Screening of Non-Conventional Yeasts on Low-Cost Carbon Sources and Valorization of Mizithra Secondary Cheese Whey for Metabolite Production

 ,
, 
 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Media
2.2.1. Lactose-Based Media
2.2.2. Glycerol-Based Media
2.2.3. Glucose-Based Media
2.2.4. Scale-Up, Fed-Batch Cultivation on Secondary Cheese Whey
2.3. Analytical Methods
3. Results
3.1. Growth of C. curvatus on Lactose-Based Media Under Various Nitrogen Conditions
3.2. Growth of C. curvatus on Glycerol-Based Media Under Various Nitrogen Conditions
3.3. Growth of C. curvatus on Glucose-Based Media Under Various Nitrogen Conditions
3.4. Growth of P. laurentii on Lactose-Based Media Under Various Nitrogen Conditions
3.5. Growth of P. laurentii on Glycerol-Based Media Under Various Nitrogen Conditions
3.6. Growth of P. laurentii on Glucose-Based Media Under Various Nitrogen Conditions
3.7. Fed-Batch Bioreactor Cultivation on Mizithra Secondary Cheese Whey and Polysaccharides Secretion by P. laurentii
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SCW | Secondary cheese whey |
| C/N | Carbon-to-nitrogen |
| FAN | Free amino nitrogen |
| FAN0 | Initial free amino nitrogen |
| FANCON | Consumed free amino nitrogen |
| TKN | Total Kjeldahl nitrogen |
| Vw | Working volume |
| X | Dry biomass |
| L | Total lipids |
| FAMEs | Fatty acid methyl esters |
| cPS | Cellular polysaccharide |
| SR | Residual sugar |
| SCON | Consumed sugar |
| MAN | Mannitol |
| YX/S | Substrate-to-biomass conversion coefficient |
| YL/S | Substrate-to-lipid conversion coefficient |
| YcPS/S | Substrate-to-cellular polysaccharide conversion coefficient |
| YMAN/S | Substrate-to-mannitol conversion coefficient |
| KL/X | Lipid content on dry biomass |
| KcPS/X | Cellular polysaccharide content on dry biomass |
| PX | Productivity value for dry biomass |
| PL | Productivity value for lipids |
| PMAN | Productivity value for mannitol |
| PcPS | Productivity value for cellular polysaccharide |
| rS | Substrate’s consumption rate |
References
- Coma, M.; Chatzifragkou, A. Chemicals from Food Supply Chain By-Products and Waste Streams. Molecules 2019, 24, 978. [Google Scholar] [CrossRef]
- Maina, S.; Kachrimanidou, V.; Koutinas, A. A roadmap towards a circular and sustainable bioeconomy through waste valorization. Curr. Opin. Green Sustain. C Hemistry 2017, 8, 18–23. [Google Scholar]
- Papanikolaou, S.; Aggelis, G. Microbial products from wastes and residues. FEMS Microbiol. Lett. 2020, 367, fnaa156. [Google Scholar]
- Di Fidio, N.; Liuzzi, F.; Mastrolitti, S.; Albergo, R.; De Bari, I. Single Cell Oil Production from Undetoxified Arundo donax L. hydrolysate by Cutaneotrichosporon curvatus. J. Microbiol. Biotechnol. 2019, 29, 256–267. [Google Scholar]
- Diamantopoulou, P.; Papanikolaou, S. Biotechnological production of sugar-alcohols: Focus on Yarrowia lipolytica and edible/medicinal mushrooms. Process Biochem. 2023, 124, 113–131. [Google Scholar]
- Eurostat. Milk and Milk Product Statistics. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?%20title=Milk_and_milk_product_statistics#Milk_products (accessed on 16 January 2025).
- Vasilakis, G.; Karayannis, D.; Massouras, T.; Politis, I.; Papanikolaou, S. Biotechnological conversions of Mizithra second cheese whey by wild-type non-conventional yeast strains: Production of yeast cell biomass, single-cell oil and polysaccharides. Appl. Sci. 2022, 12, 11471. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Sakinah, A.M.; Zularisam, A.W.; Pandey, A. Glycerol waste to value added products and its potential applications. Syst. Microbiol. Biomanuf. 2021, 1, 378–396. [Google Scholar]
- Valta, K.; Damala, P.; Panaretou, V.; Orli, E.; Moustakas, K.; Loizidou, M. Review and assessment of waste and wastewater treatment from fruits and vegetables processing industries in Greece. Waste Biomass Valorization 2016, 8, 1629–1648. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Moving Forward on Food Loss and Waste Reduction. In The State of Food and Agriculture 2019; FAO: Rome, Italy, 2019; Available online: https://www.fao.org/3/ca6030en/ca6030en.pdf (accessed on 20 November 2024).
