

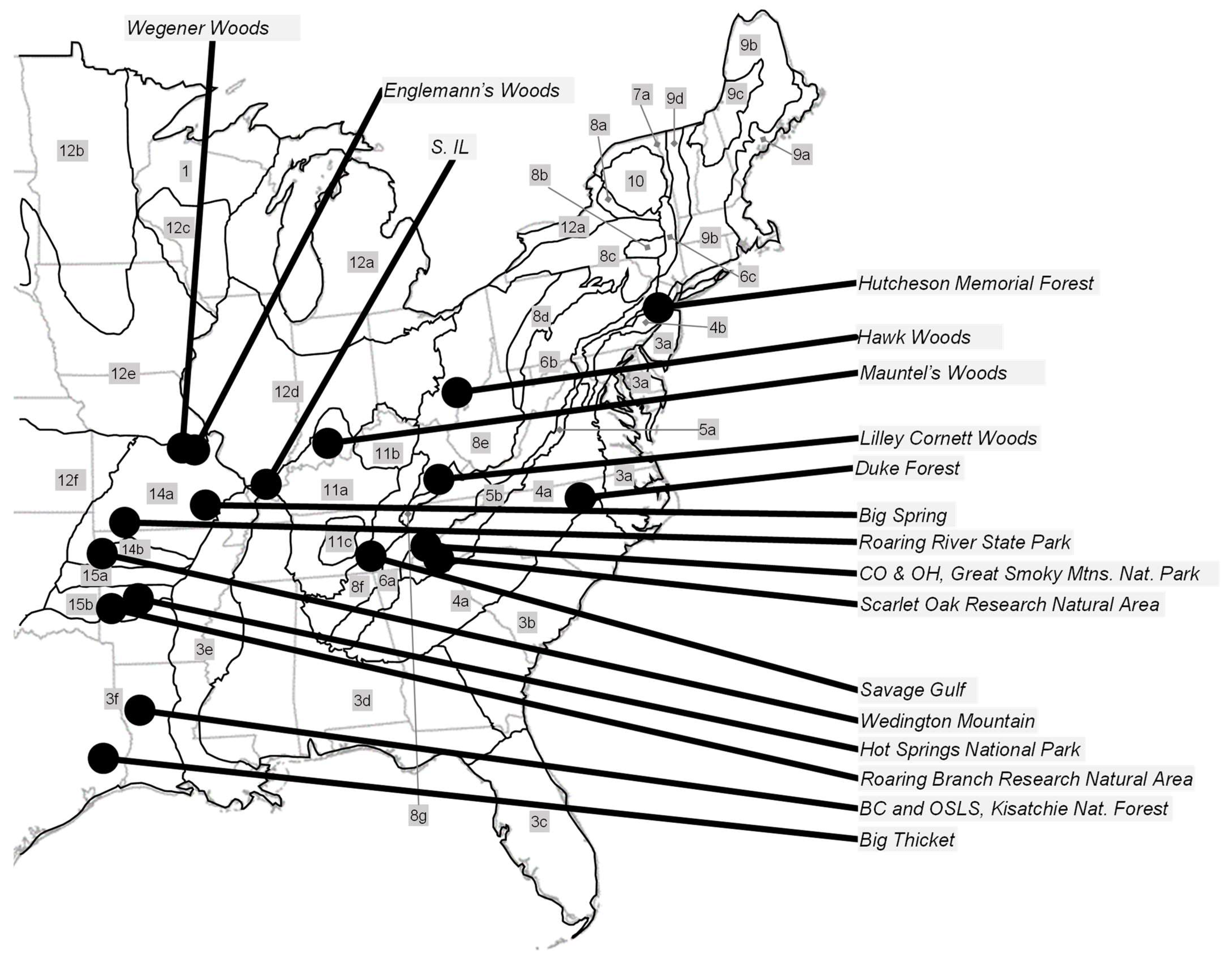
Characteristics of Dry-Mesic Old-Growth Oak Forests in the Eastern United States

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Physiographic Region
3.1.1. Piedmont
3.1.2. Appalachian Plateaus
3.1.3. Ridge and Valley

{kind=link}
{kind=link}
| Quantifiable Attribute | Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| STAND DENSITY (# ha−1) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| (≥10 cm dbh) | 556–564 | 560 | 1 | [23] [24] |
| Duke Forest | ||||
| White oak (≥2.5 cm dbh) | 1789–1853 | 1831 | 3 | [13] |
| Post oak (≥2.5 cm dbh) | 1460–2300 | 1826 | 3 | |
| Appalachian Plateaus | ||||
| Allegheny Plateau | ||||
| Hawk Woods | ||||
| (≥10 cm dbh) | 250–443 | 341 | 3 | [16] |
| [24] | ||||
| Cumberland Plateau | ||||
| Lilley Cornett Woods | ||||
| (≥12.7 cm dbh) | 289–395 | 346 | 5 | [17] |
| Chestnut oak-R. maple (≥10 cm dbh) | 450 | 1 | [19] | |
| Savage Gulf | ||||
| Chestnut oak (≥12.7 cm dbh) | 398 | 1 | [21] | |
| Hickory-Red oak (≥12.7 cm dbh) | 454 | 1 | [21] | |
| White oak (≥10 cm dbh) | 620 | 12 | [25] | |
| Ridge and Valley | ||||
| South Central Pennsylvania | ||||
| Ridge and upper slopes (≥10 cm dbh) | 408–714 | 521 | 9 | [22] 1991 (Data) |
| Middle Slopes (≥10 cm dbh) | 403–546 | 477 | 4 | |
| Blue Ridge Mountains | ||||
| Great Smoky Mountain NP | ||||
| Oak-Hickory (≥10 cm dbh) | 403 | 1 | [26] (data) | |
| Chestnut oak (≥10 cm dbh) | 620 | 1 | [26] (data) | |
| Scarlet Oak | ||||
| (≥7.6 cm dbh) | 620–991 | 783 | 4 | [27] |
| Interior Low Plateau | ||||
| Shawnee Hills | ||||
| Southern Indiana—Mauntel’s Woods | ||||
| (≥10 cm dbh) | 430 | [28] | ||
| Southern Illinois | ||||
| South Slope (≥10 cm dbh) | 415 | 1 | [29] p 300 | |
| Upper North Slope (≥10 cm dbh) | 377 | 1 | ||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Hot Springs N.P. (≥9 cm dbh) | 536–1527 | 1206 | 4 | [30] |
| Roaring Branch R.N.A. | ||||
| Upper & Mid N Slope (≥15 cm dbh) | 368 | 1 | [31] | |
| Boston Mountains | ||||
| Upper West Slope (≥15 cm dbh) | 600 | 1 | [32] | |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| 1992 | ||||
| North & East Slopes (≥10 cm dbh) | 310–600 | 442 | 1 | [33] (1992 Data) |
| Ridge + S & W Slopes (≥10 cm dbh) | 300–580 | 443 | 1 | [33] (1992 Data) |
| 2011 | ||||
| North & East Slopes (≥10 cm dbh) | 240–590 | 390 | 1 | [34] (2011 Data) |
| Ridge + S & W Slopes (≥10 cm dbh) | 280–600 | 435 | 1 | [34] (2011 Data) |
| Salem Plateau | ||||
| Big Spring (≥10 cm dbh) | 310–770 | 469 | 1 | [33] (1992 Data) |
| Big Spring (≥10 cm dbh) | 280–560 | 402 | 1 | [34] (2011 Data) |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + S & W Slopes (≥10 cm dbh) | 460–670 277–530 | 535 407 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Wegener Woods (≥9 cm dbh) | 376 | 1 | [35] | |
| Southern Coastal Plain | ||||
| Western Gulf | ||||
| Kisatchie Natl Forest (≥12.7 cm dbh) | ||||
| Old Shortleaf Slope | 280 | 1 | [36] | |
| Bob’s Creek | 151 | 1 |
| Quantifiable Attribute | Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| BASAL AREA (m2 ha−1) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| (≥10 cm dbh) | 26–27 | 27 | 1 | [24] |
| Duke Forest | ||||
| White oak (≥2.5 cm dbh) | 37–44 | 40 | 3 | [13] |
| Post oak (≥2.5 cm dbh) | 27–38 | 32 | 3 | |
| Appalachian Plateaus | ||||
| Allegheny and Cumberland Plateaus | ||||
| Hawk Woods | ||||
| (≥10 cm dbh) | 22–41 | 29 | 3 | [16] |
| Lilley Cornett Woods | ||||
| (≥12.7 cm dbh) | 24–29 | 26 | 5 | [17] |
| Chestnut oak-red maple (≥10 cm dbh) | 26 | 1 | [19] | |
| Savage Gulf | ||||
| Chestnut oak (≥12.7 cm dbh) | 37 | 1 | [21] | |
| Hickory-Red oak (≥12.7 cm dbh) White oak (≥10 cm dbh) | 37 29 | 1 12 | [25] | |
| Ridge and Valley | ||||
| South Central Pennsylvania | ||||
| Ridge & upper slopes (≥10 cm dbh) | 22–38 | 29 | 9 | [22] 1991 (Data) |
| Middle Slopes (≥10 cm dbh) | 31–42 | 36 | 4 | |
| Blue Ridge Mountains | ||||
| Great Smoky Mountain NP | ||||
| Oak-Hickory (≥10 cm dbh) | 28 | 1 | [26] | |
| Chestnut oak (≥10 cm dbh) | 30 | 1 | [26] | |
| Scarlet Oak | ||||
| (≥7.6 cm dbh) | 17–26 | 22 | 4 | [27] |
| Interior Low Plateau | ||||
| Shawnee Hills | ||||
| Southern Indiana—mauntel’s woods | ||||
| (≥10 cm dbh) | 33 | 1 | [28] | |
| Southern Illinois | ||||
| South Slope (≥10 cm dbh) | 21 | 1 | [29] | |
| Upper North Slope (≥10 cm dbh) | 26 | 1 | ||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Hot Springs N.P. (≥9 cm dbh) | 13–23 | 19 | 4 | [30] |
| Roaring Branch R.N.A. (≥10 cm dbh) | ||||
| Upper and Mid North Slope | 18 | 1 | [31] | |
| Boston Mountains | ||||
| Upper West Slope (≥15 cm dbh) | 25 | 1 | [32] | |
| Springfield Plateau | ||||
| Roaring River State Park 1992 | ||||
| North & East Slopes (≥10 cm dbh) | 14–32 | 23 | 1 | [33] (1992 Data) |
| Ridge + S & W Slopes (≥10 cm dbh) | 12–28 | 20 | 1 | |
