
Relative Water Economy Is a Useful Index of Aridity Tolerance for Australian Poephiline Finches


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
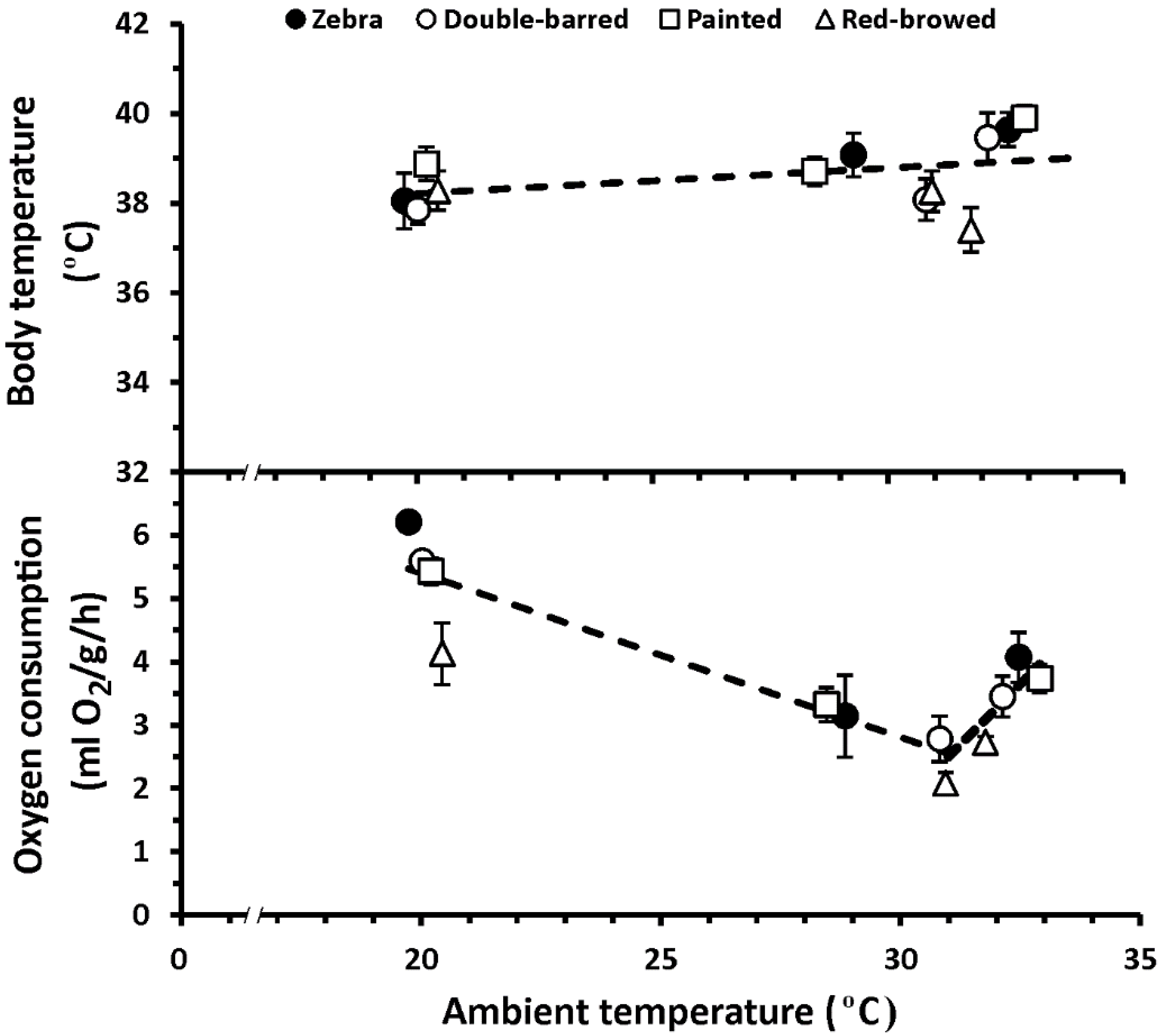
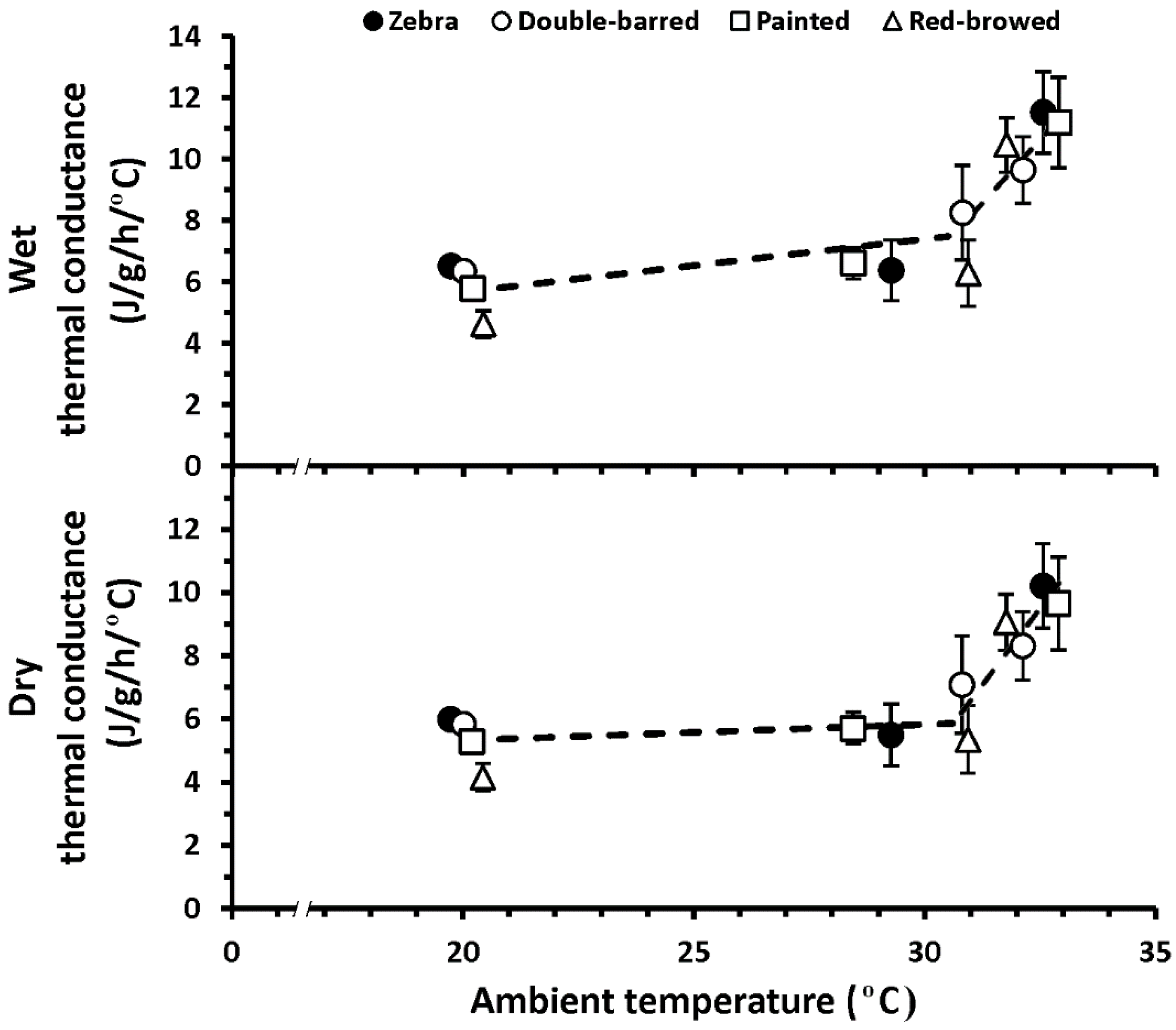
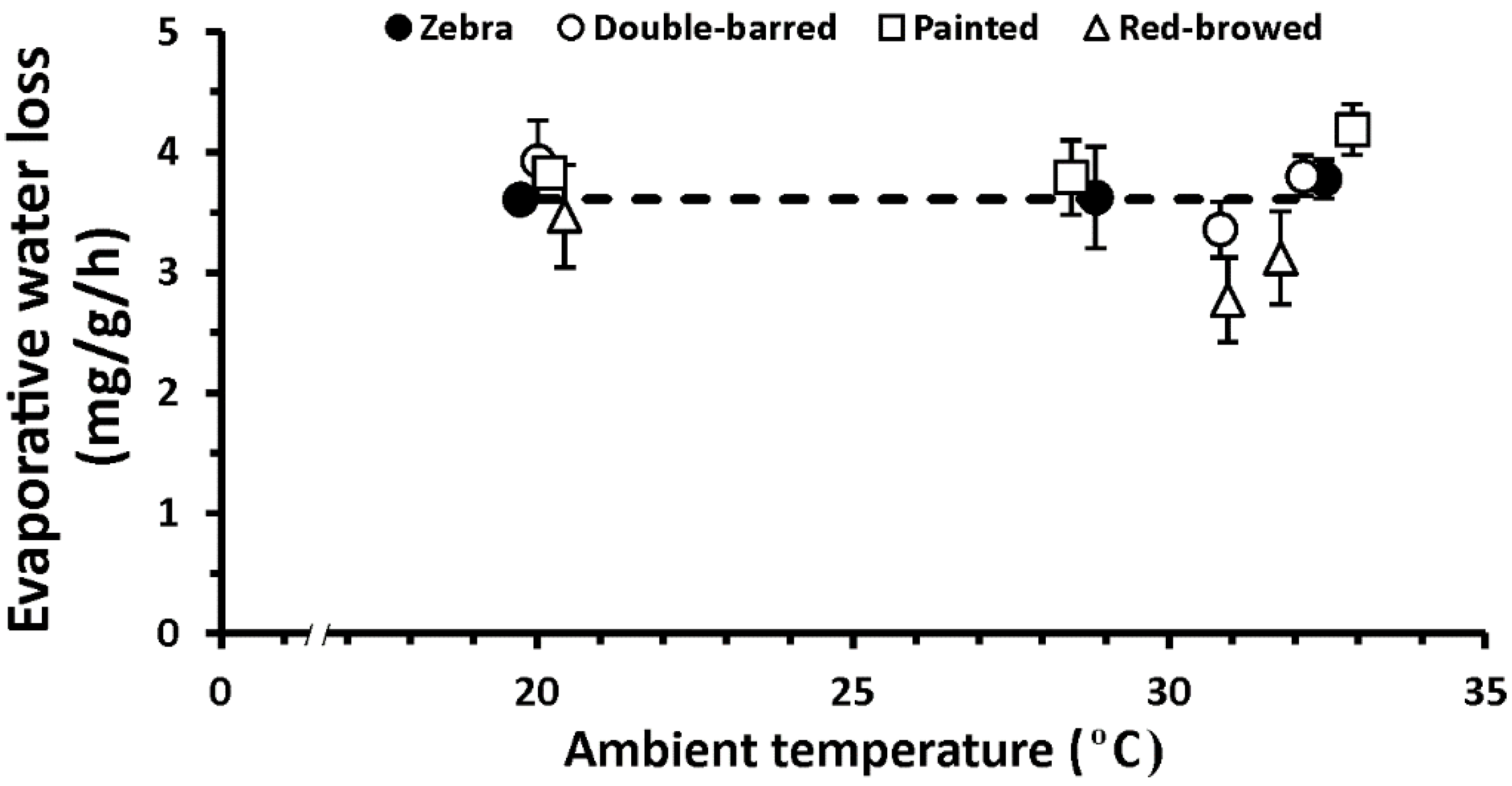
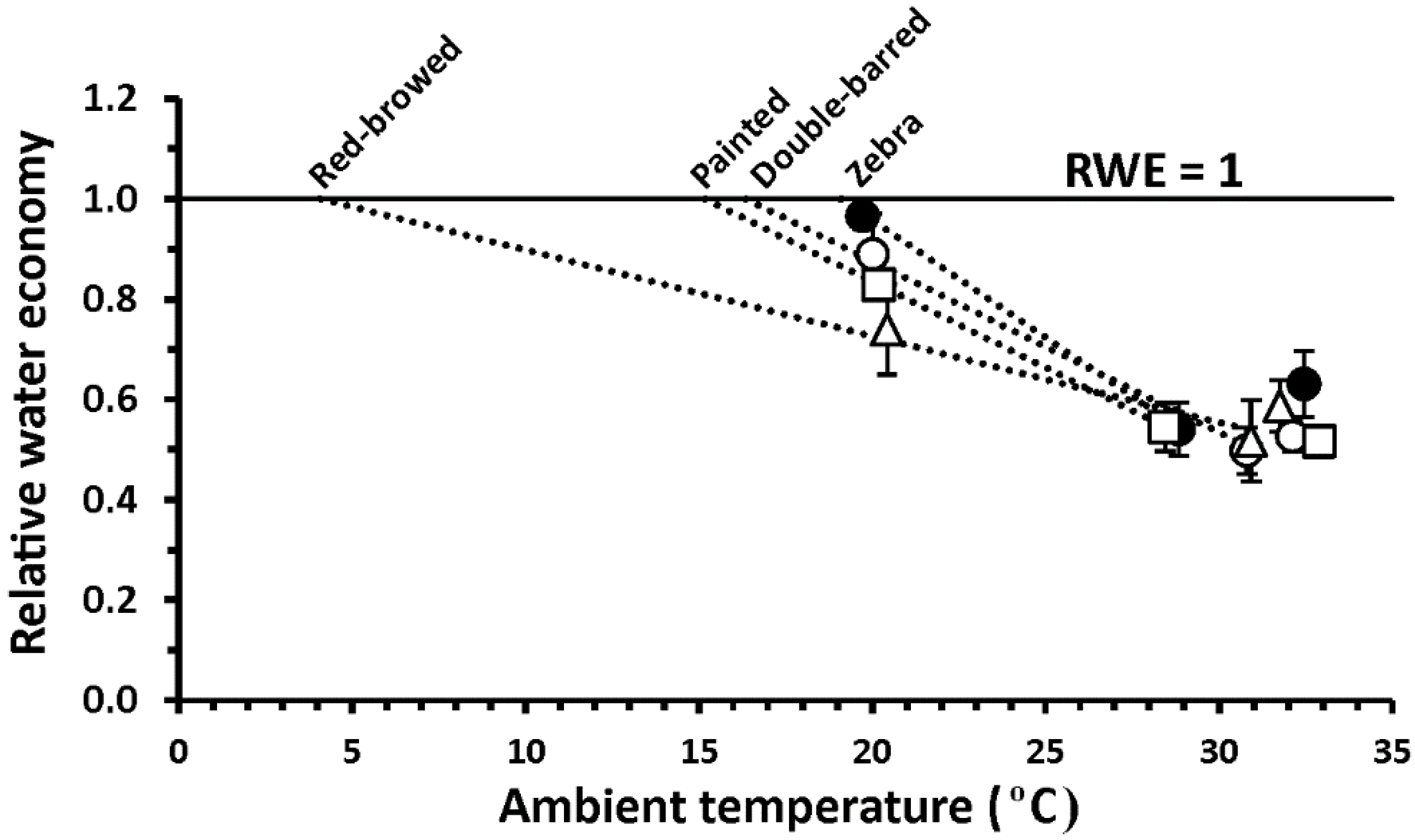
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Immelmann, K. Ecólogy and behaviour of African and Australian grass finches—A comparison. Ostrich 1966, 37, 371–379. [Google Scholar] [CrossRef]
- Immelmann, K. Australian Finches; Angus and Robertson: Sydney, Australia, 1982. [Google Scholar]
- Arnaiz-Villena, A.; Ruiz-del-Valle, V.; Gomez-Prieto, P.; Reguera, R.; Parga-Lozano, C.; Serrano-Vela, I. Estrildinae finches (Aves, Passeriformes) from Africa, South Asia and Australia: A molecular phylogeographic study. Open Ornithol. J. 2009, 2, 29–36. [Google Scholar] [CrossRef]
- Del Hoyo, J.; Elliot, A.; Christie, D. Handbook of Birds of the World, Volume 15: Weavers to New World Warblers; Lynx Edicions: Barcelona, Spain, 2010. [Google Scholar]
- Olsson, U.; Alström, P. A comprehensive phylogeny and taxonomic evaluation of the waxbills (Aves: Estrildidae). Mol. Phylogenet. Evol. 2020, 146, 106757. [Google Scholar] [CrossRef] [PubMed]
- Cade, T.J.; Tobin, C.A.; Gold, A. Water economy and metabolism of two estrildine finches. Physiol. Zool. 1965, 38, 9–33. [Google Scholar] [CrossRef]
