-forming species in China, S. tamariscina and S. pulvinata, but morphologically similar to S. pseudotamariscina X.C. Zhang and C.W. Chen, a new species recently described from Vietnam [12]. Then, we conducted phylogenetic analyses using rbcL sequences, including this new species and other rosette species within Selaginella. In this paper, we present evidence from morphology and molecular phylogenetic analysis to support the new species and conduct ancestral character reconstruction to reveal the evolutionary history of rosette character in Selaginella.
2. Materials and Methods
2.1. Morphological Observations
Morphology of the newly collected specimens was examined and photographed under a Leica S9D stereo microscope, and compared with S. pseudotamariscina, S. pulvinata and S. tamariscina. The sterile leaves, strobili, and sporophylls were observed and measured.
2.2. Taxon Sampling and Sequencing
We sampled three individuals of the new species from one site on Hainan Island (Figure 1). Total genomic DNA was extracted from silica gel dried materials using the Plant Genomic DNA Kit DP305 (TianGen Biotech, Beijing, China) following the manufacturer’s protocol. Libraries for pair-end 150 bp sequencing with a 350 bp insert size were conducted using an Illumina NovaSeq 6000 platform at BioMarker Co (Beijing, China). Raw reads were filtered based on the following criteria: pair-end reads with >10% ‘N’ bases; reads, on which more than 50% of the bases have a quality score less than 10 (Phred-like score). Finally, approximate 6 Gb high-quality sequences were obtained for each sample (only rbcL sequences were extracted for this study, this high-throughput data will be used for further study).
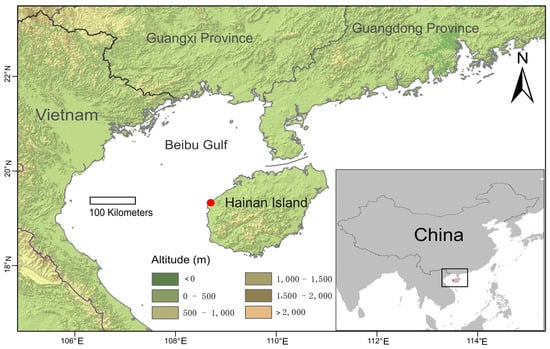
Figure 1.
Distribution map of Selaginella iridescens. The red dot represents the location of S. iridescens. The color scale indicates the elevation in the meters above sea level as recorded in the raster layers of 30 s resolution of the NASA Shuttle Radar topographic Mission (SRTM) elevation database deposited in WorldClim (https://www.worldclim.org, accessed on 24 October 2021). The basic map of China was downloaded from National Catalogue Service for Geographic Information (https://www.webmap.cn, accessed on 25 October 2021).
2.3. Phylogenetic Analysis
The chloroplast rbcL gene was the most commonly used marker in Selaginella [3,5,6,10]. Therefore, rbcL was selected to reconstruct a phylogeny with an extensive species sampling to clarify the relationships between the new Selaginella species and other rosette species. In total, 32 individuals of 28 ingroup species representing all the seven subgenera of Selaginella according to Weststrand and Korall [3,4] were included, while Isoetes histrix Bory and Durand and I. sinensis Palmer were selected as outgroups. The rbcL of new species were obtained by Geneious mapping using Illumina short reads in Geneious v. 11.1.4 [13] and rbcL sequences for other species were downloaded from GenBank (voucher information and GenBank accession numbers listed in the Table 1).

Table 1.
Species names and GenBank accession numbers of rbcL sequences used in this study.
All the sequences were aligned using MAFFTT v7.313 [14,15]. Both maximum likelihood (ML) analysis and Bayesian inference (BI) were carried out in this study. ML analysis was performed using RAxML 7.2.6 [16], with 1000 bootstrap replicates under the GTRGAMMA model selected according to the Bayesian Information Criterion (BIC) by ModelFinder [17]. BI analysis was performed using MrBayes v. 3.2.6. [18], under the SYM + G4 model selected according to the BIC by ModelFinder [17]. For each Bayesian analysis, four MCMC chains were run simultaneously for 1 million generations and sampled every 1000 generations. The average standard deviation of split frequencies (<0.01) was used to assess the convergence. ML and BI trees and the branch support values were visualized using FigTree v.1.4.2 [19].