| 2011 | ||||
| North & East Slopes (≥10 cm dbh) | 15–33 | 23 | 1 | [34] (2011 Data) |
| Ridge + S & W Slopes (≥10 cm dbh) | 16–29 | 21 | 1 | |
| Salem Plateau | ||||
| Big Spring (≥10 cm dbh) | 17–28 | 22 | [33] (1992 Data) | |
| Big Spring (≥10 cm dbh) | 15–32 | 22 | 1 | [34] (2011 Data) |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| 1992 | ||||
| Ridge + S & W Slopes (≥10 cm dbh) | 16–29 | 22 | 1 | [33] (1992 Data) |
| 2011 | ||||
| Ridge + S & W Slopes (≥10 cm dbh) | 12–27 | 21 | 1 | [34] (2011 Data) |
| Wegener Woods (≥10 cm dbh) | 20 | 1 | [35] | |
| Southern Coastal Plain | ||||
| Western Gulf | ||||
| Kisatchie National Forest (≥12.7 cm dbh) | ||||
| Old Shortleaf Slope | 32 | 1 | [36] | |
| Bob’s Creek | 14 | 1 | ||
| Big Thicket (≥1 cm dbh) | ||||
| Mid Slope Oak Pine Forest | 32 | 1 | [37] |
3.1.4. Blue Ridge Mountains
3.1.5. Interior Low Plateaus
3.1.6. Interior Highlands
3.1.7. Southern Coastal Plain: Gulf Coast
3.2. Old Growth Conditions
3.2.1. Ownership
3.2.2. Living Tree Component
| Quantifiable Attribute | Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| AGE OF LARGE TREES (years) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| White oak | ||||
| Red oak | 200–324 | 1 | [52] | |
| Black oak | [53] | |||
| Ridge and Valley | ||||
| South Central Pennsylvania | ||||
| Ridge and Upper Slopes | ||||
| N. Red oak and black oak | 84–196 | 119 | 9 | [22] 1991 (Data) |
| Chestnut oak | 84–365 | 170 | ||
| Sweet birch | 102–230 | 129 | ||
| White pine | 101–119 | 112 | ||
| Middle Slopes | ||||
| N. Red oak and black oak | 80–236 | 123 | 4 | [22] 1991 (Data) |
| Chestnut oak | 80–324 | 159 | ||
| Mockernut hickory | 133–197 | 171 | ||
| Blue Ridge Mountains | ||||
| Scarlet Oak | ||||
| Scarlet oak | 73–123 | 2 | [44] | |
| Chestnut oak | 66–170 | |||
| Pitch pine | 117–163 | |||
| Chattooga Watershed | Max.Age (dbh-cm) | |||
| Chestnut oak | 362 (53) | [54] | ||
| White oak | 348 (69) | |||
| Northern Red oak | 240 (66) | |||
| Post oak | 225 (36) | |||
| Black oak | 211 (46) | |||
| Scarlet oak | 190 (61) | |||
| Blackgum | 360 (61) | |||
| Mockernut hickory | 335 (66) | |||
| Shortleaf pine | 300 (71) | |||
| Pitch pine | 235 (69) | |||
| Virginia pine | 108 (46) | |||
| Blue Ridge Mountains | ||||
| Great Smoky Mountain NP | Max. Age (dbh-cm) | |||
| Chestnut oak | 347 (56) | [49] | ||
| White oak | 344 (69) | |||
| Northern Red oak | 270 (56) | |||
| Black oak | 180 (66) | |||
| Scarlet oak | 165 (41) | |||
| Blackgum | 319 (43) | |||
| Red maple | 220 (66) | |||
| Pignut hickory | 327 (46) | |||
| Pitch pine | 148 | |||
| Virginia pine | 135 (64) | |||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Hot Springs N.P. | ||||
| Shortleaf pine | 124–247 | 4 | [40] | |
| Roaring Branch RNA | ||||
| Northern Red oak | 65–120 | 110 | 1 | [39] |
| White oak | 51–150 | 130 | 1 | [31] |
| Shortleaf pine | 121–300 | 150+ | 1 | [40] |
| Boston Mountains | ||||
| Upper West Slope | ||||
| Post oak | 140-253 | 192 | 1 | [32] |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + S & W Slopes | ||||
| White oak | 120 | 1 | [33] | |
| Black oak | 103 | |||
| Chinkapin oak | 139–204 | 172 | ||
| Wegener Woods | [40] | |||
| White oak | 250-320 | 1 | [35] | |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| White oak | 200–250 | 1 | [40] | |
| Post oak | 200–300 | 1 | ||
| Salem Plateau | ||||
| Big Spring | ||||
| White oak | 87–137 | 120 | 1 | [33] |
| Black oak | 107–135 | 120 | ||
| Post oak | (max >150) | 141 | ||
| Shortleaf pine | 106–116 | 112 |
| Quantifiable Attribute | Value Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| DBH OF LARGE TREES (cm) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| White oak | 51–102 | 1 | [48] | |
| Black oak | 51–102 | |||
| Northern Red oak & scarlet oak | 51–76 | |||
| Red Hickory | 38–64 | |||
| Ridge and Valley | ||||
| South Central Pennsylvania | ||||
| Ridge and Upper Slopes | ||||
| Northern Red oak & black oak | 31–64 | 41 | 9 | [22] 1991 (Data) |
| Chestnut oak | 31–89 | 43 | 1 | |
| Birch | 31–46 | 38 | ||
| White pine | 33 | |||
| Middle Slopes | ||||
| Northern Red oak & black oak | 31–69 | 43 | 4 | [22] 1991 (Data) |
| Chestnut oak | 31–99 | 46 | ||
| Hickory | 31–51 | 36 | ||
| Blue Ridge Mountains | ||||
| Great Smoky Mountain NP | ||||
| Oak-Hickory | ||||
| Northern Red oak | 30–86 | 50 | 1 | [26] (data) |
| White oak | 37–83 | 60 | ||
| Chestnut oak | 26–53 | 38 | ||
| Red maple | 23–34 | 27 | ||
| Chestnut Oak | ||||
| Chestnut oak | 18–78 | 47 | 1 | [26] (data) |
| Scarlet oak | 26–34 | 29 | ||
| Northern Red oak | 26–41 | 32 | ||
| Red maple | 23–34 | 27 | ||
| Scarlet Oak | ||||
| Scarlet oak | 36–51 | 4 | [27] | |
| Chestnut oak | 36–46 | |||
| White oak | 36–46 | |||
| Pitch pine | 46 | |||
| Mockernut and Pignut hickory | 36–51 | |||