- Barrett, G.; Silcocks, A.; Barry, S.; Cunningham, R.; Poulter, R. The New Atlas of Australian Birds; Royal Australasian Ornithologists Union: Victoria, Australia, 2003. [Google Scholar]
- Bartholomew, G.A.; Cade, T.J. The water economy of land birds. Auk 1963, 80, 504–539. [Google Scholar] [CrossRef]
- Bartholomew, G.A. The water economy of seed eating birds which survive without drinking. In Proceedings of the 15th International Ornithological Congress, The Hague, The Netherlands, 30 August–5 September 1970; Voous, K.H., Ed.; E. J. Brill: Leiden, The Netherlands, 1972; pp. 237–254. [Google Scholar]
- Griffith, S.C.; Buchanan, K.L. The Zebra Finch: The ultimate Australian supermodel. Emu 2010, 110, v–xii. [Google Scholar] [CrossRef]
- Griffith, S.C.; Ton, R.; Hurley, L.L.; McDiarmid, C.S.; Pacheco-Fuentes, H. The ecology of the Zebra Finch makes it a great laboratory model but an outlier amongst passerine birds. Birds 2021, 2, 60–76. [Google Scholar] [CrossRef]
- Calder, W.A.; King, J.R. Evaporative cooling in the zebra finch. Experientia 1963, 19, 603–604. [Google Scholar] [CrossRef] [PubMed]
- Calder, W.A. Gaseous metabolism and water relations of the zebra finch Taeniopygia castanotis. Physiol. Zool. 1964, 37, 400–413. [Google Scholar] [CrossRef]
- Skadhauge, E.; Bradshaw, S.D. Saline drinking and cloacal excretion of salt and water in the zebra finch. Am. J. Physiol. 1974, 227, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.E.; Withers, P.C.; Hurley, L.L.; Griffith, S.C. The field metabolic rate, water turnover, and feeding and drinking behavior of a small avian desert granivore during a summer heatwave. Front. Physiol. 2019, 10, 1405. [Google Scholar] [CrossRef]