2.4. Character Evolution Analysis
We used Mesquite v.3.61 [20] to infer the ancestral states of rosette character. The ancestral character reconstruction was performed under ML models “Mk1” [21], with the phylograms of 1000 RAxML bootstrap trees based on rbcL sequences as input phylogenies in order to consider any phylogenetic uncertainties. The results were finally summarized as percentage of changes of character states on a given branch across all 1000 trees using the “Average-frequencies-across-trees” option.
3. Results
3.1. Taxonomy Treatment
Selaginella iridescens X. C. Zhang and Y. R. Wang, sp. nov.
Type. CHINA, Hainan, Changjiang, Changhua, at foot of Mt. Daling, on quartzite rocky slope, in open grass, 108.700° E, 19.328° N, altitude 31 m, 10 January 2020, Y. R. Wang and L. X. Yuan wyr20200110 (holotype, PE!); ibid. 11 June 2021, X. C. Zhang and E. F. Huang 11,594 (paratype, PE!).
Diagnosis. Selaginella iridescens differs from S. pseudotamariscina by its iridescent leaves and apex with long arista (0.35–1.2 mm), dorsal leaves adaxially sulcate, margins of axillary leaves widely membranaceous (ca. 2/3), and main stem slender, dorsiventrally prostrate (Figure 2, Figure 3 and Figure 4; Table 2).

Figure 2.
Habitat of Selaginella iridescens X. C. Zhang and Y. R. Wang sp. nov. (A) Habitat; (B) prostrate stems; (C) semi-dry individuals; (D) fresh individuals.

Figure 3.
Morphology of Selaginella iridescens X. C. Zhang and Y. R. Wang sp. nov. (A) Individual; (B) adaxial view of branchlets; (C) abaxial view of branchlets; (D) axillary leaf; (E) ventral leaf; (F) dorsal leaf; (G) ventral sporophyll; (H) dorsal sporophyll; (I) adaxial view of strobili (arrows).

Figure 4.
Comparison of leaf morphology of Selaginella iridescens, S. pseudotamariscina, S. tamariscina, and S. pulvinata. (A1–C1): S. iridescens, Hainan, Y. R. Wang, wyr20200110 (PE); (A2–C2): S. pseudotamariscina, Vietnam, C. W. Chen, Wade 5314 (PE); (A3–C3): S. tamariscina, Hainan, H. Y. Liang 63,716 (PE); (A4–C4): S. pulvinata, Yunnan, X. C. Zhang 088 (PE). A. ventral leaves; B. dorsal leaves; C. axillary leaves. Scale bars = 0.2 mm.
Table 2.
Character comparison of S. iridescens, S. pseudotamariscina, S. pulvinata, and S. tamariscina.
Description. Rosettes, xerophytic. Rhizophores restricted to base of stem; rhizophores much forked, forming thick massive rootstock. Stems and roots entangled forming prostrate dorsiventral trunks; main stems branched near or above the base, pinnately or anisotomously branched, stramineous or brown; primary leafy 2–3 pinnately branched, branchlets compact and regular. Leaves thick and surface smooth, fresh leaves iridescent. Axillary leaves on branches symmetrical, ovate-oblong, ca. 2.02 × 0.87 mm, margin widely membranaceous (about 2/3 of the entire leaf), lacerate or subentire, base obtuse, ciliolate or denticulate, apex arista (ca. 0.52 mm long). Dorsal leaves strongly ascending, imbricate, symmetrical, lanceolate, 2.27 × 0.70 mm, lower side sulcate extends to the top, upper side carinate, base obtuse, margin ciliolate or denticulate, not membranaceous, apex arista (ca. 0.76 mm long). Ventral leaves slightly ascending, overlapping, asymmetrical, ovate-lanceolate to ovate-triangular, ca. 2.6 × 1.5 mm, adaxially sulcate extends to the top, apex arista (ca. 0.35–1.20 mm long); basiscopic margin ciliolate or denticulate; acroscopic base enlarged, broader, widely membranaceous, lacerate or subentire, ciliolate or lacerate at base. Strobili solitary, terminal, compact, slightly dorsoventrally complanate, very short (ca. 1–5 mm long); sporophylls anisophyllous; dorsal sporophylls smaller than the ventral ones, symmetrical, lanceolate, ca. 2.0 × 0.8 mm, margin slightly membranaceous, shortly ciliolate, apex arista; ventral sporophylls triangular, ca. 2.21 × 1.30 mm, margin widely membranaceous, denticulate, ciliolate or lacerate, apex arista; sporangia borne only on the ventral sporophylls; microspores yellowish orange, megaspores pale yellow to white (Figure 2 and Figure 3).
