

Overproduction of Phenolic Compounds in Pseudomonas putida KT2440 Through Endogen Deregulation of the Shikimate Pathway


Abstract
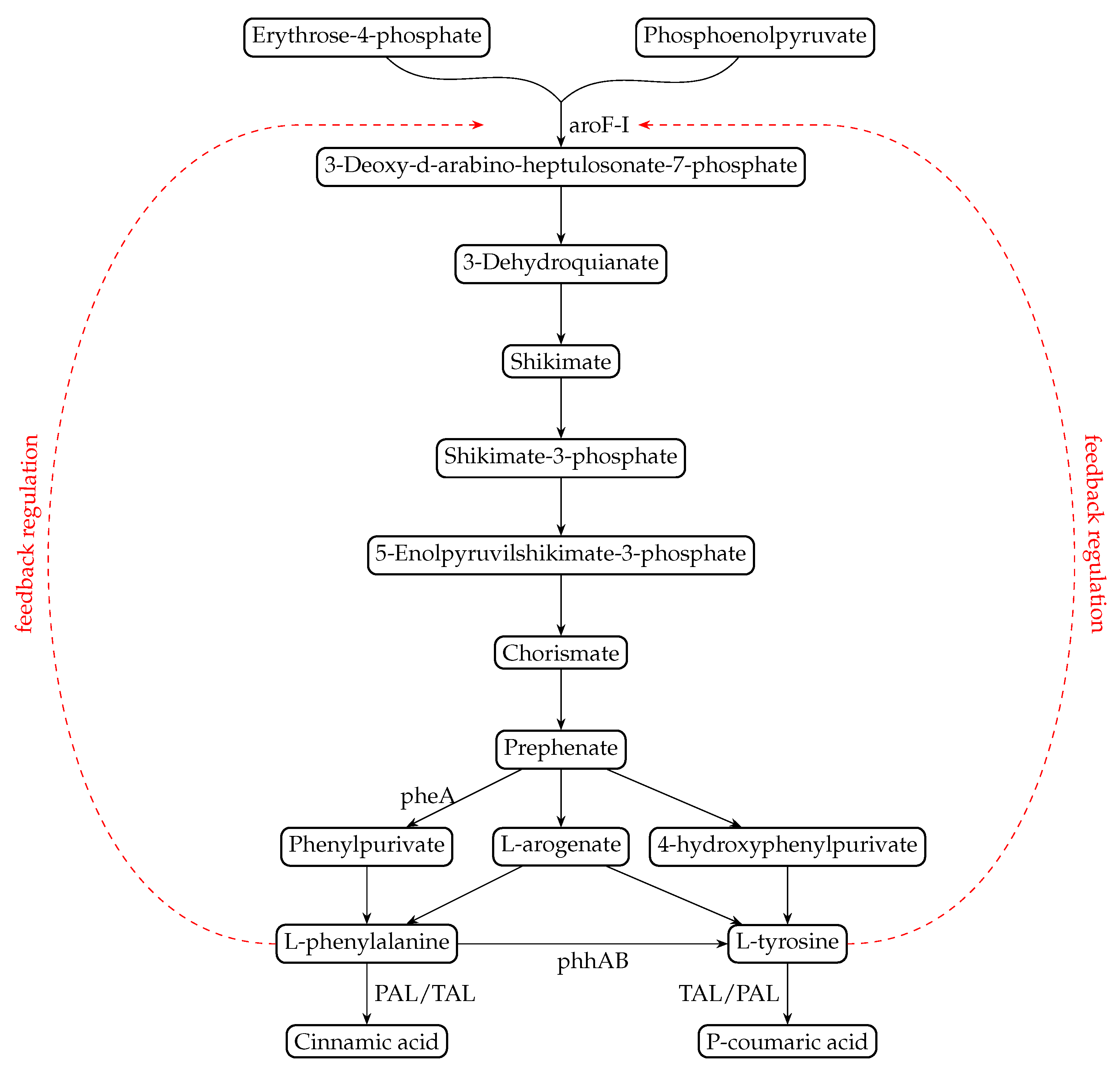
1. Introduction
2. Results
2.1. AroF-I from Pseudomonas putida Is Highly Sensitive to and Inhibited by Tyrosine
2.2. Bioinformatics Helps in the Prediction of Allosteric Inhibition of the AroF-I Enzyme
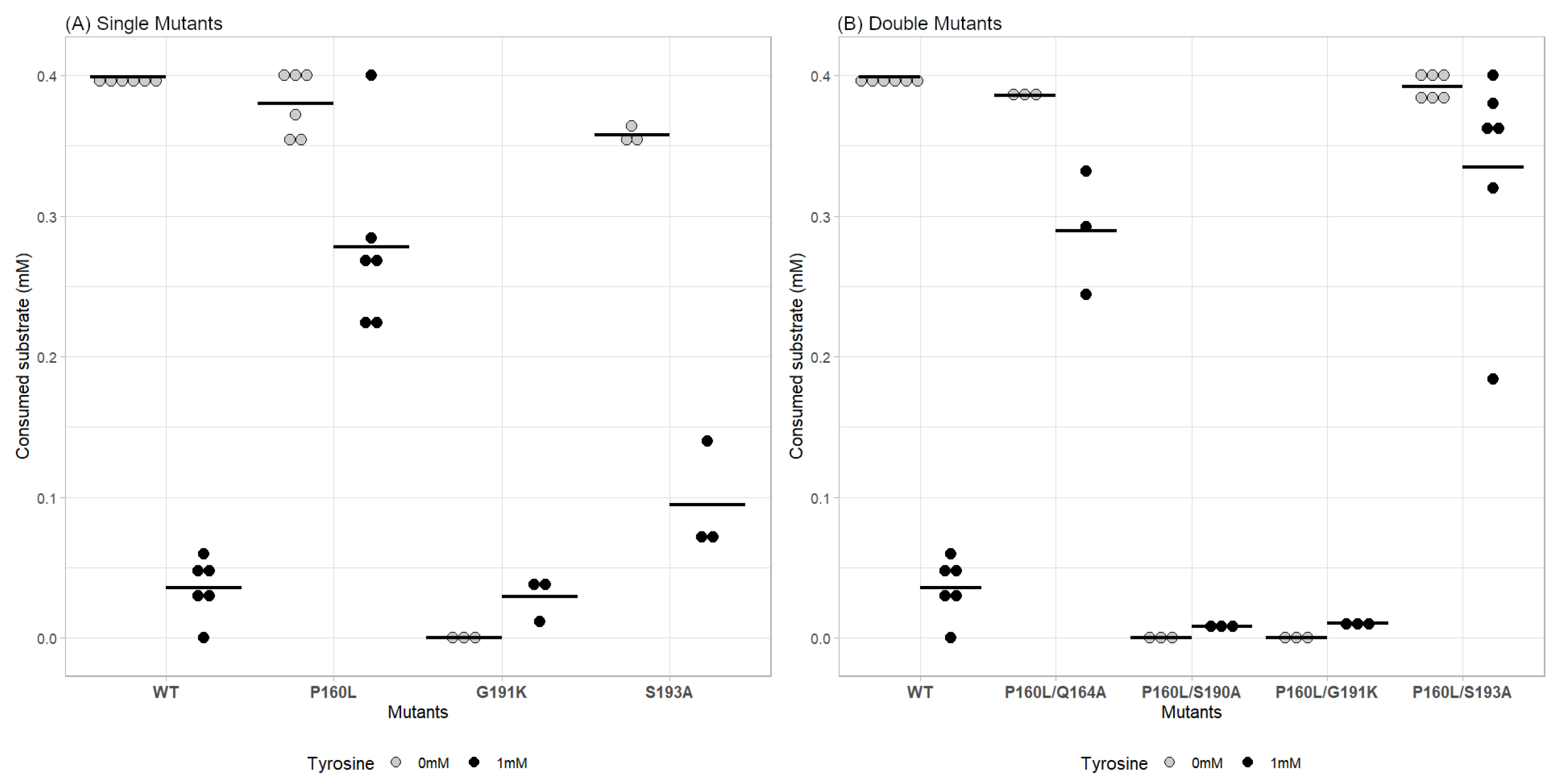
2.3. Effect of Point Mutations on AroF-I Activity in the Presence or Absence of an Inhibitor
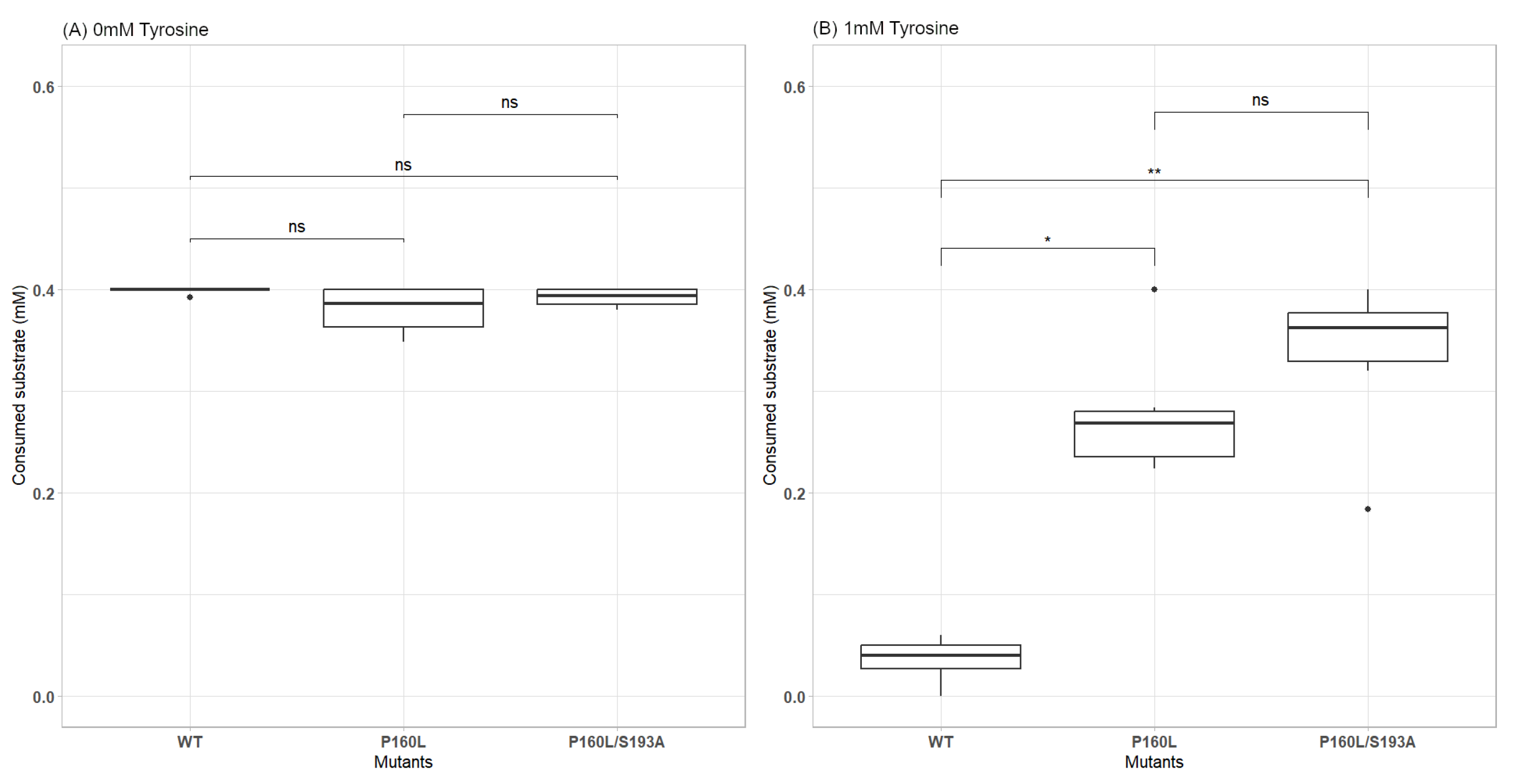
2.4. A Double Mutation Improves the Tyrosine Resistance of AroF-I and Its Enzymatic Activity
2.5. Construction of Pseudomonas putida Strains Overproducing Aromatic Amino Acids
2.6. A Synergistic Effect of AroF-I Fbr and phhAB Improves Production of p-Coumaric Acid
3. Materials and Methods
3.1. Bacterial Strains, Plasmids, and Growth Conditions
3.2. Vector Construction and Strain Engineering
3.3. Production and Purification of the AroF-I Proteins
3.4. Enzymatic Assay for Determination of AroF-I Activity
3.5. Replacement of aroH by Mutated AroF-I
3.6. Overexpression of phhAB Genes
3.7. Assessment of pCA Production in Batch Culture
3.8. Statistical Analysis
3.9. Bioinformatics Protocols
4. Discussion
5. Patents
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | OD600 | [pCA] (mM) | [CA] (mM) | pCA/1DO | CA/1DO | Total Phenolics/1DO |
|---|---|---|---|---|---|---|
| Ppu201 | 5.4 | 0.28 | 0.14 | 0.05 | 0.003 | 0.08 |
| Ppu201/CP | 3.4 | 1.95 | 0.12 | 0.57 | 0.04 | 0.61 |
| Ppu201/PP | 3.4 | 1.99 | 0.13 | 0.59 | 0.04 | 0.62 |
| Ppu235 | 4.1 | 1.94 | 0.66 | 0.47 | 0.16 | 0.63 |
| Ppu235/CP | 3.8 | 2.38 | 0.38 | 0.63 | 0.10 | 0.72 |
| Ppu235/PP | 3.7 | 2.46 | 0.37 | 0.66 | 0.10 | 0.76 |
| Ppu201 | 4.6 | 0.28 | 0.14 | 0.06 | 0.03 | 0.09 |
| Ppu201/CP | 4.6 | 1.83 | 0.15 | 0.40 | 0.03 | 0.43 |
| Ppu201/PP | 3.1 | 1.80 | 0.03 | 0.58 | 0.01 | 0.59 |
| Ppu235 | 3.4 | 1.92 | 0.66 | 0.56 | 0.20 | 0.76 |
| Ppu235/CP | 5.1 | 2.07 | 0.30 | 0.41 | 0.06 | 0.46 |
| Ppu235/PP | 1.6 | 2.21 | 0.07 | 1.38 | 0.04 | 1.42 |
| Ppu201 | 5.1 | 0.28 | 0.14 | 0.06 | 0.03 | 0.08 |
| Ppu201/CP | 3.3 | 1.92 | 0.13 | 0.58 | 0.04 | 0.62 |
| Ppu201/PP | 3.4 | 2.06 | 0.13 | 0.61 | 0.04 | 0.64 |
| Ppu235 | 3.9 | 1.93 | 0.65 | 0.49 | 0.17 | 0.66 |
| Ppu235/CP | 3.7 | 2.37 | 0.37 | 0.64 | 0.10 | 0.74 |
| Ppu235/PP | 3.6 | 2.39 | 0.37 | 0.66 | 0.10 | 0.77 |
| Ppu201 | 4.3 | 0.27 | 0.14 | 0.06 | 0.03 | 0.09 |
| Ppu201/CP | 4.1 | 1.82 | 0.14 | 0.44 | 0.03 | 0.48 |
| Ppu201/PP | 3.1 | 1.76 | 0.03 | 0.57 | 0.01 | 0.58 |
| Ppu235 | 3.8 | 1.83 | 0.65 | 0.48 | 0.17 | 0.65 |
| Ppu235/CP | 4.6 | 2.19 | 0.36 | 0.48 | 0.08 | 0.55 |
| Ppu235/PP | 1.1 | 2.26 | 0.06 | 2.05 | 0.05 | 2.10 |
| Ppu201 | 5 | 0.28 | 0.14 | 0.06 | 0.03 | 0.09 |
| Ppu201/CP | 3.5 | 1.94 | 0.13 | 0.55 | 0.04 | 0.59 |
| Ppu201/PP | 3.4 | 2.04 | 0.13 | 0.60 | 0.04 | 0.64 |
| Ppu235 | 3.9 | 2.00 | 0.64 | 0.51 | 0.17 | 0.68 |
| Ppu235/CP | 3.8 | 2.40 | 0.37 | 0.63 | 0.10 | 0.73 |
| Ppu235/PP | 3.6 | 2.44 | 0.36 | 0.68 | 0.10 | 0.78 |
| Ppu201 | 4.4 | 0.27 | 0.14 | 0.06 | 0.03 | 0.09 |
| Ppu201/CP | 4.7 | 1.90 | 0.15 | 0.41 | 0.03 | 0.44 |
| Ppu201/PP | 3.2 | 1.85 | 0.03 | 0.58 | 0.01 | 0.59 |
| Ppu235 | 3.5 | 1.98 | 0.68 | 0.56 | 0.20 | 0.76 |
| Ppu235/CP | 5.4 | 2.45 | 0.39 | 0.45 | 0.07 | 0.53 |
| Ppu235/PP | 1.6 | 2.37 | 0.07 | 1.48 | 0.04 | 1.52 |
| Ppu201 | 3.7 | 1.5 | 0.30 | 0.41 | 0.08 | 0.49 |
| Ppu235 | 4 | 1.3 | 0.80 | 0.32 | 0.20 | 0.52 |
| Ppu201/PP | 3.3 | 1.4 | 0.10 | 0.42 | 0.03 | 0.45 |
| Ppu235/PP | 3.6 | 1.8 | 0.36 | 0.50 | 0.10 | 0.60 |
| Ppu201 | 3.8 | 1.4 | 0.30 | 0.38 | 0.08 | 0.46 |
| Ppu235 | 3.9 | 1.3 | 0.74 | 0.33 | 0.19 | 0.52 |
| Ppu201/PP | 0.4 | 0.5 | 0.01 | 1.26 | 0.03 | 1.29 |
| Ppu235/PP | 0.4 | 0.8 | 0.02 | 2.08 | 0.05 | 2.13 |
| Ppu201 | 4 | 1.5 | 0.28 | 0.38 | 0.07 | 0.45 |
| Ppu235/PP | 4.2 | 1.4 | 0.76 | 0.34 | 0.18 | 0.52 |
| Ppu201/PP | 3.6 | 1.4 | 0.11 | 0.40 | 0.03 | 0.43 |
| Ppu235/PP | 4 | 1.8 | 0.32 | 0.46 | 0.08 | 0.54 |
| Ppu201 | 4 | 1.5 | 0.32 | 0.38 | 0.08 | 0.46 |
| Ppu235 | 4.1 | 1.5 | 0.74 | 0.36 | 0.18 | 0.54 |
| Ppu201/PP | 0.5 | 0.5 | 0.01 | 0.99 | 0.02 | 1.01 |
| Ppu235/PP | 0.8 | 1.1 | 0.02 | 1.40 | 0.03 | 1.43 |
| Ppu201 | 4.5 | 2.2 | 0.32 | 0.49 | 0.07 | 0.56 |
| Ppu235 | 4.6 | 2.1 | 0.74 | 0.45 | 0.16 | 0.61 |
| Ppu235/PP | 4 | 2.2 | 0.36 | 0.56 | 0.09 | 0.65 |
| Ppu201 | 5.1 | 3.0 | 0.46 | 0.58 | 0.09 | 0.67 |
| Ppu235 | 5.4 | 2.5 | 0.86 | 0.47 | 0.16 | 0.63 |
| Ppu235/PP | 0.5 | 1.2 | 0.04 | 2.43 | 0.08 | 2.51 |
References