- Vasilakis, G.; Rigos, E.-M.; Giannakis, N.; Diamantopoulou, P.; Papanikolaou, S. Spent mushroom substrate hydrolysis and utilization as potential alternative feedstock for anaerobic co-digestion. Microorganisms 2023, 11, 532. [Google Scholar] [CrossRef]
- Vasilakis, G.; Gamraoui, A.; Karayannis, D.; Giannakis, N.; Chatti, A.; Politis, I.; Diamantopoulou, P.; Papanikolaou, S. Mycelial mass, microbial lipids and γ-linolenic acid (GLA) by Cunninghamella elegans cultivated on agro-industrial residues. Resour. Chem. Mater. 2025, in press. [Google Scholar] [CrossRef]
- Lin, C.S.K.; Pfaltzgraff, L.A.; Herrero-Davila, L.; Mubofu, E.B.; Abderrahim, S.; Clark, J.H.; Koutinas, A.A.; Kopsahelis, N.; Stamatelatou, K.; Dickson, F.; et al. Food waste as a valuable resource for the production of chemicals, materials and fuels. Current situation and global perspective. Energy Environ. Sci. 2013, 6, 426. [Google Scholar]
- Ladakis, D.; Papapostolou, H.; Vlysidis, A.; Koutinas, A. Inventory of food processing side streams in European Union and prospects for biorefinery development. In Food Industry Wastes; Academic Press: Cambridge, MA, USA, 2020; pp. 181–199. [Google Scholar]
- Haq, I.; Qaisar, K.; Nawaz, A.; Akram, F.; Mukhtar, H.; Zohu, X.; Xu, Y.; Mumtaz, M.W.; Rashid, U.; Ghani, W.A.W.A.K.; et al. Advances in valorization of lignocellulosic biomass towards energy generation. Catalysts 2021, 11, 309. [Google Scholar] [CrossRef]
- Pires, A.F.; Marnotes, N.G.; Rubio, O.D.; Garcia, A.C.; Pereira, C.D. Dairy by-products: A review on the valorization of whey and second cheese whey. Foods 2021, 10, 1067. [Google Scholar] [CrossRef]
- Asopa, R.P.; Bhoi, R.; Saharan, V.K. Valorization of glycerol into value-added products: A comprehensive review on biochemical route. Bioresour. Technol. Rep. 2022, 20, 101290. [Google Scholar]
- Rywinska, A.; Rymowicz, W.; Marcinkiewicz, M. Valorization of raw glycerol for citric acid production by Yarrowia lipolytica yeast. Electron. J. Biotechnol. 2010, 13, 1–9. [Google Scholar]
- Carota, E.; Crognale, S.; D’Annibale, A.; Gallo, A.M.; Stazi, S.R.; Petruccioli, M. A sustainable use of Ricotta Cheese Whey for microbial biodiesel production. Sci. Total Environ. 2017, 584–585, 554–560. [Google Scholar]
- Kothri, M.; Mavrommati, M.; Elazzazy, A.M.; Baeshen, M.N.; Moussa, T.A.A.; Aggelis, G. Microbial sources of polyunsaturated fatty acids (PUFAs) and the prospect of organic residues and wastes as growth media for PUFA-producing microorganisms. FEMS Microbiol. Lett. 2020, 367, fnaa028. [Google Scholar]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic activities of biotechnological interest in Yarrowia lipolytica grown on glycerol in repeated batch cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar]
- Karayannis, D.; Papanikolaou, S.; Vatistas, C.; Paris, C.; Chevalot, I. Yeast lipid produced through glycerol conversions and its use for enzymatic synthesis of amino acid-based biosurfactants. Int. J. Mol. Sci. 2023, 24, 714. [Google Scholar]