Distribution and habitat. Selaginella iridescens is only known from Western Hainan, Changjiang County, on quartzite rocky slope, open area (Figure 1).
Etymology. The specific epithet ‘iridescens’ refers to its iridescent leaves.
Conservation status. Selaginella iridescens is known only from one population with about 300 individuals in the type locality. No other localities were discovered during our investigation conducted both in 2020 and 2021. Moreover, detailed examination of relevant specimens collected from Hainan and adjacent regions of rosette Selaginella species in virtual herbaria (CVH and GBIF) revealed no additional specimens of this species. Even if the assignment of a conservation status of this new species could be premature, it is most likely an endemic to Hainan Island and could be temporarily considered as Vulnerable (VU) according to the IUCN Red List guidelines criterion D1D2 based on current data [22].
3.2. Phylogenetic Analysis
The total length of rbcL alignment is 1428 bp, with 475 parsimony informative sites. The sequences of three Sealginella iridescens individuals are identical and have 14 nucleotide variations when compared with S. pseudotamariscina, the most related species to S. iridescens.
Our phylogenetic analyses of 28 species of Selaginella based on rbcL gene are generally similar to the former phylogeny studies and consistent with the seven subgenera classification [3,4,5]. Three individuals of S. iridescens are clustered together and sister to S. pseudotamariscina, and formed a monophyletic clade belonging to the Subg. Stachygynandrum (Figure 5). This clade is sister to the S. helicoclada-S. imbricata clade with strong support (BS = 100/PP = 100). The clade grouping of these two latter clades (S. iridescens-S. pseudotamariscina clade and S. helicoclada-S. imbricata clade) is sister to the S. pulvinata-S. stauntoniana clade (BS = 99/PP = 100).
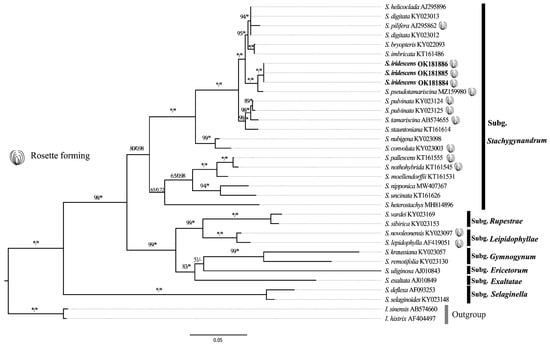
Figure 5.
Maximum likelihood tree based on the rbcL gene. Maximum likelihood bootstrap (BS) and Bayesian inference posterior probability (PP) are noted above the branch (PP/BS, * = 1 or 100%).
3.3. Ancestral Reconstruction of Rosette Character
Ancestral character reconstruction results reveal that rosette is apomorphic in Selaginella. This character has experienced a complicated evolutionary history and has evolved independently at least six times in Selaginella. There were five times in subg. Stachygynandrum and once in subg. Lepidophyllae. The five times in subg. Stachygynandrum appeared in S. nothohybrida-S. pallescens clade, S. convoluta-S. nubigena clade, S. pulvinata-S. tamariscina clade, S. irisdescens-S. pseudotamariscina clade, and S. pilifera clade, respectively (Figure 6).
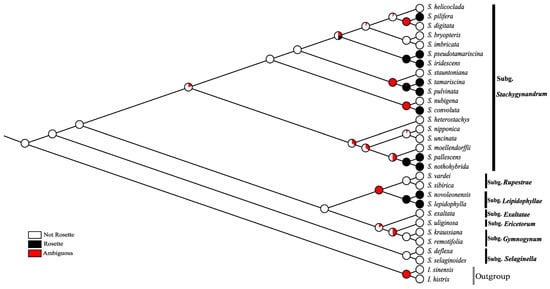
Figure 6.
The inferred ancestral states for rosette habit based on the Mk1 model. Black and white in the pie charts on each branch indicate the probabilities of ancestral states inferred by the analysis based on the 1000 RAxML bootstrap trees.