- Cooper, C.E.; Hurley, L.L.; Griffith, S.C. Effect of acute exposure to high ambient temperature on the thermal, metabolic and hygric physiology of a small desert bird. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2020, 244, 110684. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.J.; Peter, J.M.; Cowling, S.J. Handbook of Australian, New Zealand and Antarctic Birds. Volume 7. Boatbill to Starlings; Oxford University Press: Melbourne, Australia, 2006. [Google Scholar]
- Menkhorst, P.; Rogers, D.; Clarke, R.; Davies, J.; Marsack, P.; Franklin, K. The Australian Bird Guide, Revised Edition; CSIRO Publishing: Clayton, Victoria, Australia, 2019. [Google Scholar]
- Oksche, A.; Farner, D.S.; Serventy, D.L.; Wolff, F.; Nicholls, C. The hypothalamo-hypophysial neurosecretory system of the Zebra Finch, Taeniopygia castanotis. Zeit. Zellforsch. Mikro. Anat. 1963, 58, 846–914. [Google Scholar] [CrossRef]
- Lee, P.; Schmidt-Nielsen, K. Respiratory and cutaneous evaporation in the zebra finch—Effect on water balance. Am. J. Physiol. 1971, 220, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.J.J.F. Nomadism as a response to desert conditions in Australia. J. Arid Environ. 1984, 7, 183–195. [Google Scholar] [CrossRef]
- Burbidge, A.A.; Fuller, P.J. Gibson desert birds: Responses to drought and plenty. Emu-Aust. Ornith. 2007, 107, 126–134. [Google Scholar] [CrossRef]