- Giannakis, N.; Carmona-Cabello, M.; Makri, A.; Leiva-Candia, D.; Filippi, K.; Argeiti, C.; Pateraki, C.; Dorado, M.P.; Koutinas, A.; Stylianou, E. Spent coffee grounds and orange peel residues based biorefinery for microbial oil and biodiesel conversion estimation. Renew. Energy 2023, 209, 382–392. [Google Scholar]
- Gamraoui, A.; Vasilakis, G.; El Ouaer, M.; Karayannis, D.; Papanikolaou, S.; Landoulsi, A.; Chatti, A. Innovative combined method for tuna wastewater treatment using Yarrowia lipolytica, TiO2 nanoparticles and sounds. Waste Biomass Valorization 2025, in press. [Google Scholar] [CrossRef]
- Koukoumaki, D.I.; Papanikolaou, S.; Ioannou, Z.; Gkatzionis, K.; Sarris, D. The development of novel edible films from single-cell protein produced by the biotechnological valorization of cheese whey. Appl. Microbiol. 2024, 4, 1030–1041. [Google Scholar] [CrossRef]
- Hofmeyer, T.; Hackenschmidt, S.; Nadler, F.; Thürmer, A.; Daniel, R.; Kabisch, J. Draft genome sequence of Cutaneotrichosporon curvatus DSM 101032 (formerly Cryptococcus curvatus), an oleaginous yeast producing polyunsaturated fatty acids. Genome Announc. 2016, 4, e00362-16. [Google Scholar] [PubMed]
- Shoff, C.J.; Perfect, J.R. Uncommon yeasts and molds causing human disease. Ref. Modul. Life Sci. 2021, 1, 813–834. [Google Scholar]
- Chattopadhyay, A.; Maiti, M.K. Lipid production by oleaginous yeasts. Adv. Appl. Microbiol. 2021, 116, 1–98. [Google Scholar]
- Di Fidio, N.; Minonne, F.; Antonetti, C.; Raspolli Galletti, A.M. Cutaneotrichosporon oleaginosus: A versatile whole-cell biocatalyst for the production of single-cell oil from agro-industrial wastes. Catalysts 2021, 11, 1291. [Google Scholar] [CrossRef]
- Pavlova, K.; Rusinova-Videva, S.; Kuncheva, M.; Kratchanova, M.; Gocheva, M.; Dimitrova, S. Synthesis and characterization of an exopolysaccharide by antarctic yeast strain Cryptococcus laurentii AL100. Appl. Biochem. Biotechnol. 2011, 163, 1038–1052. [Google Scholar]
- Smirnou, D.; Hrubošová, D.; Kulhánek, J.; Švík, K.; Bobková, L.; Moravcová, V.; Krčmář, M.; Franke, L.; Velebný, V. Cryptococcus laurentii extracellular biopolymer production for application in wound management. Appl. Biochem. Biotechnol. 2014, 174, 1344–1353. [Google Scholar]
- De Almeida, E.L.M.; Ventorim, R.Z.; de Moura Ferreira, M.A.; da Silveira, W.B. Papiliotrema laurentii: General features and biotechnological applications. Appl. Microbiol. Biotechnol. 2022, 106, 6963–6976. [Google Scholar]
- Nigam, P.; Singh, D. Production of Secondary Metabolites—Fungi. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 1999; pp. 1319–1328. [Google Scholar]
- Bellou, S.; Baeshen, M.N.; Elazzazy, A.M.; Aggeli, D.; Sayegh, F.; Aggelis, G. Microalgal lipids biochemistry and biotechnological perspectives. Biotechnol. Adv. 2014, 32, 1476–1493. [Google Scholar]