- Kitney, R.; Freemont, P. Synthetic biology—The state of play. FEBS Lett. 2012, 586, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Martău, G.A.; Călinoiu, L.F.; Vodnar, D.C. Bio-vanillin: Towards a sustainable industrial production. Trends Food Sci. Technol. 2021, 109, 579–592. [Google Scholar] [CrossRef]
- Wang, J.; Shen, X.; Rey, J.; Yuan, Q.; Yan, Y. Recent advances in microbial production of aromatic natural products and their derivatives. Appl. Microbiol. Biotechnol. 2018, 102, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Dudnik, A.; Gaspar, P.; Neves, A.R.; Forster, J. Engineering of Microbial Cell Factories for the Production of Plant Polyphenols with Health-Beneficial Properties. Curr. Pharm. Des. 2018, 24, 2208–2225. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Yahya, P.P.; Zhang, F.; del Cardayre, S.B.; Keasling, J.D. Microbial engineering for the production of advanced biofuels. Nature 2012, 488, 320–328. [Google Scholar] [CrossRef]
- Aggarwal, N.; Pham, H.L.; Ranjan, B.; Saini, M.; Liang, Y.; Hossain, G.S.; Ling, H.; Foo, J.L.; Chang, M.W. Microbial engineering strategies to utilize waste feedstock for sustainable bioproduction. Nat. Rev. Bioeng. 2023, 2, 155–174. [Google Scholar] [CrossRef]
- Li, B.; Zhang, B.; Wang, P.; Cai, X.; Chen, Y.Y.; Yang, Y.F.; Liu, Z.Q.; Zheng, Y.G. Rerouting Fluxes of the Central Carbon Metabolism and Relieving Mechanism-Based Inactivation of l-Aspartate-α-decarboxylase for Fermentative Production of β-Alanine in Escherichia coli. Acs Synth. Biol. 2022, 11, 1908–1918. [Google Scholar] [CrossRef]
- Maurya, R.; Gohil, N.; Nixon, S.; Kumar, N.; Noronha, S.B.; Dhali, D.; Trabelsi, H.; Alzahrani, K.J.; Reshamwala, S.M.; Awasthi, M.K.; et al. Rewiring of metabolic pathways in yeasts for sustainable production of biofuels. Bioresour. Technol. 2023, 372, 128668. [Google Scholar] [CrossRef]
- Leonard, E.; Lim, K.H.; Saw, P.N.; Koffas, M.A.G. Engineering Central Metabolic Pathways for High-Level Flavonoid Production in Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 3877–3886. [Google Scholar] [CrossRef]
- Suzuki, S.; Koeduka, T.; Sugiyama, A.; Yazaki, K.; Umezawa, T. Microbial production of plant specialized metabolites. Plant Biotechnol. 2014, 31, 465–482. [Google Scholar] [CrossRef]
- Limem, I.; Guedon, E.; Hehn, A.; Bourgaud, F.; Chekir Ghedira, L.; Engasser, J.M.; Ghoul, M. Production of phenylpropanoid compounds by recombinant microorganisms expressing plant-specific biosynthesis genes. Process Biochem. 2008, 43, 463–479. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, S.; Xiao, A.; Rasmussen, M.; Skidmore, C.; Zhan, J. Metabolic engineering of Escherichia coli for the biosynthesis of various phenylpropanoid derivatives. Metab. Eng. 2015, 29, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Averesch, N.J.H.; Krömer, J.O. Metabolic Engineering of the Shikimate Pathway for Production of Aromatics and Derived Compounds—Present and Future Strain Construction Strategies. Front. Bioeng. Biotechnol. 2018, 6, 32. [Google Scholar] [CrossRef]
- Vargas-Tah, A.; Gosset, G. Production of Cinnamic and p-Hydroxycinnamic Acids in Engineered Microbes. Front. Bioeng. Biotechnol. 2015, 3, 116. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. The Shikimate Pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Jayaraman, K.; Trachtmann, N.; Sprenger, G.A.; Gohlke, H. Protein engineering for feedback resistance in 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase. Appl. Microbiol. Biotechnol. 2022, 106, 6505–6517. [Google Scholar] [CrossRef]
- Wu, J.; Howe, D.L.; Woodard, R.W. Thermotoga maritima 3-Deoxy-D-arabino-heptulosonate 7-Phosphate (DAHP) Synthase. J. Biol. Chem. 2003, 278, 27525–27531. [Google Scholar] [CrossRef]
- Luttik, M.; Vuralhan, Z.; Suir, E.; Braus, G.; Pronk, J.; Daran, J. Alleviation of feedback inhibition in Saccharomyces cerevisiae aromatic amino acid biosynthesis: Quantification of metabolic impact. Metab. Eng. 2008, 10, 141–153. [Google Scholar] [CrossRef]
- Bilal, M.; Wang, S.; Iqbal, H.M.N.; Zhao, Y.; Hu, H.; Wang, W.; Zhang, X. Metabolic engineering strategies for enhanced shikimate biosynthesis: Current scenario and future developments. Appl. Microbiol. Biotechnol. 2018, 102, 7759–7773. [Google Scholar] [CrossRef]
- Lütke-Eversloh, T.; Stephanopoulos, G. L-Tyrosine production by deregulated strains of Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 75, 103–110. [Google Scholar] [CrossRef]