- Jordan, R.; James, A.I.; Moore, D.; Franklin, D.C. Boom and bust (or not?) among birds in an Australian semi-desert. J. Arid Environ. 2017, 139, 58–66. [Google Scholar] [CrossRef]
- MacMillen, R.E. Water economy of granivorous birds—A predictive model. Condor 1990, 92, 379–392. [Google Scholar] [CrossRef]
- Williams, J.B. A phylogenetic perspective of evaporative water loss in birds. Auk 1996, 113, 457–472. [Google Scholar] [CrossRef]
- MacMillen, R.E.; Hinds, D.S. Water economy of granivorous birds: California House Finches. Condor 1998, 100, 493–503. [Google Scholar] [CrossRef]
- Tieleman, B.I.; Williams, J.B.; Bloomer, P. Adaptation of metabolism and evaporative water loss along an aridity gradient. Proc. R. Soc. B Biol. Sci. 2003, 270, 207–214. [Google Scholar] [CrossRef]
- Williams, J.B.; Tieleman, B.I. Physiological adaptation in desert birds. Bioscience 2005, 55, 416–425. [Google Scholar] [CrossRef]
- White, C.R.; Blackburn, T.M.; Martin, G.R.; Butler, P.J. Basal metabolic rate of birds is associated with habitat temperature and precipitation, not primary productivity. Proc. R. Soc. B Biol. Sci. 2007, 274, 287–293. [Google Scholar] [CrossRef]
- McKechnie, A.E.; Gerson, A.R.; Wolf, B.O. Thermoregulation in desert birds: Scaling and phylogenetic variation in heat tolerance and evaporative cooling. J. Exp. Biol. 2021, 224 (Suppl. S1), jeb229211. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, G.A.; Dawson, W.R. Respiratory water loss in some birds of the south western United States. Physiol. Zool. 1953, 26, 162–166. [Google Scholar] [CrossRef]