- Fouillaud, M.; Dufossé, L. Microbial Secondary Metabolism and Biotechnology. Microorganisms 2022, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Aggelis, G.; Marc, I. Kinetic profile of the cellular lipid composition in an oleaginous Yarrowia lipolytica capable of producing a cocoa-butter substitute from industrial fats. Antonie Van Leeuwenhoek 2001, 80, 215–224. [Google Scholar]
- Boshagh, F.; Rostami, K.; van Niel, E.W.J. Application of kinetic models in dark fermentative hydrogen production—A critical review. Int. J. Hydrogen Energy 2022, 47, 21952–21968. [Google Scholar]
- Seiboth, B.; Pakdaman, B.S.; Hartl, L.; Kubicek, C.P. Lactose metabolism in filamentous fungi: How to deal with an unknown substrate. Fungal Biol. Rev. 2007, 21, 42–48. [Google Scholar]
- Evans, C.T.; Ratledge, C. The physiological significance of citric acid in the control of metabolism in lipid-accumulating yeasts. Biotechnol. Genet. Eng. Rev. 1985, 3, 349–375. [Google Scholar]
- Bellou, S.; Triantaphyllidou, I.E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial oils as food additives: Recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [PubMed]
- Caporusso, A.; De Bari, I.; Valerio, V.; Albergo, R.; Liuzzi, F. Conversion of cardoon crop residues into single cell oils by Lipomyces tetrasporus and Cutaneotrichosporon curvatus: Process optimizations to overcome the microbial inhibition of lignocellulosic hydrolysates. Ind. Crops Prod. 2021, 159, 113030. [Google Scholar]
- Broos, W.; Wittner, N.; Geerts, J.; Dries, J.; Vlaeminck, S.E.; Gunde-Cimerman, N.; Richel, A.; Cornet, I. Evaluation of lignocellulosic wastewater valorization with the oleaginous yeasts R. kratochvilovae EXF7516 and C. oleaginosum ATCC 20509. Fermentation 2022, 8, 204. [Google Scholar] [CrossRef]
- Gill, I.; Valivety, R. Polyunsaturated fatty acids, part 1: Occurrence, biological activities and applications. Trends Biotechnol. 1997, 15, 401–409. [Google Scholar]
- Bellou, S.; Makri, A.; Sarris, D.; Michos, K.; Rentoumi, P.; Celik, A.; Papanikolaou, S.; Aggelis, G. The olive mill wastewater as substrate for single cell oil production by Zygomycetes. J. Biotechnol. 2014, 170, 50–59. [Google Scholar]
- Vasilakis, G.; Roidouli, C.; Karayannis, D.; Giannakis, N.; Rondags, E.; Chevalot, I.; Papanikolaou, S. Study of different parameters affecting production and productivity of polyunsaturated fatty acids (PUFAs) and γ-linolenic acid (GLA) by Cunninghamella elegans through glycerol conversion in shake flasks and bioreactors. Microorganisms 2024, 12, 2097. [Google Scholar] [CrossRef] [PubMed]
- Gallego-García, M.; Susmozas, A.; Moreno, A.D.; Negro, M.J. Evaluation and Identification of Key Economic Bottlenecks for Cost-Effective Microbial Oil Production from Fruit and Vegetable Residues. Fermentation 2022, 8, 334. [Google Scholar] [CrossRef]
- Gonçalves, C.; Ferreira, C.; Gonçalves, L.G.; Turner, D.L.; Leandro, M.J.; Salema-Oom, M.; Santos, H.; Gonçalves, P. A new pathway for mannitol metabolism in yeasts suggests a link to the evolution of alcoholic fermentation. Front. Microbiol. 2019, 10, 2510. [Google Scholar]