- Kang, S.Y.; Choi, O.; Lee, J.K.; Hwang, B.Y.; Uhm, T.B.; Hong, Y.S. Artificial biosynthesis of phenylpropanoic acids in a tyrosine overproducing Escherichia coli strain. Microb. Cell Factories 2012, 11, 153. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.N.S.; Koffas, M.; Stephanopoulos, G. Optimization of a heterologous pathway for the production of flavonoids from glucose. Metab. Eng. 2011, 13, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Juminaga, D.; Baidoo, E.E.K.; Redding-Johanson, A.M.; Batth, T.S.; Burd, H.; Mukhopadhyay, A.; Petzold, C.J.; Keasling, J.D. Modular Engineering of l-Tyrosine Production in Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Kildegaard, K.R.; Li, M.; Borodina, I.; Nielsen, J. Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis. Metab. Eng. 2015, 31, 181–188. [Google Scholar] [CrossRef]
- Nikel, P.I.; Martínez-García, E.; de Lorenzo, V. Biotechnological domestication of pseudomonads using synthetic biology. Nat. Rev. Microbiol. 2014, 12, 368–379. [Google Scholar] [CrossRef]
- Ankenbauer, A.; Schäfer, R.A.; Viegas, S.C.; Pobre, V.; Voß, B.; Arraiano, C.M.; Takors, R. Pseudomonas putida KT2440 is naturally endowed to withstand industrial-scale stress conditions. Microb. Biotechnol. 2020, 13, 1145–1161. [Google Scholar] [CrossRef]
- Schwanemann, T.; Otto, M.; Wierckx, N.; Wynands, B. Pseudomonas as Versatile Aromatics Cell Factory. Biotechnol. J. 2020, 15, 1900569. [Google Scholar] [CrossRef]
- Molina-Santiago, C.; Cordero, B.F.; Daddaoua, A.; Udaondo, Z.; Manzano, J.; Valdivia, M.; Segura, A.; Ramos, J.L.; Duque, E. Pseudomonas putida as a platform for the synthesis of aromatic compounds. Microbiology 2016, 162, 1535–1543. [Google Scholar] [CrossRef]
- Calero, P.; Jensen, S.I.; Bojanovič, K.; Lennen, R.M.; Koza, A.; Nielsen, A.T. Genome-wide identification of tolerance mechanisms toward p-coumaric acid in Pseudomonas putida. Biotechnol. Bioeng. 2017, 115, 762–774. [Google Scholar] [CrossRef]
- Mohamed, E.T.; Werner, A.Z.; Salvachúa, D.; Singer, C.A.; Szostkiewicz, K.; Rafael Jiménez-Díaz, M.; Eng, T.; Radi, M.S.; Simmons, B.A.; Mukhopadhyay, A.; et al. Adaptive laboratory evolution of Pseudomonas putida KT2440 improves p-coumaric and ferulic acid catabolism and tolerance. Metab. Eng. Commun. 2020, 11, e00143. [Google Scholar] [CrossRef]
- Nijkamp, K.; Westerhof, R.G.M.; Ballerstedt, H.; de Bont, J.A.M.; Wery, J. Optimization of the solvent-tolerant Pseudomonas putida S12 as host for the production of p-coumarate from glucose. Appl. Microbiol. Biotechnol. 2007, 74, 617–624. [Google Scholar] [CrossRef]
- Wierckx, N.J.P.; Ballerstedt, H.; de Bont, J.A.M.; Wery, J. Engineering of Solvent-Tolerant Pseudomonas putida S12 for Bioproduction of Phenol from Glucose. Appl. Environ. Microbiol. 2005, 71, 8221–8227. [Google Scholar] [CrossRef] [PubMed]
- Calero, P.; Jensen, S.I.; Nielsen, A.T. Broad-Host-Range ProUSER Vectors Enable Fast Characterization of Inducible Promoters and Optimization of p-Coumaric Acid Production in Pseudomonas putida KT2440. ACS Synth. Biol. 2016, 5, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Wynands, B.; Lenzen, C.; Otto, M.; Koch, F.; Blank, L.M.; Wierckx, N. Metabolic engineering of Pseudomonas taiwanensis VLB120 with minimal genomic modifications for high-yield phenol production. Metab. Eng. 2018, 47, 121–133. [Google Scholar] [CrossRef]
- Otto, M.; Wynands, B.; Lenzen, C.; Filbig, M.; Blank, L.M.; Wierckx, N. Rational Engineering of Phenylalanine Accumulation in Pseudomonas taiwanensis to Enable High-Yield Production of Trans-Cinnamate. Front. Bioeng. Biotechnol. 2019, 7, 312. [Google Scholar] [CrossRef]
- Weaver, L.M.; Herrmann, K.M. Cloning of an aroF allele encoding a tyrosine-insensitive 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase. J. Bacteriol. 1990, 172, 6581–6584. [Google Scholar] [CrossRef]
- Lütke-Eversloh, T.; Stephanopoulos, G. Feedback inhibition of chorismate mutase/prephenate dehydrogenase (TyrA) of Escherichia coli: Generation and characterization of tyrosine-insensitive mutants. Appl. Environ. Microbiol. 2005, 71, 7224–7228. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J.; Liu, Z.; Qiao, Q.; Ni, X.; Yang, J.; Sun, G.; Li, F.; Zhou, W.; Guo, X.; et al. Engineering allosteric inhibition of homoserine dehydrogenase by semi-rational saturation mutagenesis screening. Front. Bioeng. Biotechnol. 2024, 11, 1336215. [Google Scholar] [CrossRef]
- Chen, M.M.; Snow, C.D.; Vizcarra, C.L.; Mayo, S.L.; Arnold, F.H. Comparison of random mutagenesis and semi-rational designed libraries for improved cytochrome P450 BM3-catalyzed hydroxylation of small alkanes. Protein Eng. Des. Sel. 2012, 25, 171–178. [Google Scholar] [CrossRef]