- Jetz, W.; Freckleton, R.P.; McKechnie, A.E. Environment, migratory tendency, phylogeny and basal metabolic rate in birds. PLoS ONE 2008, 3, e3261. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Beissinger, S.R. Environmental determinants of total evaporative water loss in birds at multiple temperatures. Auk 2020, 137, ukz069. [Google Scholar] [CrossRef]
- Maclean, G.L. Ecophysiology of Desert Birds; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Bradshaw, D. Vertebrate Ecophysiology: An Introduction to its Principles and Applications; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Williams, J.B.; Tieleman, B.I. Ecological and evolutionary physiology of desert birds: A progress report. Integr. Comp. Biol. 2002, 42, 68–75. [Google Scholar] [CrossRef] [PubMed][Green Version]
- MacMillen, R.E.; Hinds, D.S. Water regulatory efficiency in heteromyid rodents. A model and its application. Ecology 1983, 64, 152–164. [Google Scholar] [CrossRef]
- Cortés, A.; Rosenmann, M.; Bozinovic, F. Water economy in rodents: Evaporative water loss and metabolic water production. Rev. Chil. Hist. Nat. 2000, 73, 311–321. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, B.; Schmidt-Nielsen, K. A complete account of the water metabolism in kangaroo rats and an experimental verification. J. Cell. Comp. Physiol. 1951, 38, 165–181. [Google Scholar] [CrossRef]