- Klein, M.; Swinnen, S.; Thevelein, J.M.; Nevoigt, E. Glycerol metabolism and transport in yeast and fungi: Established knowledge and ambiguities. Environ. Microbiol. 2017, 19, 878–893. [Google Scholar] [PubMed]
- Vieira, N.M.; dos Santos, R.C.V.; Germano, V.K.C.; Ventorim, R.Z.; de Almeida, E.L.M.; da Silveira, F.A.; Ribeiro, J.I., Jr.; da Silveira, W.B. Isolation of a new Papiliotrema laurentii strain that displays capacity to achieve high lipid content from xylose. 3 Biotech 2020, 10, 382. [Google Scholar]
- Polburee, P.; Yongmanitchai, W.; Lertwattanasakul, N.; Ohashi, T.; Fujiyama, K.; Limtong, S. Characterization of oleaginous yeasts accumulating high levels of lipid when cultivated in glycerol and their potential for lipid production from biodiesel-derived crude glycerol. Fungal Biol. 2015, 119, 1194–1204. [Google Scholar]
- Castanha, R.F.; Mariano, A.P.; de Morais, L.A.; Scramin, S.; Monteiro, R.T. Optimization of lipids production by Cryptococcus laurentii 11 using cheese whey with molasses. Braz. J. Microbiol. 2014, 45, 379–387. [Google Scholar]
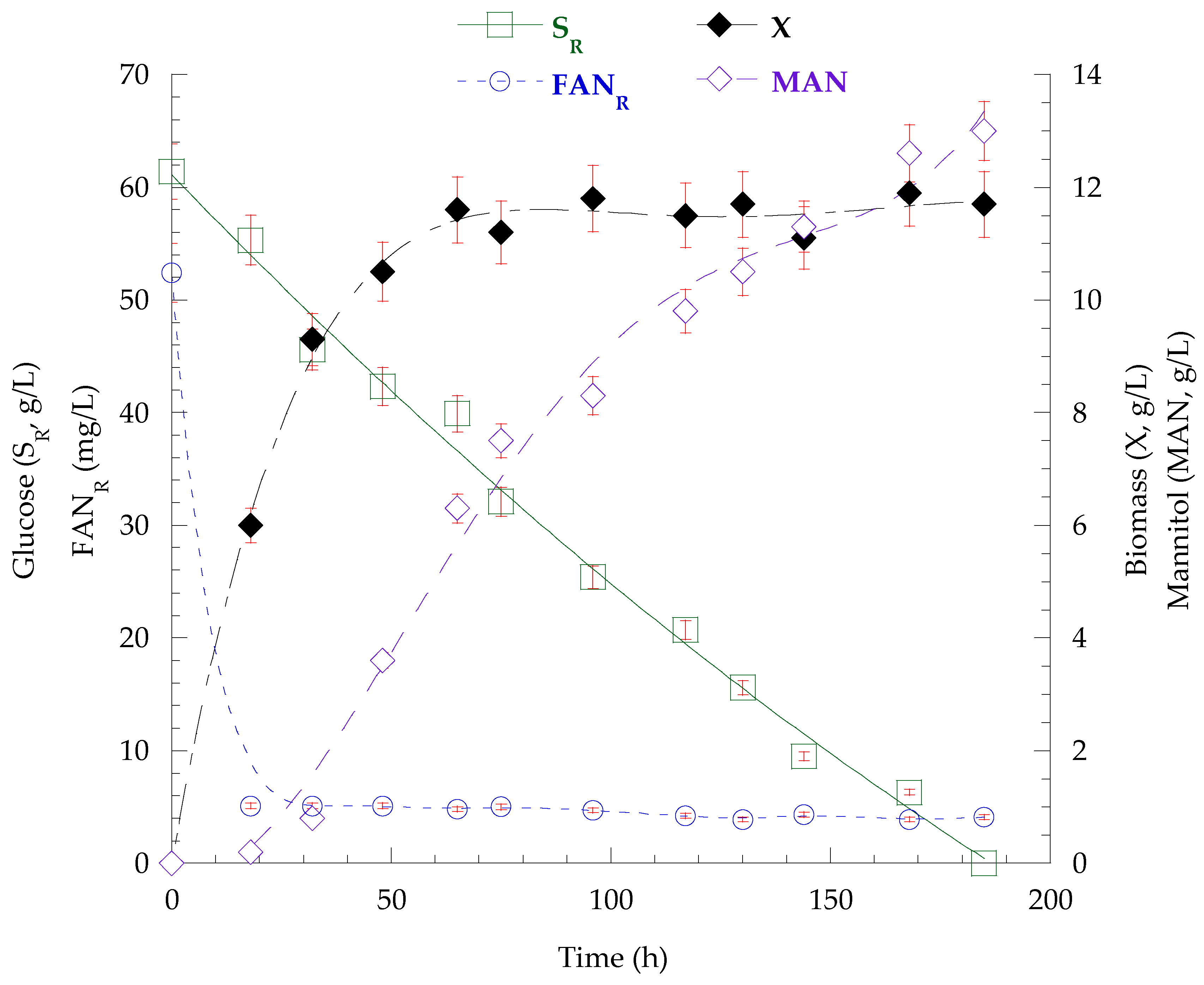
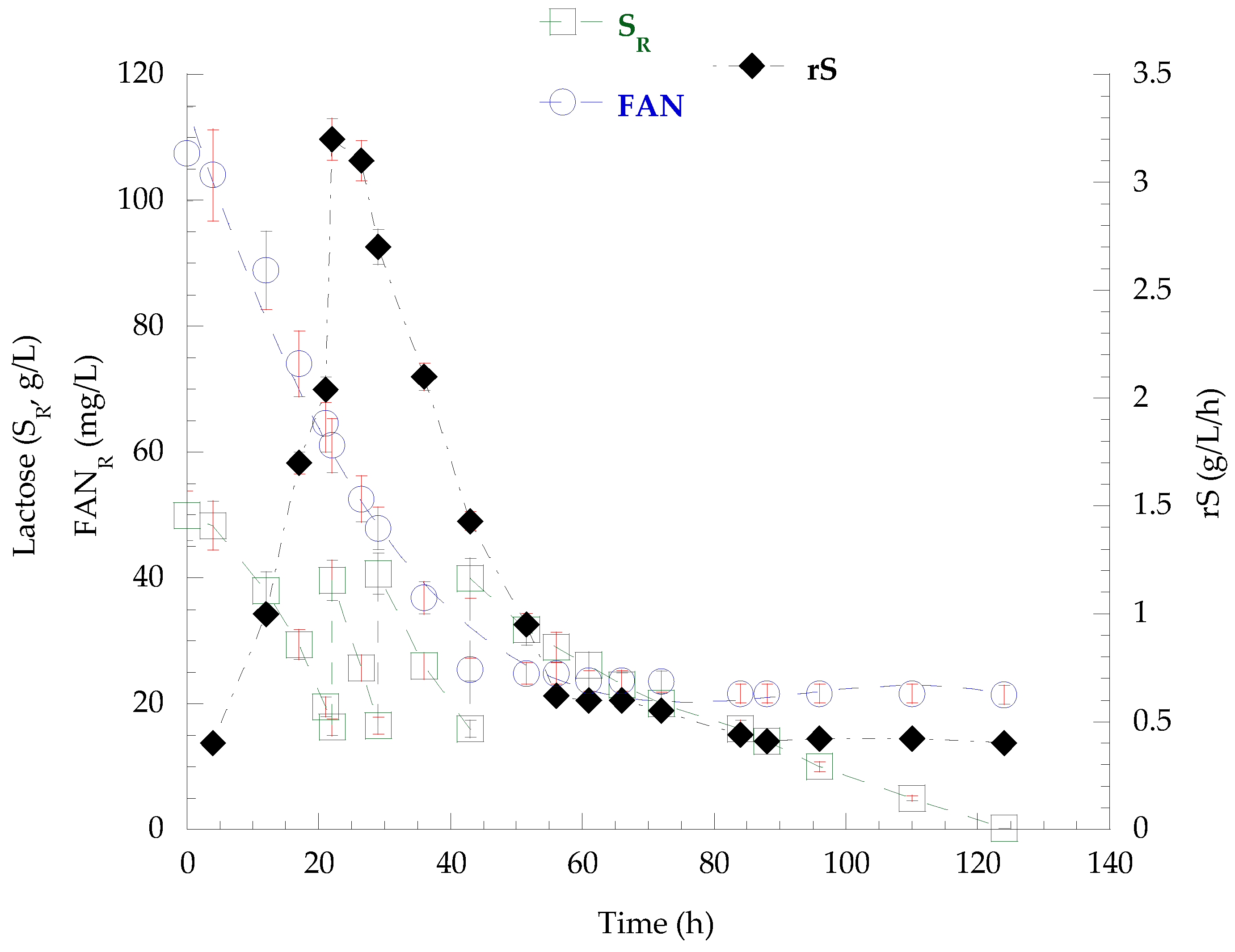
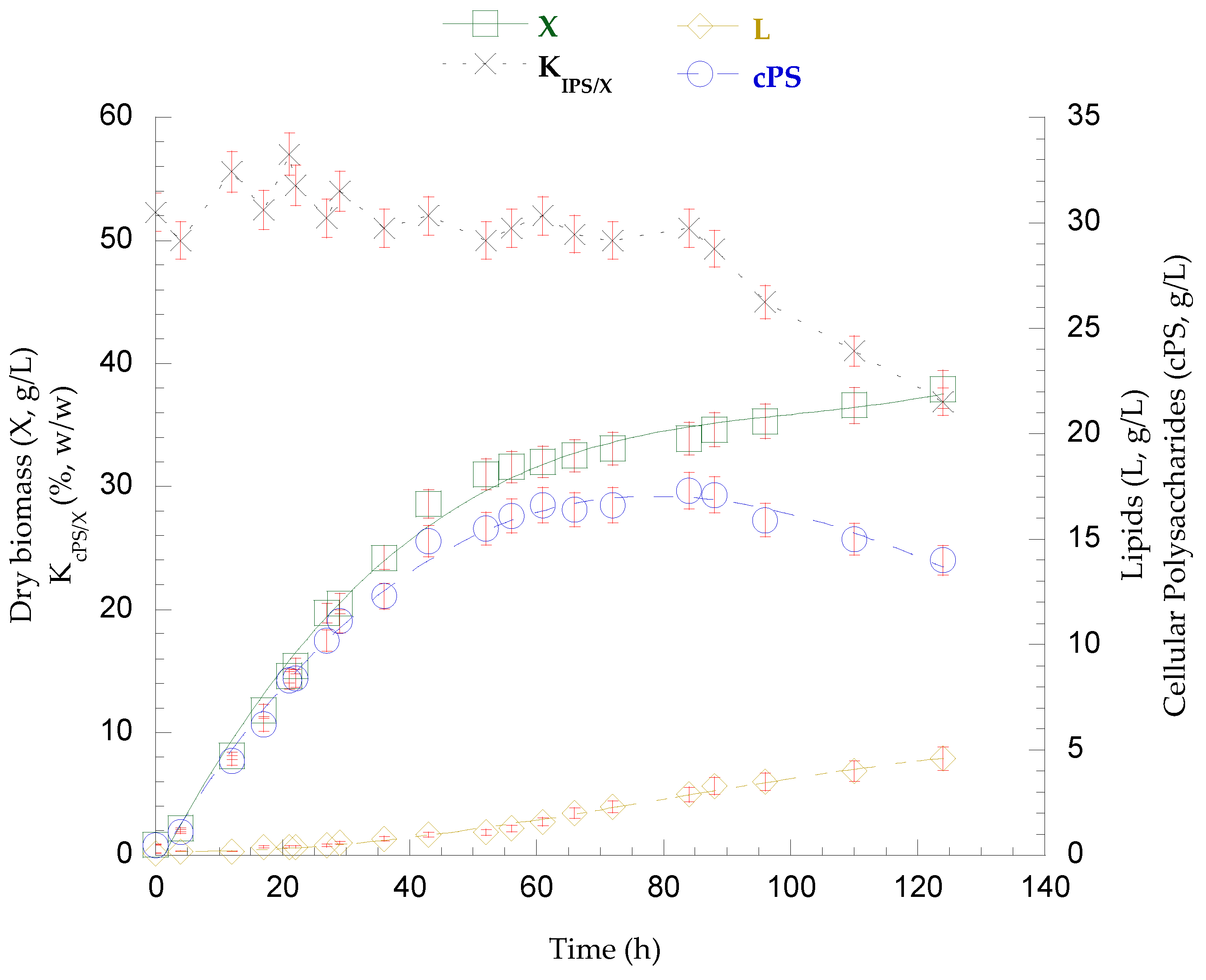

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar (g/L) | Lactose | Glycerol | Glucose | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 60 ± 2 | |||||||||
| C/N ratio (mol/mol) | 20 | 80 | 160 | 20 | 80 | 160 | 20 | 80 | 160 |
| Yeast Extract (g/L) | 7.4 | 1.8 | 0.9 | 6.8 | 1.7 | 0.9 | 7.0 | 1.7 | 0.9 |
| (NH4)2SO4 (g/L) | 3.5 | 0.9 | 0.4 | 3.2 | 0.8 | 0.4 | 3.3 | 0.8 | 0.4 |
| KH2PO4 | 7.00 | ||||||||
| Na2HPO4 | 2.50 | ||||||||
| MgSO4 × 7H2O | 1.50 | ||||||||
| MnSO4 × H2O | 0.06 | ||||||||
| ZnSO4 × 7H2O | 0.02 | ||||||||
| CaCl2 × 2H2O | 0.15 | ||||||||
| FeCl3 × 6H2O | 0.15 | ||||||||
| Total Solids (g/L) | Lactose (g/L) | FAN0 (mg/L) | TKN (g/L) | pH |
|---|---|---|---|---|
| 69.6 ± 1.2 | 56.5 ± 0.4 | 107.4 ± 3.1 | 1.1± 0.1 | 6.2 ± 0.2 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PL (mg/L/h) | PcPS (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 142 | 60.4 a ± 0.1 | 27.8 a ± 0.3 | 1.1 a ± 0.1 | 12.6 a* ± 0.2 | 0.46 a ± 0.01 | 0.02 a ± 0.00 | 0.21 a ± 0.00 | 0.039 a ± 0.001 | 0.455 a ± 0.010 | 195.8 a ± 2.1 | 7.7 a ± 0.8 | 89.1 a ± 1.0 | 416 a ± 16 |
| 80 | 142 | 33.2 b ± 0.6 | 13.4 b ± 0.2 | 1.4 b ± 0.0 | 6.9 b ± 0.1 | 0.40 a ± 0.02 | 0.04 b ± 0.00 | 0.21 ab ± 0.01 | 0.105 b ± 0.001 | 0.516 b* ± 0.014 | 94.4 b ± 1.3 | 9.9 b ± 0.0 | 48.7 b ± 0.6 | 93 b ± 4 |
| 160 | 164 | 17.1 c ± 0.3 | 7.9 c ± 0.3 | 1.8 c ± 0.2 | 3.1 c ± 0.0 | 0.46a ± 0.03 | 0.10 c ± 0.02 | 0.18 b ± 0.01 | 0.229 c* ± 0.034 | 0.395 c ± 0.013 | 48.2 c ± 1.8 | 11.0 b ± 1.2 | 18.9 c ± 0.0 | 56 c ± 1 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 142 | 29.5 a ± 1.4 | 11.5 a ± 0.3 | 42.0 a ± 1.4 | 17.0 a ± 0.7 | 41.0 a ± 1.8 | 59.0 a ± 2.0 |
| 80 | 142 | 27.3 a ± 1.1 | 10.9 a ± 0.3 | 47.2 b ± 1.0 | 14.6 b ± 0.2 | 39.1 a ± 1.5 | 60.9 a ± 1.2 |