- Rowlands, R. Industrial strain improvement: Mutagenesis and random screening procedures. Enzym. Microb. Technol. 1984, 6, 3–10. [Google Scholar] [CrossRef]
- Zhao, S.; Tan, M.Z.; Wang, R.X.; Ye, F.T.; Chen, Y.P.; Luo, X.M.; Feng, J.X. Combination of genetic engineering and random mutagenesis for improving production of raw-starch-degrading enzymes in Penicillium oxalicum. Microb. Cell Factories 2022, 21, 272. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Mildvan, A.S. Inverse thinking about double mutants of enzymes. Biochemistry 2004, 43, 14517–14520. [Google Scholar] [CrossRef] [PubMed]
- Chaparro-Riggers, J.F.; Polizzi, K.M.; Bommarius, A.S. Better library design: Data-driven protein engineering. Biotechnol. J. Healthc. Nutr. Technol. 2007, 2, 180–191. [Google Scholar] [CrossRef]
- Wierckx, N.; Ruijssenaars, H.J.; de Winde, J.H.; Schmid, A.; Blank, L.M. Metabolic flux analysis of a phenol producing mutant of Pseudomonas putida S12: Verification and complementation of hypotheses derived from transcriptomics. J. Biotechnol. 2009, 143, 124–129. [Google Scholar] [CrossRef]
- Glick, B.R. Metabolic load and heterologous gene expression. Biotechnol. Adv. 1995, 13, 247–261. [Google Scholar] [CrossRef]
- Bertani, G. STUDIES ON LYSOGENESIS I: The Mode of Phage Liberation by Lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop II, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Studier, F.W. Protein production by auto-induction in high-density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G. Synthesis of DNA fragments in yeast by one-step assembly of overlapping oligonucleotides. Nucleic Acids Res. 2009, 37, 6984–6990. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Jones, D.H.; Snedecor, G.W.; Cochran, W.G. Statistical Methods. J. Educ. Behav. Stat. 1994, 19, 304. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple Comparisons Using Rank Sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge: London, UK, 2013. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Shumilin, I.A.; Zhao, C.; Bauerle, R.; Kretsinger, R.H. Allosteric Inhibition of 3-Deoxy-d-arabino-heptulosonate-7-phosphate Synthase Alters the Coordination of Both Substrates. J. Mol. Biol. 2002, 320, 1147–1156. [Google Scholar] [CrossRef]
- Cui, D.; Deng, A.; Bai, H.; Yang, Z.; Liang, Y.; Liu, Z.; Qiu, Q.; Wang, L.; Liu, S.; Zhang, Y.; et al. Molecular basis for feedback inhibition of tyrosine-regulated 3-deoxy-d-arabino-heptulosonate-7-phosphate synthase from Escherichia coli. J. Struct. Biol. 2019, 206, 322–334. [Google Scholar] [CrossRef]
- Ding, R.; Liu, L.; Chen, X.; Cui, Z.; Zhang, A.; Ren, D.; Zhang, L. Introduction of two mutations into AroG increases phenylalanine production in Escherichia coli. Biotechnol. Lett. 2014, 36, 2103–2108. [Google Scholar] [CrossRef]
- Shimizu, K. Metabolic Regulation of a Bacterial Cell System with Emphasis on Escherichia coli Metabolism. ISRN Biochem. 2013, 2013, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Chubukov, V.; Gerosa, L.; Kochanowski, K.; Sauer, U. Coordination of microbial metabolism. Nat. Rev. Microbiol. 2014, 12, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Matsuoka, Y. Feedback regulation and coordination of the main metabolism for bacterial growth and metabolic engineering for amino acid fermentation. Biotechnol. Adv. 2022, 55, 107887. [Google Scholar] [CrossRef] [PubMed]
- Nudler, E. The riboswitch control of bacterial metabolism. Trends Biochem. Sci. 2004, 29, 11–17. [Google Scholar] [CrossRef]
- Wu, S.; Chen, W.; Lu, S.; Zhang, H.; Yin, L. Metabolic Engineering of Shikimic Acid Biosynthesis Pathway for the Production of Shikimic Acid and Its Branched Products in Microorganisms: Advances and Prospects. Molecules 2022, 27, 4779. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, H. Engineering the shikimate pathway for biosynthesis of molecules with pharmaceutical activities in E. coli. Curr. Opin. Biotechnol. 2016, 42, 1–6. [Google Scholar] [CrossRef]
- Cao, M.; Gao, M.; Suástegui, M.; Mei, Y.; Shao, Z. Building microbial factories for the production of aromatic amino acid pathway derivatives: From commodity chemicals to plant-sourced natural products. Metab. Eng. 2020, 58, 94–132. [Google Scholar] [CrossRef]
- Helmstaedt, K.; Strittmatter, A.; Lipscomb, W.N.; Braus, G.H. Evolution of 3-deoxy-d-arabino-heptulosonate-7-phosphate synthase-encoding genes in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 9784–9789. [Google Scholar] [CrossRef]