- Cooper, C.E.; Withers, P.C.; Turner, J.M. Physiological implications of climate change for a critically endangered Australian marsupial. Aust. J. Zool. 2020, 68, 200–211. [Google Scholar] [CrossRef]
- Dawson, W.R. Evaporative losses of water by birds. Comp. Biochem. Physiol. A Physiol. 1982, 71, 495–509. [Google Scholar] [CrossRef]
- MacMillen, R.E.; Baudinette, R.V. Water economy of granivorous birds: Australian parrots. Func. Ecol. 1993, 7, 704–712. [Google Scholar] [CrossRef]
- Cooper, C.E.; Withers, P.C. Comparative physiology of Australian quolls (Dasyurus; Marsupialia). J. Comp. Physiol. B 2010, 180, 857–868. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Withers, P.C. Design, calibration and calculation for flow-through respirometry systems. Aust. J. Zool. 2001, 49, 445–461. [Google Scholar] [CrossRef]
- Withers, P.C. Comparative Animal Physiology; Saunders College Publishing: Philadelphia, PA, USA, 1992. [Google Scholar]
- Henderson-Sellers, B. A new formula for latent heat of vaporization of water as a function of temperature. Quart. J. R. Met. Soc. 1984, 110, 1186–1190. [Google Scholar] [CrossRef]
- Douglas, T.K.; Cooper, C.E.; Withers, P.C. Avian torpor or alternative thermoregulatory strategies for overwintering? J. Exp. Biol. 2017, 220, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Statist. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.1. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 11 November 2021).