| Interior Low Plateau | ||||
| Shawnee Hills | ||||
| Southern Indiana—Mauntel’s Woods | ||||
| White oak | 51–81 | [28] | ||
| Black oak | 51–81 | |||
| Northern Red oak | 38–51 | |||
| Shagbark hickory | 25–38 | |||
| Mockernut hickory | 25–38 | |||
| Pignut hickory | 25–38 | |||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Roaring Branch RNA | ||||
| Upper and Mid North Slope | ||||
| Northern Red oak | 33–46 | 41 | 1 | [31] |
| Black oak | 33–46 | 36 | ||
| White oak | 33–61 | 41 | ||
| Blackgum | 31–46 | 38 | ||
| Boston Mountains | ||||
| Upper West Slope | ||||
| Post oak | 28–61 | 41 | 1 | [32] |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| White oak | 33–79 | 51 | [34] (2011 Data) | |
| Black oak | 33–69 | 46 | ||
| Post oak | 31–51 | 41 | ||
| Northern Red oak | 33–64 | 43 | ||
| Blackgum | 31–61 | 41 | ||
| Mockernut hickory | 31–48 | 38 | ||
| Black hickory | 31–48 | 38 | ||
| Salem Plateau | ||||
| Big Spring Natural Area | ||||
| White oak | 31–69 | 46 | 1 | [34] (2011 Data) |
| Black oak | 36–69 | 48 | ||
| Post oak | 36–53 | 43 | ||
| Scarlet oak | 33–66 | 48 | ||
| Southern Red oak | 31–61 | 46 | ||
| Northern Red oak | 31–56 | 43 | ||
| Shortleaf pine | 31–53 | 41 | ||
| Missouri River Hills | ||||
| Englemann Woods | ||||
| White oak | 36–43 | 41 | 1 | [34] (2011 Data) |
| Chinkapin oak | 23–69 | 36 | ||
| Northern Red oak | 28–61 | 43 | ||
| Shagbark hickory | 31–46 | 36 | ||
| Sugar maple | 28–41 | 31 | ||
| Eastern Red cedar | 23–41 | 28 | ||
| Missouri River Hills | ||||
| Wegener Woods | ||||
| White oak | 31–102 | 43 | 1 | [35] |
| N. Red oak | 41 | |||
| Shagbark hickory | 33 | |||
| Sugar maple | 18 | |||
| Southern Coastal Plain | ||||
| Western Gulf | ||||
| Kisatchie National Forest | ||||
| Old Shortleaf Slope | 25–76 | 41 | 1 | [36] |
| Bob’s Creek | 25–76 | 48 | 1 |
| Quantifiable Attribute | Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| NUMBER OF 10 CM. SIZE CLASSES (Starting at 10 cm) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | 9 | 1 | [48] | |
| Appalachian Plateaus | ||||
| Allegheny and Cumberland Plateaus | ||||
| Hawk Woods | ||||
| (≥10 cm dbh) | 7 | 2 | [16] | |
| Ridge and Valley | ||||
| South Central Pennsylvania | ||||
| Ridge and upper slopes (≥10 cm dbh) | 4–8 | 6 | 9 | [22] (Data) |
| Middle Slopes (≥10 cm dbh) | 4–9 | 6 | 4 | |
| Blue Ridge Mountains | ||||
| Great Smoky Mountain NP | ||||
| Oak-Hickory (≥10 cm dbh) | 7 | 1 | [26] (data) | |
| Chestnut oak (≥10 cm dbh) | 7 | 1 | [26] (data) | |
| Scarlet Oak | ||||
| (≥10 cm dbh) | 4–5 | 4 | 4 | [27] |
| Interior Low Plateau | ||||
| Shawnee Hills | ||||
| Southern Indiana—Mauntel’s Woods | ||||
| (≥10 cm dbh) | 7 | 1 | [28] | |
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Roaring Branch RNA | ||||
| Upper & Mid North Slope | ||||
| (≥10 cm dbh) | 5 | 1 | [31] | |
| Boston Mountains | ||||
| Upper West Slope (≥10 cm dbh) | 6 | 1 | [32] | |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| (≥10 cm dbh) | 7 | 1 | [34] (2011 Data) | |
| Salem Plateau | ||||
| Big Springs | ||||
| (≥10 cm dbh) | 6 | 1 | [34] (2011 data) | |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + S & W Slopes (≥10 cm dbh) | 8 | 1 | [34] (2011 Data) | |
| Wegener Woods (≥10 cm dbh) | 9 | 1 | [35] | |
| Southern Coastal Plain | ||||
| Western Gulf | ||||
| Kisatchie National Forest | ||||
| Old Shortleaf Slope (≥12.7 cm dbh) | 5 1 | 1 | [36] | |
| Bob’s Creek (≥12.7 cm dbh) | 5 1 | 1 |
3.2.3. Dead Tree Component
| Quantifiable Attribute | Value Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| STANDING SNAGS (# ha−1) except as noted below | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| (15 cm–86.4 cm dbh) | 15 | 1 | [56] | |
| Appalachian Plateaus | ||||
| Allegheny and Cumberland Plateaus | ||||
| Lilley Cornett Woods | ||||
| (≥10 cm dbh) | 78 | 1 | [57] | |
| Ridge and Valley | ||||
| Southwest Virginia | ||||
| (≥10 cm dbh) | 64 | [62] | ||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Hot Springs N.P. | ||||
| Mass only (≥ 7.6 cm dbh) Mg ha−1 | 14.4–25.1 | 19.8 | 2 | [63] |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| North and East slopes (≥10 cm dbh) | 0–70 0–80 | 30.6 34.7 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Ridge + S & W slopes (≥10 cm dbh) | 10–130 0–80 | 53.8 37.7 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Salem Plateau | ||||
| Big Spring Natural Area (≥10 cm dbh) | 0–110 0–130 | 38.0 43.3 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + S & W Slopes (≥10 cm dbh # ha−1) | 0–140 10–70 | 43.0 32.0 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Quantifiable Attribute | Value Range | Mean | No. of Stands | References |
|---|---|---|---|---|
| DOWNED CWD (either m3 ha−1 or Mg ha−1 as noted below) | ||||
| Piedmont | ||||
| Hutcheson Memorial Forest | ||||