- Jossek, R.; Bongaerts, J.; Sprenger, G.A. Characterization of a new feedback-resistant 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase AroF of Escherichia coli. FEMS Microbiol. Lett. 2001, 202, 145–148. [Google Scholar] [CrossRef]
- Mir, R.; Jallu, S.; Singh, T.P. The shikimate pathway: Review of amino acid sequence, function and three-dimensional structures of the enzymes. Crit. Rev. Microbiol. 2013, 41, 172–189. [Google Scholar] [CrossRef]
- Ger, Y.M.; Chen, S.L.; Chiang, H.J.; Shiuan, D. A Single Ser-180 Mutation Desensitizes Feedback Inhibition of the Phenylalanine-Sensitive 3-Deoxy-D-Arabino-Heptulosonate 7-Phosphate (DAHP) Synthetase in Escherichia coli. J. Biochem. 1994, 116, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.M.; Yanofsky, C.; Bauerle, R. Mutational analysis of the catalytic and feedback sites of the tryptophan-sensitive 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase of Escherichia coli. J. Bacteriol. 1988, 170, 5500–5506. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, Y.; Tsujimoto, K.; Kurahashi, O. Mutational analysis of the feedback sites of phenylalanine-sensitive 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase of Escherichia coli. Appl. Environ. Microbiol. 1997, 63, 761–762. [Google Scholar] [CrossRef] [PubMed]
- Lutz, S. Beyond directed evolution—Semi-rational protein engineering and design. Curr. Opin. Biotechnol. 2010, 21, 734–743. [Google Scholar] [CrossRef]
- Chen, H.; Ma, L.; Dai, H.; Fu, Y.; Wang, H.; Zhang, Y. Advances in Rational Protein Engineering toward Functional Architectures and Their Applications in Food Science. J. Agric. Food Chem. 2022, 70, 4522–4533. [Google Scholar] [CrossRef]
- Nixon, A.E.; Firestine, S.M. Rational and “Irrational” Design of Proteins and Their Use in Biotechnology. IUBMB Life 2000, 49, 181–187. [Google Scholar] [CrossRef]
- Faisca, P.; Travasso, R.; Ball, R.; Shakhnovich, E.I. Identifying critical residues in protein folding: Insights from ϕ-value and Pfold analysis. J. Chem. Phys. 2008, 129, 095108. [Google Scholar] [CrossRef]
- Sen, C.; Logashree, V.; Makde, R.D.; Ghosh, B. Amino acid propensities for secondary structures and its variation across protein structures using exhaustive PDB data. Comput. Biol. Chem. 2024, 110, 108083. [Google Scholar] [CrossRef]
- Chronopoulou, E.G.; Labrou, N.E. Site-saturation mutagenesis: A powerful tool for structure-based design of combinatorial mutation libraries. Curr. Protoc. Protein Sci. 2011, 63. [Google Scholar] [CrossRef]
- Marabotti, A.; Scafuri, B.; Facchiano, A. Predicting the stability of mutant proteins by computational approaches: An overview. Briefings Bioinform. 2021, 22, bbaa074. [Google Scholar] [CrossRef]
- Pucci, F.; Schwersensky, M.; Rooman, M. Artificial intelligence challenges for predicting the impact of mutations on protein stability. Curr. Opin. Struct. Biol. 2022, 72, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. A critical review of five machine learning-based algorithms for predicting protein stability changes upon mutation. Briefings Bioinform. 2019, 21, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Mao, C.; Tang, J.; Yang, S.; Cheng, Y.; Wang, W.; Gu, Q.; Han, W.; Chen, H.; Li, S.; et al. Zero-shot prediction of mutation effects with multimodal deep representation learning guides protein engineering. Cell Res. 2024, 34, 630–647. [Google Scholar] [CrossRef] [PubMed]
- Hamborg, L.; Granata, D.; Olsen, J.G.; Roche, J.V.; Pedersen, L.E.; Nielsen, A.T.; Lindorff-Larsen, K.; Teilum, K. Synergistic stabilization of a double mutant in chymotrypsin inhibitor 2 from a library screen in E. coli. Commun. Biol. 2021, 4, 980. [Google Scholar] [CrossRef]
- Greener, J.G.; Sternberg, M.J. AlloPred: Prediction of allosteric pockets on proteins using normal mode perturbation analysis. BMC Bioinform. 2015, 16, 335. [Google Scholar] [CrossRef]
- Yamato, T.; Laprévote, O. Normal mode analysis and beyond. Biophys. Physicobiol. 2019, 16, 322–327. [Google Scholar] [CrossRef]
- Kolossváry, I. A Fresh Look at the Normal Mode Analysis of Proteins: Introducing Allosteric Co-Vibrational Modes. JACS Au 2024, 4, 1303–1309. [Google Scholar] [CrossRef]
- Ruffolo, J.A.; Madani, A. Designing proteins with language models. Nat. Biotechnol. 2024, 42, 200–202. [Google Scholar] [CrossRef]