- Bicudo, J.E.P.W.; Buttemer, W.A.; Chappell, M.A.; Pearson, J.T.; Bech, C. Ecological and Environmental Physiology of Birds; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Marschall, U.; Prinzinger, R. Vergleichende ökophysiologie von fünf prachtfinkenarten (Estrildidae). J. Ornithol. 1991, 132, 319–323. [Google Scholar] [CrossRef]
- Bech, C.; Rønning, B.; Moe, M. Individual variation in the basal metabolism of Zebra finches Taeniopygia guttata: No effect of food quality during early development. Int. Congr. Ser. 2004, 1275, 306–312. [Google Scholar] [CrossRef]
- Rønning, B.; Moe, B.; Bech, C. Long-term repeatability makes basal metabolic rate a likely heritable trait in the zebra finch Taeniopygia guttata. J. Exp. Biol. 2005, 208, 4663–4669. [Google Scholar] [CrossRef]
- Rønning, B.; Jensen, H.; Moe, B.; Bech, C. Basal metabolic rate: Heritability and genetic correlations with morphological traits in the zebra finch. J. Evol. Biol. 2007, 20, 1815–1822. [Google Scholar] [CrossRef]
- Bernstein, M.H. Cutaneous water loss in small birds. Condor 1971, 73, 468–469. [Google Scholar] [CrossRef]
- Weathers, W.W.; Weathers, D.L.; Van Riper, C. Basal metabolism of the Apapane: Comparison of freshly caught birds with long-term captives. Auk 1983, 100, 977–978. [Google Scholar] [CrossRef]
- Warkentin, I.G.; West, N.H. Impact of long-term captivity on basal metabolism in birds. Comp. Biochem. Physiol. A 1990, 96, 379–381. [Google Scholar] [CrossRef]
- Cooper, C.E.; Withers, P.C. Effect of sampling regime on estimation of basal metabolic rate and standard evaporative water loss using flow-through respirometry. Physiol. Biochem. Zool. 2010, 83, 385–393. [Google Scholar] [CrossRef]
- Page, A.J.; Cooper, C.E.; Withers, P.C. Effects of experiment start time and duration on measurement of standard physiological variables. J. Comp. Physiol. B 2011, 181, 657–665. [Google Scholar] [CrossRef]
- Burton, C.T.; Weathers, W.W. Energetics and thermoregulation of the Gouldian Finch (Erythrura gouldiae). Emu 2003, 103, 1–10. [Google Scholar] [CrossRef]
- Aschoff, J.; Pohl, H. Rhythmic variations in energy metabolism. Fed. Proc. 1970, 29, 1541–1552. [Google Scholar]
- Willoughby, E.J. Evaporative water loss of a small xerophilous finch, Lonchura malabarica. Comp. Biochem. Physiol. 1969, 28, 655–664. [Google Scholar] [CrossRef]
- Lovegrove, B.G.; Smith, G.A. Is ‘nocturnal hypothermia’ a valid physiological concept in small birds?: A study on bronze mannikins Spermestes cucullatus. Ibis 2003, 145, 547–557. [Google Scholar] [CrossRef]
- Weathers, W.W. Temperature regulation in the dusky munia, Lonchura fuscens (Cassin) (Estrilidae). Aust. J. Physiol. 1977, 25, 193–199. [Google Scholar]
- Kendeigh, S.C.; Dol’nik, V.R.; Gavrilov, V.M. Avian energetics. In Granivorous Birds in Ecosystems; Pinowski, J., Kendeigh, S.C., Eds.; Cambridge University Press: Cambridge, UK, 1977; pp. 127–204. [Google Scholar]
- Qiao, Q.-G.; Liang, H.-J.; Bai, M.-L.; Zheng, W.-H.; Liu, J.-S. Interspecific variation of thermoregulation between migratory and resident passerines in Wenzhou. Zool. Res. 2016, 37, 167–175. [Google Scholar]
- Lasiewski, R.C.; Hubbard, S.H.; Moberly, W.R. Energetic relationships of a very small passerine bird. Condor 1964, 66, 212–220. [Google Scholar] [CrossRef]
- Stephens, C.M.; Siegel, R.B.; Weathers, W.W. Thermal conductance and basal metabolism of the Orange-cheeked Waxbill (Estrilda melpoda). Ostrich 2001, 72, 114–126. [Google Scholar] [CrossRef]
- Gavrilov, V.M.; Dol’nik, V.R. Basal metabolic rate, thermoregulation and existence energy in birds: World data. In Acta XVIII Congress Internationalis Ornithologici; Ilyichov, V.D., Gavrilov, V.M., Eds.; Nauka: Moscow, Russia, 1985; pp. 421–466. [Google Scholar]
- McKechnie, A.E.; Lovegrove, B.G. Facultative hypothermic responses in an Afrotropical arid-zone passerine, the red-headed finch (Amadina erythrocephala). J. Comp. Physiol. B 2003, 173, 339–346. [Google Scholar] [CrossRef]