| 160 | 164 | 27.4 a ± 1.2 | 11.3 a ± 0.1 | 51.3 c ± 1.4 | 10.0 c ± 0.1 | 31.5 b ± 1.3 | 68.5 b ± 1.5 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | Man (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | YMan/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PcPS (mg/L/h) | PMan (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 171 | 58.2 a ± 0.1 | 31.3 a* ± 0.4 | 0.9 a ± 0.1 | 12.6 a* ± 0.3 | 0.0 a ± 0.0 | 0.54 a* ± 0.01 | 0.02 a ± 0.00 | 0.22 a ± 0.01 | 0.00 a ± 0.00 | 0.029 a ± 0.003 | 0.401 a ± 0.016 | 183.0 a ± 2.3 | 73.4 a ± 2.0 | 0.0 a ± 0.0 | 438 a ± 16 |
| 80 | 331 | 59.1 a ± 0.3 | 15.3 0 b ± 0.2 | 2.1 b ± 0.1 | 7.0 b ± 0.1 | 5.9 b ± 0.3 | 0.26 b ± 0.00 | 0.04 b ± 0.00 | 0.12 b ± 0.00 | 0.10 b ± 0.01 | 0.137 b ± 0.009 | 0.459 b ± 0.011 | 46.2 b ± 0.6 | 21.2 b ± 0.3 | 17.8 b ± 0.9 | 95 b ± 4 |
| 160 | 450 | 45.6 b ± 0.4 | 9.4 c ± 0.1 | 1.4 c ± 0.0 | 4.6 c ± 0.0 | 6.2 b ± 0.2 | 0.21 c ± 0.00 | 0.03 ab ± 0.00 | 0.10 b ± 0.00 | 0.14 c ± 0.00 | 0.147 b ± 0.004 | 0.493 c ± 0.002 | 21.0 c ± 0.1 | 10.3 c ± 0.0 | 13.9 c ± 0.3 | 59 c ± 1 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 171 | 23.4 a ± 1.0 | 18.3 a ± 1.2 | 39.9 a ± 1.6 | 18.4 a ± 1.1 | 41.7 a ± 2.2 | 58.3 a ± 2.8 |
| 80 | 331 | 24.1 a ± 0.9 | 13.5 b ± 0.3 | 49.8 b ± 1.0 | 12.6 b ± 0.2 | 37.6 ab ± 1.3 | 62.4 ab ± 1.2 |
| 160 | 450 | 23.2 a ± 0.4 | 13.6 b ± 0.1 | 52.5 b ± 1.2 | 9.6 c ± 0.1 | 36.8 b ± 0.5 | 63.2 b ± 1.4 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | Man (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | YMan/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PcPS (mg/L/h) | PMan (mg/L/h) | FANCON (mg/L) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | i | 62 | 61.3 ± 0.2 | 21.7 ± 0.2 | 0.9 ± 0.0 | 7.5 ± 0.1 | 7.7 ± 0.2 | 0.35 ± 0.01 | 0.02 ± 0.00 | 0.12 ± 0.01 | 0.13 ± 0.00 | 0.042 ± 0.001 | 0.346 ± 0.008 | 350.1 ± 3.1 | 121.0 ± 1.6 | 124.2 ± 3.2 | 316 ± 4 |
| ii | 117 | 61.3 a ± 0.2 | 25.0 a ± 0.1 | 1.1 a ± 0.0 | 11.4 a ± 0.2 | 0.0 a ± 0.0 | 0.41 a ± 0.00 | 0.02 a ± 0.00 | 0.19 a ± 0.01 | 0.00 a ± 0.00 | 0.043 a ± 0.001 | 0.454 a ± 0.009 | 213.7 a* ± 0.8 | 97.3 a* ± 1.9 | 0.0 a ± 0.0 | 373 a ± 11 | |
| 80 | 129 | 61.9 a ± 0.3 | 15.2 b ± 0.2 | 1.8 b ± 0.1 | 6.8 b ± 0.1 | 8.3 b ± 0.2 | 0.25 b ± 0.00 | 0.03 ab ± 0.00 | 0.11 b ± 0.00 | 0.13 b ± 0.01 | 0.118 b ± 0.008 | 0.446 a ± 0.014 | 117.8 b ± 1.6 | 52.7 b ± 0.8 | 64.3 b ± 1.6 | 83 b ± 7 | |
| 160 | 185 | 61.4 a ± 0.2 | 13.5 c ± 0.2 | 2.2 b ± 0.2 | 6.2 b ± 0.2 | 13.0 c* ± 0.4 | 0.22 b ± 0.01 | 0.04 b ± 0.01 | 0.10 b ± 0.01 | 0.21 c ± 0.02 | 0.163 c ± 0.017 | 0.459 a ± 0.022 | 73.0 c ± 1.1 | 33.5 c ± 1.1 | 70.3 b* ± 2.1 | 49 c ± 3 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 117 | 18.6 a ± 0.4 | 10.5 a ± 1.1 | 47.3 a ± 1.2 | 23.6 a ± 0.1 | 29.1 a ± 1.4 | 70.9 a ± 1.3 |
| 80 | 129 | 22.1 b ± 0.9 | 13.1 b ± 0.5 | 50.6 ab ± 0.6 | 14.2 b ± 0.4 | 35.2 b ± 1.5 | 64.8 b ± 1.1 |