- Schmirler, R.; Heinzinger, M.; Rost, B. Fine-tuning protein language models boosts predictions across diverse tasks. Nat. Commun. 2024, 15, 7407. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, L.; Yu, Y.; Wu, B.; Li, M.; Hong, L.; Tan, P. Enhancing efficiency of protein language models with minimal wet-lab data through few-shot learning. Nat. Commun. 2024, 15, 5566. [Google Scholar] [CrossRef]
- Biswas, S.; Khimulya, G.; Alley, E.C.; Esvelt, K.M.; Church, G.M. Low-N protein engineering with data-efficient deep learning. Nat. Methods 2021, 18, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Chen, R. Bacterial expression systems for recombinant protein production: E. coli and beyond. Biotechnol. Adv. 2012, 30, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Goussé, M.; Dell’Aglio, E.; Curien, G.; Borland, S.; Renoud, S.; Ranquet, C.; Chandor-Proust, A. E. coli chromosomal-driven expression of NADK2 from A. thaliana: A preferable alternative to plasmid-driven expression for challenging proteins. Protein Expr. Purif. 2022, 195–196, 106090. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.H.; Jayaram, M.; Rowley, P.A. Metabolic engineering without plasmids. Nat. Biotechnol. 2009, 27, 729–731. [Google Scholar] [CrossRef]
- Zucca, S.; Pasotti, L.; Mazzini, G.; Cusella De Angelis, M.G.; Magni, P. Characterization of an inducible promoter in different DNA copy number conditions. BMC Bioinform. 2012, 13, S11. [Google Scholar] [CrossRef]
- Tyo, K.E.J.; Ajikumar, P.K.; Stephanopoulos, G. Stabilized gene duplication enables long-term selection-free heterologous pathway expression. Nat. Biotechnol. 2009, 27, 760–765. [Google Scholar] [CrossRef]
- Schild, F.; Chandor-Proust, A.; Ranquet, C. Method of Mutagenesis Without Scar; Institut National de la Propriété’ Industrielle: Paris, France, 2017. [Google Scholar]
- Li, Z.; Wang, H.; Ding, D.; Liu, Y.; Fang, H.; Chang, Z.; Chen, T.; Zhang, D. Metabolic engineering of Escherichia coli for production of chemicals derived from the shikimate pathway. J. Ind. Microbiol. Biotechnol. 2020, 47, 525–535. [Google Scholar] [CrossRef]
- Bentley, R.; Haslam, E. The Shikimate Pathway—A Metabolic Tree with Many Branche. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 307–384. [Google Scholar] [CrossRef]
- Liu, Q.; Yu, T.; Li, X.; Chen, Y.; Campbell, K.; Nielsen, J.; Chen, Y. Rewiring carbon metabolism in yeast for high level production of aromatic chemicals. Nat. Commun. 2019, 10, 4976. [Google Scholar] [CrossRef]
- Kroll, K.; Holland, C.K.; Starks, C.M.; Jez, J.M. Evolution of allosteric regulation in chorismate mutases from early plants. Biochem. J. 2017, 474, 3705–3717. [Google Scholar] [CrossRef]
- Brown, J.F.; Dawes, I.W. Regulation of chorismate mutase in Saccharomyces cerevisiae. Mol. Gen. Genet. MGG 1990, 220, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pohnert, G.; Kongsaeree, P.; Wilson, D.B.; Clardy, J.; Ganem, B. Chorismate Mutase-Prephenate Dehydratase from Escherichia coli. J. Biol. Chem. 1998, 273, 6248–6253. [Google Scholar] [CrossRef] [PubMed]
- Mokwatlo, S.C.; Klein, B.C.; Benavides, P.T.; Tan, E.C.; Kneucker, C.M.; Ling, C.; Singer, C.A.; Lyons, R.; i Nogué, V.S.; Hestmark, K.V.; et al. Bioprocess development and scale-up for cis, cis-muconic acid production from glucose and xylose by Pseudomonas putida. Green Chem. 2024, 26, 10152–10167. [Google Scholar] [CrossRef]
- Flourat, A.L.; Combes, J.; Bailly-Maitre-Grand, C.; Magnien, K.; Haudrechy, A.; Renault, J.H.; Allais, F. Accessing p-Hydroxycinnamic Acids: Chemical Synthesis, Biomass Recovery, or Engineered Microbial Production? ChemSusChem 2021, 14, 118–129. [Google Scholar] [CrossRef]
- Louie, G.V.; Bowman, M.E.; Moffitt, M.C.; Baiga, T.J.; Moore, B.S.; Noel, J.P. Structural Determinants and Modulation of Substrate Specificity in Phenylalanine-Tyrosine Ammonia-Lyases. Chem. Biol. 2006, 13, 1327–1338. [Google Scholar] [CrossRef]
- Watts, K.T.; Mijts, B.N.; Lee, P.C.; Manning, A.J.; Schmidt-Dannert, C. Discovery of a Substrate Selectivity Switch in Tyrosine Ammonia-Lyase, a Member of the Aromatic Amino Acid Lyase Family. Chem. Biol. 2006, 13, 1317–1326. [Google Scholar] [CrossRef]
- Jendresen, C.B.; Stahlhut, S.G.; Li, M.; Gaspar, P.; Siedler, S.; Förster, J.; Maury, J.; Borodina, I.; Nielsen, A.T. Highly Active and Specific Tyrosine Ammonia-Lyases from Diverse Origins Enable Enhanced Production of Aromatic Compounds in Bacteria and Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 4458–4476. [Google Scholar] [CrossRef]
- KeGG Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.kegg.jp/kegg/ (accessed on 15 November 2024).