- Cooper, C.E.; Withers, P.C. Effects of measurement duration on the determination of basal metabolic rate and evaporative water loss of small marsupials: How long is long enough? Physiol. Biochem. Zool. 2009, 82, 438–446. [Google Scholar] [CrossRef]
- Connolly, M.K.; Cooper, C.E. How do measurement duration and timing interact to influence estimation of basal physiological variables of a nocturnal rodent? Comp. Biochem. Physiol. A 2014, 178, 24–29. [Google Scholar] [CrossRef][Green Version]
- McKechnie, A.E.; Wolf, B.O. The allometry of avian basal metabolic rate: Good predictions need good data. Physiol. Biochem. Zool. 2004, 77, 502–521. [Google Scholar] [CrossRef]
- Bayly, I.A. Review of how indigenous people managed for water in desert regions of Australia. J. R. Soc. West. Aust. 1999, 82, 17–25. [Google Scholar]
- Pacheco-Fuentes, H.; Cooper, C.E.; Withers, P.C.; Griffith, S.C. Re-evaluating model assumptions suggests that Australian birds are more tolerant of heat and aridity than predicted: A response to Conradie et al. (2020). Cons. Physiol. 2022, 10, coac010. [Google Scholar] [CrossRef]
- Davies, S.J.J.F. Results of 40 hours’ continuous watch at five waterpoints in an Australian desert. Emu-Austral Ornithol. 1972, 72, 8–12. [Google Scholar] [CrossRef]
- Fisher, C.D.; Lindgren, E.; Dawson, W.R. Drinking patterns and behavior of Australian desert birds in relation to their ecology and abundance. Condor 1972, 74, 111–136. [Google Scholar] [CrossRef]
- Evans, S.M.; Collins, J.A.; Evans, R.; Miller, S. Patterns of drinking behaviour of some Australian estrildine finches. Ibis 1985, 127, 348–354. [Google Scholar] [CrossRef]
- Bowler, J.M. Aridity in Australia: Age, origins and expression in aeolian landforms and sediments. Earth-Sci. Rev. 1976, 12, 279–310. [Google Scholar] [CrossRef]
- Williams, J.B.; Tieleman, B.I. Physiological ecology and behavior of desert birds. In Current Ornithology; Nolan, V., Thompson, C.F., Eds.; Kluwer Academics/Plenum Publishers: New York, NY, USA, 2001; Volume 16, pp. 299–353. [Google Scholar]
- Dawson, W.R.; Bartholomew, G.A. Temperature regulation and water economy of desert birds. In Desert Biology; Brown, G.W., Ed.; Academic Press: New York, NY, USA, 1968; Volume 1, pp. 357–394. [Google Scholar]
- Cooper, C.E.; Withers, P.C.; Munns, S.L.; Geiser, F.; Buttemer, W.A. Geographical variation in the standard physiology of brushtail possums (Trichosurus): Implications for conservation translocations. Cons. Physiol. 2018, 6, coy042. [Google Scholar] [CrossRef]
- Conradie, S.R.; Woodborne, S.M.; Wolf, B.O.; Pessato, A.; Mariette, M.M.; McKechnie, A.E. Avian mortality risk during heat waves will increase greatly in arid Australia during the 21st century. Cons. Physiol. 2020, 8, coaa048. [Google Scholar] [CrossRef]
- Cooper, C.E.; Hurley, L.L.; Deviche, P.; Griffith, S.C. Physiological responses of wild zebra finches (Taeniopygia guttata) to heatwaves. J. Exp. Biol. 2020, 223, jeb225524. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Withers, P.C.; Cooper, C.E.; Larcombe, A.N. Relative Water Economy Is a Useful Index of Aridity Tolerance for Australian Poephiline Finches. Birds 2022, 3, 172-183. https://doi.org/10.3390/birds3020012
Withers PC, Cooper CE, Larcombe AN. Relative Water Economy Is a Useful Index of Aridity Tolerance for Australian Poephiline Finches. Birds. 2022; 3(2):172-183. https://doi.org/10.3390/birds3020012
Chicago/Turabian StyleWithers, Philip C., Christine E. Cooper, and Alexander N. Larcombe. 2022. "Relative Water Economy Is a Useful Index of Aridity Tolerance for Australian Poephiline Finches" Birds 3, no. 2: 172-183. https://doi.org/10.3390/birds3020012
APA StyleWithers, P. C., Cooper, C. E., & Larcombe, A. N. (2022). Relative Water Economy Is a Useful Index of Aridity Tolerance for Australian Poephiline Finches. Birds, 3(2), 172-183. https://doi.org/10.3390/birds3020012