| (≥10 cm–Mg ha−1) | 21.8 | 1 | [64] | |
| Appalachian Plateaus | ||||
| Allegheny and Cumberland Plateaus | ||||
| Lilley Cornett Woods | ||||
| (≥20 cm) m3 ha−1 | 23–85 | 64.7 | 1 | [65] |
| (≥20 cm) Mg ha−1 | 4.5–24 | 20.6 | 1 | |
| Blue Ridge Mountains | ||||
| Great Smokey Mountain NP | ||||
| Oak-Hickory (≥10 cm; m3 ha−1) | 182.5 | [26] | ||
| Chestnut oak (≥10 cm; m3 ha−1) | 74.0 | [26] | ||
| Interior Highlands | ||||
| Ouachita Mountains | ||||
| Hot Springs N.P. | ||||
| Mass only (≥7.6 cm.) Mg ha−1 | 1.6–9.4 | 5.6 | 2 | [63] |
| Springfield Plateau | ||||
| Roaring River State Park | ||||
| North and East Slopes (≥10 cm; m3 ha−1) | 6.5–87.1 9.4–67.4 | 34.2 32.5 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Ridge + South & West Slopes | ||||
| (≥ 10 cm; m3 ha−1) | 7.4–128.1 9.1–59.2 | 37.2 25.9 | 1 | [33] (1992 Data) [34] (2011 Data) |
| Salem Plateau | ||||
| Big Spring Natural Area | ||||
| (≥10 cm; m3 ha−1) | 5.3–96.4 6.4–121.0 | 32.0 48.5 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + South & West Slopes (≥10 cm dbh; m3 ha−1) | 2.5–94.6 6.3–62.5 | 23.0 30.2 | 1 1 | [33] (1992 Data) [34] (2011 Data) |
3.2.4. Other Components
4. Discussion
4.1. Ownership
4.2. Forest Dynamics and Succession
4.2.1. Site
4.2.2. Regeneration
4.2.3. Stand Development
4.2.4. Disturbance
4.3. Other Characteristics and Suggestions for the Future of These Forests
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Tree Species Characteristic of Old-Growth Dry Mesic Oak Forests
| Piedmont | Appalachian Mts. | Interior Low Plateaus | Interior Highlands | Coastal Plain | Percent of Sites where This Species Occurred | |||||||||||||||||||
| Allegheny/ Cumberland Pl. | Ridge & Valley | Blue Ridge | S. IN | S. ILL | Ouachita Mts. | Bost. Mts. | Springfield Pl. | Salem Pl. | Missouri River Hills | Western Gulf | ||||||||||||||
| SPECIES | HMF a | DF | HW | LCW | SG | Upper Slope | Mid-Slope | SO | CO | OH | MW | S. Slope | Upper N. Slope | HSPNP | RBRNA | WM | RRSP | BS | EW | WW | BC | OSLS | BT | |
| Acer saccharum | X | X | X | X | X | X | X | X | 35 | |||||||||||||||
| Acer rubrum | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 61 | |||||||||
| Carya cordiformis | X | X | X | 13 | ||||||||||||||||||||
| Carya glabra | X | X | X | X | X | X | 26 | |||||||||||||||||
| Carya ovalis | X | X | X | X | 17 | |||||||||||||||||||
| Carya ovata | X | X | X | X | X | X | 26 | |||||||||||||||||
| Carya texana | X | X | X | X | X | X | 26 | |||||||||||||||||
| Carya tomentosa | X | X | X | X | X | X | X | X | X | X | X | 48 | ||||||||||||
| Carya spp. | X | X | 9 | |||||||||||||||||||||
| Cercis canadensis | X | X | X | 13 | ||||||||||||||||||||
| Cornus florida | X | X | X | X | X | X | X | X | X | X | 43 | |||||||||||||
| Fagus grandifolia | X | X | X | X | X | 22 | ||||||||||||||||||
| Fraxinus americana | X | X | X | X | X | X | X | X | X | 39 | ||||||||||||||
| Fraxinus pennsylvanica | X | 4 | ||||||||||||||||||||||
| Ilex opaca | X | 4 | ||||||||||||||||||||||
| Ilex vomitoria | X | 4 | ||||||||||||||||||||||
| Liquidambar styraciflua | X | 4 | ||||||||||||||||||||||
| Liriodendron tulipifera | X | X | X | 13 | ||||||||||||||||||||
| Magnolia fraseri | X | 4 | ||||||||||||||||||||||
| Nyssa sylvatica | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 70 | |||||||
| Ostrya virginiana | X | X | X | X | X | 22 | ||||||||||||||||||
| Oxydendrum arboretum | X | X | X | X | X | X | X | 30 | ||||||||||||||||
| Persea borbonia | X | 4 | ||||||||||||||||||||||
| Pinus echinata | X | X | X | X | X | X | X | 30 | ||||||||||||||||
| Pinus strobus | X | X | 9 | |||||||||||||||||||||
| Pinus taeda | X | X | X | X | 17 | |||||||||||||||||||
| Sassafras albidum | X | 4 | ||||||||||||||||||||||
| Symplocos tinctoria | X | 4 | ||||||||||||||||||||||
| Quercus alba | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 91 | ||
| Quercus coccinea | X | X | X | X | X | X | X | X | X | 39 | ||||||||||||||
| Quercus falcata | X | X | X | 13 | ||||||||||||||||||||
| Quercus nigra | X | 4 | ||||||||||||||||||||||
| Quercus phellos | X | 4 | ||||||||||||||||||||||
| Quercus marilandica | X | X | X | X | X | 22 | ||||||||||||||||||
| Quercus prinus | X | X | X | X | X | X | X | X | 35 | |||||||||||||||
| Quercus rubra | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 83 | ||||
| Quercus stellata | X | X | X | X | X | X | X | X | X | X | 43 | |||||||||||||
| Quercus velutina | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | 70 | |||||||
| Robinia pseudoacacia | X | X | X | 13 | ||||||||||||||||||||
| Tsuga canadensis | X | X | 9 | |||||||||||||||||||||