| 160 | 185 | 24.6 c ± 0.3 | 13.2 b ± 0.2 | 52.8 b ± 1.1 | 9.4 c ± 0.1 | 37.8 b ± 0.5 | 62.2 b ± 1.2 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PL (mg/L/h) | PcPS (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 81 a | 58.7 ± 0.2 | 24.7 a* ± 0.2 | 1.0 a ± 0.1 | 10.1 a ± 0.2 | 0.42 a ± 0.01 | 0.02 a ± 0.00 | 0.17 a ± 0.00 | 0.042 a ± 0.003 | 0.410 a ± 0.010 | 304.4 a* ± 3.0 | 12.7 a ± 0.9 | 124.7 a* ± 2.5 | 374 a ± 11 |
| 80 | 135 b | 59.4 0.3 | 17.8 b ± 0.3 | 1.4 a ± 0.1 | 7.6 b ± 0.2 | 0.30 b ± 0.01 | 0.02 ab ± 0.01 | 0.13 b ± 0.01 | 0.079 b ± 0.007 | 0.427 a ± 0.019 | 131.9 b ± 2.2 | 10.4 a ± 0.7 | 56.3 b ± 1.5 | 89 b ± 6 |
| 160 | 219 c | 58.9 ± 0.1 | 13.1 c ± 0.2 | 2.2 b ± 0.2 | 5.2 c ± 0.1 | 0.22 c ± 0.01 | 0.04 b ± 0.01 | 0.09 c ± 0.00 | 0.168 c ± 0.018 | 0.397 a ± 0.014 | 59.8 c ± 0.9 | 10.1 a ± 0.9 | 23.7 c ± 0.4 | 48 c ± 5 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 81 | 29.0 a ± 0.9 | 18.3 a ± 0.7 | 40.7 a ± 0.6 | 12.0 a ± 0.2 | 47.3 a ± 1.6 | 52.7 a ± 0.7 |
| 80 | 135 | 29.6 a ± 0.7 | 18.1 a ± 1.0 | 41.4 ab ± 1.0 | 10.9 b ± 0.4 | 47.7 a ± 1.7 | 52.3 a ± 2.4 |
| 160 | 219 | 32.8 b ± 0.7 | 13.9 b ± 1.0 | 44.0 b ± 1.2 | 9.3 c ± 0.4 | 46.7 a ± 1.6 | 53.3 a ± 1.7 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PL (mg/L/h) | PcPS (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 340 | 59.4 a ± 0.2 | 23.5 a ± 0.4 | 1.2 a ± 0.1 | 11.1 a ± 0.2 | 0.40 a ± 0.00 | 0.02 a ± 0.00 | 0.19 a ± 0.00 | 0.051 a ± 0.005 | 0.474 a ± 0.015 | 69.1 a ± 1.2 | 3.5 a ± 0.3 | 32.8 a ± 0.4 | 344 a ± 20 |
| 80 | 440 | 39.7 b ± 0.6 | 13.9 b ± 0.5 | 1.3 a ± 0.1 | 6.2 b ± 0.2 | 0.35 b ± 0.02 | 0.03 a ± 0.01 | 0.1 b ± 0.00 | 0.094 b ± 0.010 | 0.447 ab ± 0.031 | 31.5 b ± 1.2 | 3.0 a ± 0.2 | 14.1 b ± 0.5 | 91 b ± 7 |
| 160 | 440 | 31.2 c ± 0.4 | 8.3 c ± 0.2 | 2.1 b ± 0.1 | 3.5 c ± 0.1 | 0.27 c ± 0.01 | 0.07 b ± 0.00 | 0.11 c ± 0.01 | 0.251 c* ± 0.021 | 0.419 b ± 0.025 | 19.0 c ± 0.3 | 4.8 b ± 0.2 | 8.0 c ± 0.2 | 50 c ± 4 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 340 | 24.3 a ± 0.9 | 17.2 a ± 0.7 | 41.6 a ± 1.1 | 17.0 a ± 0.2 | 41.5 a ± 1.6 | 58.5 a ± 1.4 |
| 80 | 440 | 23.6 a ± 0.7 | 16.6 a ± 0.5 | 40.0 a ± 1.0 | 19.8 b ± 0.8 | 40.2 a ± 1.3 | 59.8 a ± 1.8 |
| 160 | 440 | 25.0 a ± 1.1 | 16.1 a ± 0.6 | 40.7 a ± 0.5 | 18.2 ab ± 0.4 | 41.1 a ± 1.6 | 58.9 a ± 0.9 |
| C/N (mol/mol) | Time (h) | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PL (mg/L/h) | PcPS (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 20 | 91 a | 61.4 ± 0.2 | 21.4 a ± 0.4 | 1.1 a ± 0.0 | 10.5 a ± 0.3 | 0.35 a ± 0.01 | 0.02 a ± 0.00 | 0.17 a ± 0.01 | 0.051 a ± 0.000 | 0.491 a ± 0.014 | 235.2 a ± 4.4 | 12.1 a ± 0.0 | 115.4 a ± 3.3 | 326 a ± 13 |
| 80 | 125 b | 59.0 ± 0.1 | 16.5 b ± 0.3 | 1.7 b ± 0.2 | 7.6 b ± 0.1 | 0.28 b ± 0.01 | 0.03 ab ± 0.00 | 0.13 b ± 0.00 | 0.103 b ± 0.014 | 0.461 a ± 0.021 | 132.0 b ± 2.4 | 13.6 a ± 1.6 | 60.8 b ± 0.8 | 85 b ± 3 |
| 160 | 188 c | 58.7 ± 0.3 | 11.9 c ± 0.1 | 2.1 b ± 0.0 | 4.9 c ± 0.1 | 0.20 c ± 0.01 | 0.04 b ± 0.00 | 0.08 c ± 0.01 | 0.176 c ± 0.002 | 0.412 b ± 0.013 | 63.3 c ± 0.5 | 11.2 a ± 0.0 | 26.1 c ± 0.5 | 50 c ± 4 |