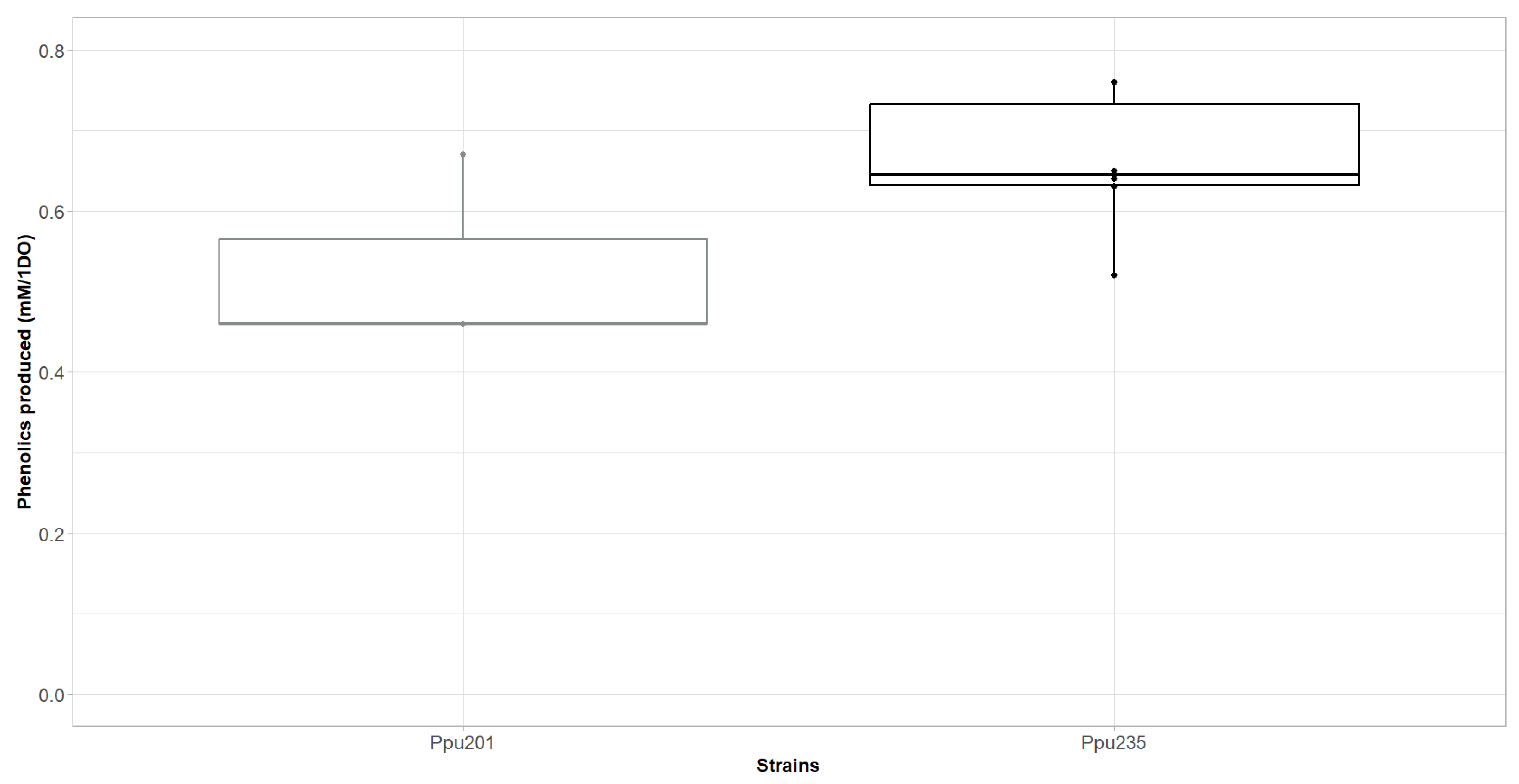

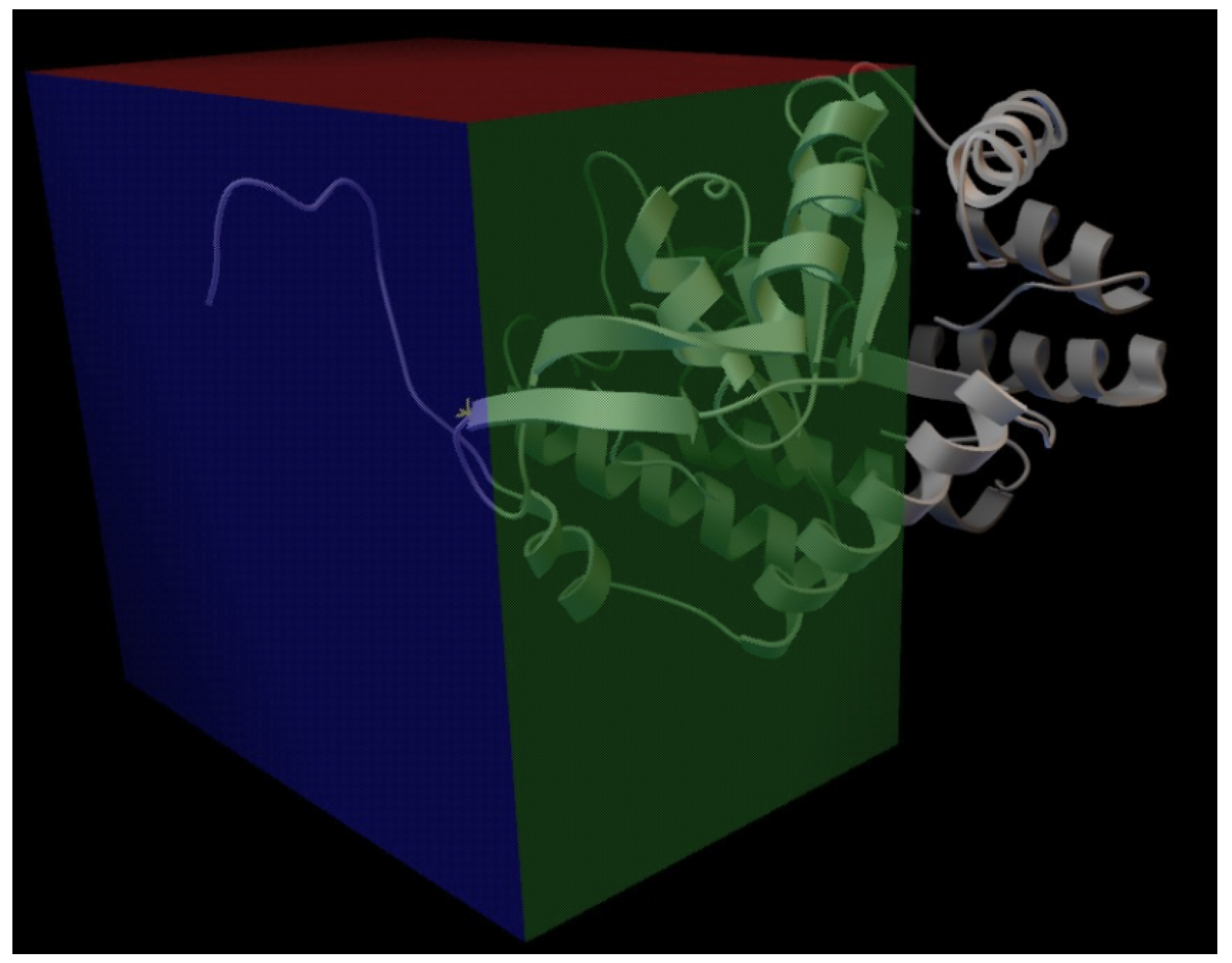
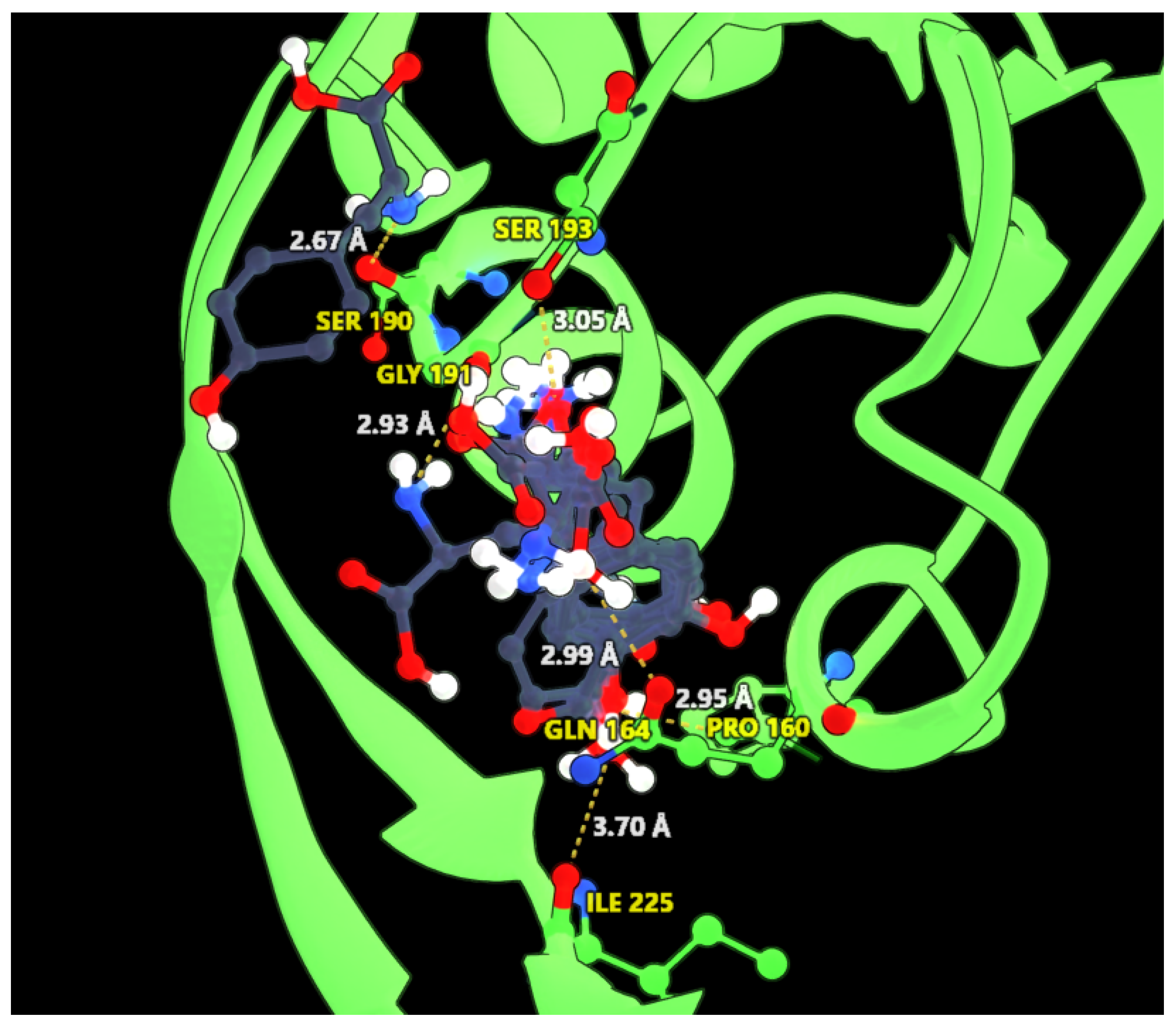
| Original Amino Acid (AroF-I) | Position | Amino Acids Substitutes |
|---|---|---|
| P | 160 | L |
| Q | 164 | A |
| S | 190 | A |
| G | 191 | K |
| S | 193 | A |
| I | 225 | P |
| Strains and Plasmids | Genotype or Markers; Characteristics and Uses | Source or References |
|---|---|---|
| Strains | ||
| Escherichia coli BL21(DE3) | , , , , , [, , , , ]), [ | Novagen |
| Escherichia coli S17.1 | , , , | Simon et al. [48] |
| Pseudomonas putida KT2440 | WT | NBRC100650 |
| Pseudomonas putida | p-coumaric productive strain with aroH under strong constitutive promoter | This study |
| Pseudomonas putida | , | This study |
| Plasmids | ||
| pBBR1MCS-2 | Broad-host-range cloning vector, mobilizable, | Kovach et al. [49] |
| pUC-araC/pBAD | pUC containing the araC/pBAD promoter, | SEVA plasmid collection |
| pBBR1-araC/pBAD | pBBR1MCS-2 containing the araC/pBAD promoter | This study |
| pC2F387 | pBBR1-araC/pBAD with the phhA/B genes cloned downstream of the araC/pBAD promoter | This study |
| pPpu226 | pBBR1-araC/pBAD with the eGFP gene cloned downstream of the araC/pBAD promoter | This study |
| pET28a(+) | Novagen | |
| pET28-AroF-I WT | This study | |
| pET28-AroF-I G191K | This study | |
| pET28-AroF-I P160L | This study | |
| pET28-AroF-I S193A | This study | |
| pET28-AroF-I P160L/G191K | This study | |