| a HMF = Hutcheson Memorial Forest [48]. Not listed in the table above and only found on this site: Prunus avium, Acer Platanoides, Ulmus americana, Quercus palustris and Quercus Bicolor. DF = Duke Forest [13]. HW = Hawk Woods, white oak, oak-hickory and oak-heath types [16]. LCW = Lilley Cornett Woods, chestnut oak-red maple species group [19]. Lilley Cornett Woods, white oak, mixed oak, chestnut oak communities [17]. SG = Savage Gulf, mockernut hickory-northern red oak and chestnut oak community types [21]. Ridge and Valley = unpublished plot data from upper and mid-slopes, central Pennsylvania, G. Nowacki. SO = Scarlet Oak [27]. CO = Chestnut oak forest, Great Smoky Mountains National Park [26]. OH = Oak-hickory forest, Great Smoky Mountains National Park [26]. MW = Mauntel’s Woods [28]. S. IL = Southern Illinois [29]. HSNP = Hot Springs National Park, upland hardwoods and oak-hickory-pine types [30]. RBRNA = Roaring Branch Research Natural Area, mid and upper north slopes [31]. WM = Boston Mts. = Wedington Mountain [32]. RRSP = Roaring River State Park (unpublished data provided by S. Shifley, USDA-Forest Service, North Central Experiment Station, Columbia, MO, USA). BS = Big Spring (unpublished data provided by S. Shifley, USDA-Forest Service, North Central Experiment Station, Columbia, MO, USA). EW = Englemanns’s Woods, (unpublished data provided by S. Shifley, USDA-Forest Service, North Central Experiment Station, Columbia, MO). WW = Wegener Woods [35]. BC = Bob’s Creek stand, Kisatchie National Forest [36]. OSLS = Old Shortleaf Slope stand, Kisatchie National Forest [36]. BT = Big Thicket, midslope oak-pine forest [37]. An X in the Table Represents the Presence of a Species within the Respective Site. | ||||||||||||||||||||||||
Appendix B. Shrub and Vine Species Characteristic of Old-Growth Dry Mesic Oak Forests
| Piedmont | Appalachian Mts. | Interior Low Plateaus | Interior Highlands | Coastal Plain | ||||||||||
| Alleghany/ Cumberland Pl. | Blue Ridge | S.IN | Ouachita Mts. | Salem Pl. | Missouri R. Hills | Western Gulf | ||||||||
| SPECIES | HMF a | DF | SG | SO | CO | OH | MW | HSNP | RBRNA | BS | EW | WW | BC | OSLS |
| Calycanthus floridus | X | |||||||||||||
| Chionanthus virginicus | X | |||||||||||||
| Crataegus spp. | X | X | ||||||||||||
| Gaylussacia baccata | X | X | X | |||||||||||
| Hamamelis virginiana | X | |||||||||||||
| Kalmia latifolia | X | X | X | X | ||||||||||
| Lindera benzoin | X | |||||||||||||
| Lonicera japonica | X | |||||||||||||
| Parthenocissus quinquefolia | X | X | X | X | X | X | X | |||||||
| Polycodium spp. | X | |||||||||||||
| Rhododendron calendulaceum | X | |||||||||||||
| Rhododendron maximum | X | |||||||||||||
| Rubus spp. | X | X | ||||||||||||
| Rosa multiflora | X | |||||||||||||
| Rosa virginiana | X | |||||||||||||
| Smilax glauca | X | X | ||||||||||||
| Smilax herbacea | X | |||||||||||||
| Smilax rotundifolia | X | X | ||||||||||||
| Smilax spp. | X | X | X | |||||||||||
| Toxicodendron radicans | X | X | X | X | X | X | ||||||||
| Vaccinium arboreum | X | X | X | X | ||||||||||
| Vaccinium pallidum | X | X | ||||||||||||
| Vaccinium stamineum | X | X | X | |||||||||||
| Vaccinium vacillans | X | X | X | |||||||||||
| Vaccinium spp. | X | X | ||||||||||||
| Viburnum acerifolium | X | X | ||||||||||||
| Viburnum lantanoides | X | |||||||||||||
| Viburnum affine var. hypomalacum | X | |||||||||||||
| Viburnum prunifolium | X | X | ||||||||||||
| Viburnum rufidulum | X | |||||||||||||
| Vitis aestivalis | X | X | ||||||||||||
| Vitis rotundifolia | X | X | X | X | ||||||||||
| Vitis spp. | X | X | ||||||||||||
| a HMF = Hutcheson Memorial Forest [12]. Not listed in the table above and only found at this site: Viburnum dentatum, Lonicera maackii, Sambucus canadensis, Berberis thunbergii and Ligustrum vulgaris. DF = Duke Forest [13]. SG = Savage Gulf, mockernut hickory-northern red oak and chestnut oak community types [21]. SO = Scarlet Oak [27]. CO = Chestnut oak forest, Great Smoky Mountains National Park (NPS unpublished data). OH = Oak-hickory forest, Great Smoky Mountains National Park (NPS unpublished data). MW = Mauntel’s Woods [28]. HSNP = Hot Springs National Park, upland hardwoods and oak-hickory-pine types [30]. RBRNA = Roaring Branch Research Natural Area, mid and upper north slopes [31]. BS = Big Spring (unpublished data provided by S. Shifley, USDA-Forest Service, North Central Experiment Station, Columbia, MO. WW = Wegener Woods [35]. BC = Bob’s Creek stand, Kisatchie National Forest [36]. OSLS = Old Shortleaf Slope stand, Kisatchie National Forest [36]. Note: No Shrub Data available for HW, LCW, Ridge and Valley, S. ILL, Boston Mountains, RRSP, or EW. An X in the Table Represents the Presence of a Species within the Respective Site. | ||||||||||||||
Appendix C. Herbaceous Species Table
| Herb Species | Piedmont | Cumberland Plateau | Appalachian Mts. | Interior Low Plateau | Interior Highlands | Coastal Plain | |||||