4. Discussion
4.1. Morphological Comparison with Three Related Rosette Species
Morphological comparison with S. pulvinata and S. tamariscina shows that S. iridescens is distinguished by its fresh leaves iridescent; dorsal leaves symmetrical, lanceate, adaxially sulcate, margin ciliolate or denticulate; ventral leaves adaxially shallow sulcate; strobili dorsoventrally complanate, similar shape of dorsal sporophylls and dorsal sterile leaves; dorsal sporophylls smaller than the ventral ones, and sporangia borne only on the ventral sporophylls (Figure 3 and Table 2). The leaves of S. pulvinata and S. tamariscina are not iridescent, dorsal leaves asymmetrical, ventral leaves adaxially not sulcate (Figure 4). The strobili of S. pulvinata and S. tamariscina are tetragonal, sporophylls isophyllous, margin denticulate, sporangia borne on both dorsal and ventral sporophylls (Table 2).
Morphological comparison with S. pseudotamariscina shows that S. iridescens is similar to S. pseudotamariscina by its rosette-forming habit and lanceate symmetrical dorsal leaves. However, S. iridescens is further characterized by the following unique features: the leaves iridescent with long arista apex (0.35–1.20 mm), dorsal leaves sulcate extending to the top, margins of axillary leaves widely membranaceous (ca. 2/3), and main stem slender, dorsiventrally prostrate (Figure 2, Figure 3 and Figure 4 and Table 2). The leaves of S. pseudotamariscina are not iridescent and the apex is short arista (0.02–0.40 mm), dorsal leaves sulcate to the middle of leaves, margins of axillary leaves widely membranaceous (ca. 1/2) (Figure 4 and Table 2). The main stem of S. pseudotamariscina is thick, radial, and erect. It is obvious that these two similar species are very distinct in detail.
4.2. How Many Rosette Species in Selaginella?
Rosette is a special morphological feature, which has evolved independently at least six times in Selaginella (Figure 6). However, there are only about 12 Selaginella species known to be rosette-forming according to our new finding and former studies [3,6,12]. These 12 rosette species only disperse in two subgenera of Selaginella, S. subg. Lepidophyllae, and subg. Stachygynandrum. Subg. Lepidophyllae contains four rosette species mainly distributed in North America, but only two species (S. lepidophylla and S. novoleonensis) have been sequenced and obtained a confirmed position [3,4]. There are still two rosette-forming resurrection species, S. gypsophila A. R. Sm. and T. Reeves and S. ribae Valdespino with unknown systematic position, that may still be members of subg. Lepidophyllae [4,23,24]. There are eight rosette-forming species in subg. Stachygynandrum, mainly distributed in America (S. nothohybrida, S. pallescens, S. convoluta, and S. pilifera) and East Asia (S. iridescens, S. pseudotamariscina, S. pulvinata, and S. tamariscina) [3,10,12]. These eight rosette-forming species are not monophyletic, they fell into four small groups and occupied different positions both in present and previous phylogenetic analyses [3,10,12].
4.3. Key to S. iridescens, S. pseudotamariscina, S. pulvinata, and S. tamariscina
1. Strobili dorsoventrally complanate; sporangia borne only on the ventral sporophylls; dorsal leaves symmetrical, lanceolate, adaxially sulcate; ventral leaves abaxially shallowly sulcate………………….…………………………..………………………………… 2.
2. Main stem slender, dorsiventrally prostrate; leaves iridescent, apex long arista (0.35–1.20 mm long); margins of axillary leaves widely membranaceous (ca. 2/3); dorsal leaves adaxially sulcate to the apex………………………….……….……….…. S. iridescens.
2. Main stem thick, radially erect; leaves not iridescent, apex shortly arista (0.02–0.40 mm long); margins of axillary leaves membranaceous (ca. 1/2); dorsal leaves adaxially sulcate not to the apex ………………………………………………….. S. pseudotamariscina.
1. Strobili tetragonal; sporangia borne on both ventral and dorsal sporophylls; dorsal leaves asymmetrical, obliquely ovate, triangular, or elliptic, not sulcate………………………………..………………..……………………………………………. 3.
3. Stems and roots entangled not forming treelike trunk; inner margins of dorsal leaves lacerate or entire, outer margins revolute and entire………………..… S. pulvinata.
3. Stems and roots entangled often forming treelike trunk; inner margins of dorsal leaves denticulate, outer margins denticulate or shortly ciliolate…………… S. tamariscina.