| C/N (mol/mol) | Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | ||
| 20 | 91 | 32.1 a ± 1.1 | 12.6 a ± 0.5 | 42.2 a ± 1.7 | 13.1 a ± 0.1 | 44.7 a ± 1.7 | 55.3 a ± 1.8 |
| 80 | 125 | 28.7 b ± 1.1 | 18.0 b ± 0.7 | 42.5 a ± 0.5 | 10.8 b ± 0.4 | 46.7 a ± 1.8 | 53.3 a ± 1.0 |
| 160 | 188 | 29.0 b ± 0.6 | 17.1 b ± 0.3 | 44.8 a ± 1.0 | 9.1 c ± 0.6 | 46.1 a ± 0.8 | 53.9 a ± 1.6 |
| Time (h) | pH | Scons (g/L) | X (g/L) | L (g/L) | cPS (g/L) | YX/S (g/g) | YL/S (g/g) | YcPS/S (g/g) | KL/X (g/g) | KcPS/X (g/g) | PX (mg/L/h) | PL (mg/L/h) | PcPS (mg/L/h) | FANCON (mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 21 | 6.1 a | 30.4 a ± 1.1 | 14.6 a ± 0.5 | 0.4 a ± 0.2 | 8.3 a ± 0.3 | 0.4 8 a ± 0.05 | 0.01 a ± 0.01 | 0.27 a ± 0.02 | 0.032 a ± 0.011 | 0.570 a ± 0.040 | 696.2 a ± 22.8 | 20.9 a ± 7.6 | 396.8 a ± 12.7 | 49 a ± 4 |
| 56 | 6.5 b | 92.4 b ± 0.9 | 31.6 b ± 0.4 | 1.3 b ± 0.1 | 16.1 b ± 0.2 | 0.34 b ± 0.01 | 0.01 a ± 0.01 | 0.17 b ± 0.01 | 0.040 a ± 0.005 | 0.510 a ± 0.012 | 564.3 b ± 7.1 | 22.6 a ± 2.4 | 287.8 b ± 3.3 | 82 b ± 7 |
| 88 | 7.3 c | 107.4 c ± 1.3 | 34.6 c ± 0.6 | 3.3 c ± 0.2 | 17.1 b ± 0.4 | 0.32 b ± 0.01 | 0.03 b ± 0.00 | 0.16 b ± 0.01 | 0.095 b ± 0.008 | 0.493 a ± 0.022 | 393.2 c ± 6.8 | 37.2 b ± 2.6 | 193.8 c ± 5.1 | 86 b ± 2 |
| 124 | 7.3 c | 121.2 d ± 0.1 | 37.9 d ± 0.5 | 4.6 d ± 0.2 | 14.0 c ± 0.3 | 0.31 b ± 0.01 | 0.04 b ± 0.00 | 0.12 c ± 0.00 | 0.121 c ± 0.002 | 0.369 b ± 0.013 | 305.9 d ± 3.8 | 36.7 b ± 2.0 | 112.9 d ± 2.4 | 86 b ± 3 |
| Time (h) | g/100 g of Total FA | |||||
|---|---|---|---|---|---|---|
| C16:0 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | SFA | UFA | |
| 188 | 35.6 ± 1.4 | 19.0 ± 0.8 | 36.1 ± 1.0 | 9.4 ± 0.3 | 54.6 ± 2.3 | 45.5 ± 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilakis, G.; Tefa, R.; Georgoulakis, A.; Karayannis, D.; Politis, I.; Papanikolaou, S. Screening of Non-Conventional Yeasts on Low-Cost Carbon Sources and Valorization of Mizithra Secondary Cheese Whey for Metabolite Production. BioTech 2025, 14, 24. https://doi.org/10.3390/biotech14020024
Vasilakis G, Tefa R, Georgoulakis A, Karayannis D, Politis I, Papanikolaou S. Screening of Non-Conventional Yeasts on Low-Cost Carbon Sources and Valorization of Mizithra Secondary Cheese Whey for Metabolite Production. BioTech. 2025; 14(2):24. https://doi.org/10.3390/biotech14020024
Chicago/Turabian StyleVasilakis, Gabriel, Rezart Tefa, Antonios Georgoulakis, Dimitris Karayannis, Ioannis Politis, and Seraphim Papanikolaou. 2025. "Screening of Non-Conventional Yeasts on Low-Cost Carbon Sources and Valorization of Mizithra Secondary Cheese Whey for Metabolite Production" BioTech 14, no. 2: 24. https://doi.org/10.3390/biotech14020024
APA StyleVasilakis, G., Tefa, R., Georgoulakis, A., Karayannis, D., Politis, I., & Papanikolaou, S. (2025). Screening of Non-Conventional Yeasts on Low-Cost Carbon Sources and Valorization of Mizithra Secondary Cheese Whey for Metabolite Production. BioTech, 14(2), 24. https://doi.org/10.3390/biotech14020024