| Central Eastern TN | Great Smoky Mountains NP | Southern IN | Ouachita Mts. | MO River Hills | Western Gulf | ||||||
| HMF a | DF | FCSP * | CO | OH | MW | HSNP | RBRNA | WW | BC | OSLS | |
| Ageratina altissium | X | ||||||||||
| Agrostis perennans | X | X | |||||||||
| Amphicarpaea bracteata | X | X | |||||||||
| Anemonella thalictroides | X | ||||||||||
| Antennaria plantaginifolia | X | X | |||||||||
| Arisaema dracontium | X | ||||||||||
| Arisaema spp. | X | ||||||||||
| Arisaema triphyllum | X | X | X | ||||||||
| Aureolaria laevigata | X | ||||||||||
| Aster spp. | X | ||||||||||
| Botrychium virginianum | X | X | X | ||||||||
| Brachyelytrum erectum | X | ||||||||||
| Carex leptonervia | X | ||||||||||
| Carex spp. | X | X | X | ||||||||
| Chasmanthium spp. | X | X | |||||||||
| Chimaphila maculata | X | X | |||||||||
| Circaca quadrisculcata | X | ||||||||||
| Claytonia virginica | X | X | |||||||||
| Clintonia umbellulata | X | ||||||||||
| Conopholis americana | X | X | |||||||||
| Desmodium glutinosum | X | ||||||||||
| Desmodium laevigatum | X | X | X | ||||||||
| Desmodium marilandicum | X | ||||||||||
| Desmodium nudiflorum | X | X | X | X | |||||||
| Desmodium rotundifolium | X | ||||||||||
| Desmodium spp. | X | ||||||||||
| Dicentra cucullaria | X | ||||||||||
| Dichanthelium dichotomum | X | ||||||||||
| Dioscorea villosa | X | ||||||||||
| Disporum lanuginosum | X | ||||||||||
| Dryopteris marginalis | X | ||||||||||
| Epigaea repens | X | ||||||||||
| Eupatorium dubium | X | ||||||||||
| Eupatorium steelei | X | ||||||||||
| Galium lanceolatum | X | ||||||||||
| Gillenia stipulata | X | ||||||||||
| Goodyera pubescens | X | X | |||||||||
| Heuchera americana | X | ||||||||||
| Heuchera pubescens | X | ||||||||||
| Hepatica americana | X | ||||||||||
| Hieracium gronovii | X | ||||||||||
| Hieracium venosum | X | ||||||||||
| Hieracium scabrum | X | ||||||||||
| Houstonia purpurea | X | ||||||||||
| Houstonia longifolia | X | ||||||||||
| Iris cristata | X | ||||||||||
| Lilium superbum | X | ||||||||||
| Liparis liliifolia | X | ||||||||||
| Luzula echinata | X | ||||||||||
| Maianthemum racemosa | X | ||||||||||
| Medeola virginiana | X | ||||||||||
| Melampyrum lineare | X | ||||||||||
| Mitchella repens | X | X | |||||||||
| Monarda clinopodia | X | ||||||||||
| Monotropa uniflora | X | ||||||||||
| Oenothera fruticosa | X | ||||||||||
| Panicum sphaerocarpoa | X | ||||||||||
| Panicum spp. | X | X | X | X | X | ||||||
| Phegopteris hexagonoptera | X | ||||||||||
| Poa alsodes | X | ||||||||||
| Polygonatum biflorum | X | ||||||||||
| Polystichum acrostichoides | X | X | X | X | |||||||
| Porteranthus trifoliatus | X | ||||||||||
| Potentilla canadensis | X | ||||||||||
| Prenanthes spp. | X | ||||||||||
| Pteridium aquilinum | X | ||||||||||
| Pycnanthemum spp. | X | ||||||||||
| Ranunculus spp. | X | ||||||||||
| Solidago curtisii | X | ||||||||||
| Solidago spp. | X | X | |||||||||
| Symphyotrichum anomalus | X | ||||||||||
| Symphyotrichum cordifolium | X | ||||||||||
| Symphyotrichum undulatum | X | ||||||||||
| Thalictrum thalictroides | X | ||||||||||
| Tradescantia subaspera | X | ||||||||||
| Uvularia puberula | X | ||||||||||
| Viola hastata | X | ||||||||||
| Viola spp. | X | X | |||||||||
| a HMF = Hutcheson Memorial Forest [12]. Not listed in the above table and only found at this site: Lonicera japonica V, Parthenocissus quinquefolia V., Smilacina racemosa, Pilea pumila, Impatiens biflora, Toxicodendron radicans V, Phytolacca americana, Vitis aestivalis V and 25 others listed as “other species” in [12]. DF = Duke Forest ([13] Oosting 1942). FCSP = Falls Creek State Park, Tennessee [92]. CO = Chestnut oak forest, Great Smoky Mountains National Park (NPS unpublished data). OH = Oak-hickory forest, Great Smoky Mountains National Park (NPS unpublished data). Note that this site had a total of 51 species, however we only list 38 of those in this table. MW = Mauntel’s Woods [28]. HSNP = Hot Springs National Park, upland hardwoods and oak-hickory-pine types [30]. RBRNA = Roaring Branch Research Natural Area, mid and upper north slopes [31]. WW = Wegener Woods [35]. BC = Bob’s Creek stand, Kisatchie National Forest [36]. OSLS = Old Shortleaf Slope stand, Kisatchie National Forest [36]. * Major species listed for Falls Creek State Park. For an extensive list see [93]. An X in the Table Represents the Presence of a Species within the Respective Site. | |||||||||||
Appendix D. Common Birds, Mammals, and Herpetofauna of Oak-Hickory Forests
References
- Oswalt, S.N.; Smith, W.B.; Miles, P.D.; Pugh, S.A. Forest Resources of the United States, 2012: A Technical Document Supporting the Forest Service 2015 Update of the RPA Assessment; General Technical Reports, WO-91; U.S. Department of Agriculture Forest Service, Washington Office: Washington, DC, USA, 2014; p. 218.