| pET28-AroF-I P160L/Q164A | This study | |
| pET28-AroF-I P160L/S190A | This study | |
| pET28-AroF-I P160L/S193A | This study | |
| pK18mobsacB | Widely used gene modifications suicide vector, and for counter-selection | Schäfer et al. [50] |
| pK18-aroHAroF-I P160L/S193A | pK18mobsacB containing the homologous arms of and the P160L/S193A gene to be inserted, | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merre, W.; Andrade, R.; Perot, C.; Chandor-Proust, A.; Ranquet, C. Overproduction of Phenolic Compounds in Pseudomonas putida KT2440 Through Endogen Deregulation of the Shikimate Pathway. BioChem 2025, 5, 4. https://doi.org/10.3390/biochem5010004
Merre W, Andrade R, Perot C, Chandor-Proust A, Ranquet C. Overproduction of Phenolic Compounds in Pseudomonas putida KT2440 Through Endogen Deregulation of the Shikimate Pathway. BioChem. 2025; 5(1):4. https://doi.org/10.3390/biochem5010004
Chicago/Turabian StyleMerre, William, Ricardo Andrade, Cyril Perot, Alexia Chandor-Proust, and Caroline Ranquet. 2025. "Overproduction of Phenolic Compounds in Pseudomonas putida KT2440 Through Endogen Deregulation of the Shikimate Pathway" BioChem 5, no. 1: 4. https://doi.org/10.3390/biochem5010004
APA StyleMerre, W., Andrade, R., Perot, C., Chandor-Proust, A., & Ranquet, C. (2025). Overproduction of Phenolic Compounds in Pseudomonas putida KT2440 Through Endogen Deregulation of the Shikimate Pathway. BioChem, 5(1), 4. https://doi.org/10.3390/biochem5010004