Author Contributions
Conceptualization, X.-C.Z.; formal analysis, J.Y. and M.-H.Z.; investigation, Y.-R.W. and L.-X.Y.; writing—original draft preparation, J.Y. and M.-H.Z.; writing—review and editing, X.-C.Z.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Beijing Natural Science Foundation, grant number 5202019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Er-Feng Huang for his help during the fieldwork and providing habitat photos of the new species.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jermy, A.C. Subgeneric names in Selaginella. Fern Gaz. 1986, 13, 117–118. [Google Scholar]
- Zhang, X.C.; Nooteboom, H.P.; Kato, M. Selaginellaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2013; Volumes 2–3, pp. 37–66. [Google Scholar]
- Weststrand, S.; Korall, P. Phylogeny of Selaginellaceae: There is value in morphology after all! Am. J. Bot. 2016, 103, 2136–2159. [Google Scholar] [CrossRef]
- Weststrand, S.; Korall, P. A subgeneric classification of Selaginella (Selaginellaceae). Am. J. Bot. 2016, 103, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Korall, P.; Kenrick, P. Phylogenetic relationships in Selaginellaceae based on rbcL sequences. Am. J. Bot. 2002, 89, 506–517. [Google Scholar] [CrossRef]
- Zhou, X.M.; Zhang, L.B. A classification of Selaginella (Selaginellaceae) based on molecular (chloroplast and nuclear), macromorphological, and spore features. Taxon 2015, 64, 1117–1140. [Google Scholar] [CrossRef]
- Zhang, H.R.; Wei, R.; Xiang, Q.P.; Zhang, X.C. Plastome-based phylogenomics resolves the placement of the sanguinolenta group in the spikemoss of lycophyte (Selaginellaceae). Mol. Phylogenet. Evol. 2020, 147, 106788. [Google Scholar] [CrossRef] [PubMed]
- Jermy, A.C. Selaginellaceae. In The Families and Genera of Vascular Plants, Pteridophytes and Gymnosperms; Kramer, K.U., Green, P.S., Eds.; Springer: Berlin, Germany, 1990; Volume 1, pp. 39–45. [Google Scholar]
- Zhang, X.C. Selaginellaceae. In Flora Reipublica Popularis Sinicae; Zhang, X.C., Ed.; Science Press: Beijing, China, 2004; Volume 6, pp. 86–219. [Google Scholar]
- Zhou, X.M.; Rothfels, C.J.; Zhang, L.; He, Z.R.; Le Péchon, T.; He, H.; Lu, N.T.; Knapp, R.; Lorence, D.; He, X.J.; et al. A large-scale phylogeny of the lycophyte genus Selaginella (Selaginellaceae: Lycopodiopsida) based on plastid and nuclear loci. Cladistics 2015, 32, 360–389. [Google Scholar] [CrossRef]
- Zhang, X.C.; Xiang, Q.P.; Nooteboom, H.P. A new species of Selaginella from Hainan Island, China, with a key to the Hainan species. Bot. J. Linnean Soc. 2005, 148, 323–327. [Google Scholar] [CrossRef][Green Version]
- Zhang, M.H.; Yang, J.; Luu, H.T.; Tran, G.; Zhang, X. Selaginella pseudotamariscina (Selaginellaceae), an overlooked rosette-forming resurrection spikemoss from Vietnam. Guihaia 2021. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 491–587. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree, Version 1.4.2. Computer Program and Documentation Distributed by the Author. 2014. Available online: https://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 September 2021).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. 2019. Available online: https://www.mesquiteproject.org (accessed on 14 September 2021).
- Lewis, P.O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef] [PubMed]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria, Version 14. Prepared by the Standards and Petitions Committee. 2019. Available online: https://www.iucnredlist.org/documents/RedListGuidelines.pdf (accessed on 24 December 2020).
- Smith, A.R.; Reeves, T. Selaginella gypsophila (Selaginellaceae), yet another new edaphic endemic from northern Mexico. SIDA Contrib. Bot. 1984, 10, 211–215. [Google Scholar]
- Mickel, J.T.; Smith, A.R.; Valdespino, I.A. Selaginella. In The Pteridophytes of Mexico; Mickel, J.T., Smith, A.R., Eds.; Parts I and II Memoirs of the New York Botanical Garden; New York Botanical Garden Press: New York, NY, USA, 2004; pp. 88, 550–602, 968–989. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).