- Jenkins, M.A.; Webster, C.R.; Parker, G.R.; Spetich, M.A. Coarse woody debris in managed Central Hardwood Forests of Indiana, USA. For. Sci. 2004, 50, 781–792. [Google Scholar]
- Roleček, J. Vegetation types of dry-mesic oak forests in Slovakia. Preslia 2005, 77, 241–261. [Google Scholar]
- Bölöni, J.; Ódor, P.; Ádám, R.; Keeton, W.S.; Aszalós, R. Quantity and dynamics of dead wood in managed and unmanaged dry-mesic oak forests in the Hungarian Carpathians. For. Ecol. Manag. 2017, 399, 120–131. [Google Scholar] [CrossRef]
- White, D.L.; Lloyd, F.T. Defining old-growth: Implications for management. In General Technical Reports-SRS-1, Proceedings of the Eighth Biennial Southern Silvicultural Conference, Auburn, AL, USA, 1–3 November 1994; Edwards, M.B., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 1994; pp. 51–62. [Google Scholar]
- McNab, W.H.; Spetich, M.A.; Perry, R.; Clark, S. Chapter 11: Climate induced migration of native tree populations and consequences for forest composition. In Climate Change Adaptation and Mitigation Management Options: A Guide for Natural Resource Managers.; Vose, J.M., Klepzig, K., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 307–378. [Google Scholar]
- Allen, M.F. How oaks respond to water limitation. In Proceedings of the Seventh California Oak Symposium: Managing Oak Woodlands in a Dynamic World, Visalia, CA, USA, 3–6 November 2014; General Technical Reports PSW-GTR-251. Standiford, R.B., Purcell, K.L., Eds.; U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station: Berkeley, CA, USA, 2015; pp. 13–22. [Google Scholar]
- Eyre, F.H. (Ed.) Forest Cover Types of the United States and Canada; Society America Foresters: Washington, DC, USA, 1980; 148p. [Google Scholar]
- Braun, E.L. Deciduous Forests of Eastern North America; The Blakiston Co.: Philadelphia, PA, USA, 1950; 596p. [Google Scholar]
- Davison, S.E. Vegetational change in a mature oak forest: A thirty-year study. Master’s Thesis, Rutgers University, New Brunswick, NJ, USA, 1980; 93p. [Google Scholar]
- Smalley, G.W. Carya glabra (Mill.) Sweet Pignut Hickory. In Silvics of North America: 2. Hardwoods; Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990; Volume 2, pp. 198–204. 877p. [Google Scholar]
- Davison, S.E.; Forman, R.T.T. Herb and shrub dynamics in a mature oak forest: A thirty-year study. Bull. Torrey Bot. Club 1982, 109, 64–73. [Google Scholar] [CrossRef]
- Oosting, H.J. An ecological analysis of the plant communities of Piedmont, North Carolina. Am. Midl. Nat. 1942, 22, 333–350. [Google Scholar] [CrossRef]
- Fenneman, N.M. Physiography of Eastern United States; McGraw-Hill Book Co.: New York, NY, USA, 1938; 714p. [Google Scholar]
- Buol, S.W. (Ed.) Soils of the Southern United States and Puerto Rico; Southern Cooperative Series Bulletin No. 174; Agricultural Experiment Stations of the Southern United States and Puerto Rico Land-Grant Universities with Cooperative Assistance by USDA-Soil Conservation Service: Wenatchee, WA, USA, 1973; 105p.
- McCarthy, B.C.; Hammer, C.A.; Kauffman, G.L.; Cantino, P.D. Vegetation patterns and structure of an old-growth forest in southeast Ohio. Bull. Torrey Bot. Club 1987, 114, 33–45. [Google Scholar] [CrossRef]
- Martin, W.H. The Lilley Cornett Woods: A stable mixed mesophytic forest in Kentucky. Bot. Gazette 1975, 136, 171–183. [Google Scholar] [CrossRef]
- Galbraith, S.L.; Martin, W.H. Three decades of overstory and species change in a mixed mesophytic forest in eastern Kentucky. Castanea 2005, 70, 115–128. [Google Scholar] [CrossRef]
- Muller, R.N. Vegetation patterns in the mixed mesophytic forest of eastern Kentucky. Ecology 1982, 63, 1901–1917. [Google Scholar] [CrossRef]
- McEwan, R.W.; Muller, R.N. Spatial and temporal dynamics in canopy dominance of an old-growth central Appalachian forest. Can. J. For. Res. 2006, 36, 1536–1550. [Google Scholar] [CrossRef]
- Sherman, M.D. Community Composition, Species Diversity, Forest Structure and Dynamics as Affected by Soil and Site Factors and Selective Logging in Savage Gulf, Tennessee. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 1978; 146p. [Google Scholar]
- Nowacki, G.J.; Abrams, M.D. Community and edaphic analysis of mixed oak forests in the ridge and valley province of central Pennsylvania. In Proceedings of the Eighth Central Hardwood Conference, University Park, PA, USA, 4–6 March 1991; McCormick, L.H., Gottschalk, K.W., Eds.; General Technical Reports NE-148. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA; pp. 247–260. [Google Scholar]
- Sulser, J.S. Twenty years of change in the Hutcheson Memorial Forest. Wm. L. Hutcheson Meml. For. Bull. 1971, 3, 24–26. [Google Scholar]
- Bölöni, J.; Aszalós, R.; Frank, T.; Ódor, P. Forest type matters: Global review about the structure of oak dominated old-growth temperate forests. For. Ecol. Mgmt. 2021, 500, 119629. [Google Scholar] [CrossRef]
- Hart, J.L.; Clark, S.L.; Torreano, S.J.; Buchanan, M.L. Composition, structure, and dendroecology of an old-growth Quercus forest on the tablelands of the Cumberland Plateau, USA. For. Ecol. Mgmt. 2011, 266, 11–24. [Google Scholar] [CrossRef]
- Jenkins, M.A. Vegetation Communities of Great Smoky Mountains National Park. Southeast. Nat. 2007, 6, 35–56. [Google Scholar] [CrossRef]
- DeLapp, J.A.; Wentworth, T.R. Proposed Research Natural Areas in the Southern Appalachian Mountains; Unpublished Report; Highlands Biological Station: Highlands, NC, USA, 1977; 117p. [Google Scholar]
- Potzger, J.E.; Friesner, R.C. Some comparisons between virgin forest and adjacent areas of secondary succession. Butl. Univ. Bot. Stud. 1934, 3, 85–98. [Google Scholar]
- Fralish, J.S.; Crooks, F.B.; Chambers, J.L.; Harty, F.M. Comparison of Presettlement, Second-growth and Old-growth Forest on Six Site Types in the Illinois Shawnee Hills. Am. Midl. Nat. 1991, 125, 294–309. [Google Scholar] [CrossRef]
- Dale, E.E.; Watts, M.R. Vegetation of Hot Springs National Park, Arkansas; Final Report for the Southwest Region; NPS, U.S. Department Interior (#CX-70299001); U.S. Department of Interior, National Park Service: Washington, DC, USA, 1980; p. 85.
- Fountain, M.S.; Sweeney, J.M. Ecological Assessment of the Roaring Branch Research Natural Area; Research Paper SO-213; U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station: Asheville, NC, USA, 1985; p. 15.
- Stahle, D.W.; Hehr, J.G.; Hawks, G.G., Jr. The Development of Modern Tree-Ring Chronologies in the Midwest: Arkansas, North Texas, Oklahoma, Kansas, and Missouri; Final Report to Climate Dynamics Program; Arkansas Archeological Survey and Department of Geography, University of Arkansas: Fayetteville, AR, USA; NSF: Washington, DC, USA, 1982.
- Shifley, S.R.; Thompson, F.R., III; Schlesinger, R.C.; Ponder, F., Jr.; Parker, G.R.; Spetich, M.A. Study Plan: Composition of Old-Growth Hardwood Forests in the Midwest; Study FS-NC-4154(91-04), Research Work Unit FS-NC-4154; USDA Forest Service North Central Forest Experiment Station: Columbia, MO, USA, 1991.
- Lowney, C.A. Two Decades of Change in the Structure and Composition of Old-Growth Forests in the Central Hardwood Region, U.S.A. Master’s Thesis, Purdue University, West Lafayette, IN, USA, 2013; p. 156. [Google Scholar]
- Wuenscher, J.E. A Vegetational Analysis of a Virgin Hardwood Stand in East-Central Missouri. Master’s Thesis, University of Missouri, Columbia, MO, USA, 1967; p. 74. [Google Scholar]
- Martin, D.L.; Smith, L.M. A Survey and Description of the Natural Plant Communities of the Kisatchie Forest Winn and Kisatchie Districts; Louisiana Department of Wildlife and Fisheries: Baton Rouge, LA, USA, 1991; p. 372.
- Marks, P.L.; Harcombe, P.A. Forest vegetation of the big thicket, southeast Texas. Ecol. Monogr. 1981, 5, 287–305. [Google Scholar] [CrossRef]
- Greenberg, C.H.; McLeod, D.E.; Loftis, D.L. An Old-Growth Definition for Western and Mixed Mesophytic Forests; Technical Report SRS-16; USDA Forest Service General Technical Reports: Asheville, NC, USA, 1997.
- Ferguson, E.R. Establishment Report—Roaring Branch Research Natural Area; Unpublished Manuscript; Southern Forest Experiment Station: New Orleans, LA, USA, 1968; p. 7. [Google Scholar]
- Stahle, D.W.; Hehr, J.G.; Hawks, C.G., Jr.; Cleaveland, M.K.; Baldwin, J.R. Tree Ring Chronologies for the Southcentral United States; Tree-Ring Lab., University Arkansas: Fayetteville, AR, USA, 1985; 135p. [Google Scholar]
- Shifley, S.R.; Roovers, L.M.; Brookshire, B.L. Structural and compositional differences between old-growth and mature second-growth forests in the Missouri Ozarks. In Proceedings of the 10th Central Hardwood Forest Conference, Radnor, PA, USA, 5–8 March 1995; USDA Forest Service General Technical Reports NE-197. U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1995; pp. 23–36. [Google Scholar]
- Thom, R.H.; Iffrig, G. Directory of Missouri Natural Areas; Missouri Natural Areas Committee, Missouri Department of Conservation and Missouri Department of Natural Resources: Jefferson City, MO, USA, 1985; p. 115.
- Nigh, T.; Putman, C.; Moore, D. Draft Missouri Natural Area Nomination: Big Spring Pines; Missouri Department of Conservation: Jefferson City, MO, USA, 1992; p. 12.
- White, D.L.; Lloyd, F.T. Old-Growth Definition for Dry and Dry-Mesic Oak-Pine Forests; General Technical Report SRS-23; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 1998; pp. 1–43.
- Parker, G.R. Old-growth forests of the Central Hardwood Region. Nat. Areas J. 1989, 19, 5–11. [Google Scholar]
- Burns, R.M.; Honkala, B.H. Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; USDA Forest Service: Washington, DC, USA, 1990; Volume 2, p. 877.
- Martin, W.H. Characteristics of old growth mixed-mesophytic forests. Nat. Areas J. 1992, 12, 127–135. [Google Scholar]
- Monk, C.D. The vegetation of the William L. Hutcheson Memorial Forest, New Jersey. Bull. Torrey Bot. Club 1961, 88, 156–166. [Google Scholar] [CrossRef]
- Blozan, W. The importance of increment core samples and disturbance history in the evaluation of old-growth forests in Great Smoky Mountains National Park. Nat. Areas J. 1994, 10, 19–27. [Google Scholar]
- Batista, W.B.; Platt, W.J. An Old-Growth Definition for Southern Mixed Hardwood Forests; General Technical Report SRS-9; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 1997; p. 15.
- Murphy, P.A.; Nowacki, G.J. An Old-Growth Definition for Xeric Pine and Pine-Oak Woodlands; General Technical Report SRS-7; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 1997; p. 7.
- Monk, C.D. Past and present influences on reproduction in the William L. Hutcheson Memorial Forest, New Jersey. Bull. Torrey Bot. Club 1961, 88, 167–175. [Google Scholar] [CrossRef]
- Frei, K.R.; Fairbrothers, D.E. Floristic study of the William L. Hutcheson Memorial Forest (New Jersey). Bull. Torrey Bot. Club 1963, 90, 338–353. [Google Scholar] [CrossRef]
- Carlson, P.J. An Assessment of the Old-Growth Forest Resource on National Forest System Lands in the Chattooga River Watershed; Chattooga Ecosystem Demonstration Management Project; USDA Forest Service-Region 8: Franklin, NC, USA, 1995; 79p.
- Adjemovitch, M. A study of seven alien species in the William L. Hutcheson Memorial Forest. Master’s Thesis, Rutgers—The State University, Piscataway, NJ, USA, 1958. [Google Scholar]
- Reiners, N.M.; Reiners, W.A. Natural harvesting of trees. William Hutcheson Memorial For. Bull. 1965, 2, 9–17. [Google Scholar]
- McComb, W.C.; Muller, R.N. Snag densities in old-growth and second growth Appalachian Forests. J. Wildl. Manag. 1983, 47, 376–382. [Google Scholar] [CrossRef]
- Shifley, S.R.; Brookshire, B.L.; Larsen, D.R.; Herbeck, L.A. Snags and down wood in Missouri old-growth and mature second-growth forests. North. J. Appl. For. 1997, 14, 165–172. [Google Scholar] [CrossRef]
- Spetich, M.A.; Shifley, S.R.; Parker, G.R. Regional distribution and dynamics of coarse woody debris in Midwestern old-growth forests. For. Sci. 1999, 45, 302–313. [Google Scholar]
- Walker, L.C. The southern pine region. In Regional Silviculture of the United States; Barrett, J.W., Ed.; Wiley: New York, NY, USA, 1980; pp. 231–276. 551p. [Google Scholar]
- MacMillan, P.C. Decomposition of coarse woody debris in an old-growth Indiana forest. Can. J. For. Res. 1988, 18, 1353–1362. [Google Scholar] [CrossRef]
- Rosenberg, D.K.; Fraser, J.D.; Stauffer, D.F. Use and characteristics of snags in young and old forest stands in southwest Virginia. For. Sci. 1988, 34, 224–228. [Google Scholar]
- Johnson, F.L.; Schnell, G.D. Wildland Fire History and the Effects of Fire on Vegetative Communities at Hot Springs National Park, Arkansas; (contract #CX-702930034); National Park Service: Santa Fe, NM, USA, 1985.
- Lang, G.E.; Forman, R.T.T. Detrital dynamics in a mature oak forest: Hutcheson Memorial Forest, New Jersey. Ecology 1978, 59, 580–595. [Google Scholar] [CrossRef]
- Muller, R.N.; Liu, Y. Coarse woody debris in an old-growth deciduous forest on the Cumberland Plateau, southeastern Kentucky. Can. J. For. Res. 1991, 21, 1567–1572. [Google Scholar] [CrossRef]
- Monk, C.D. Plant communities of Hutcheson Memorial Forest based on shrub distribution. Bull. Torrey Bot. Club 1957, 88, 156–166. [Google Scholar] [CrossRef]
- Spetich, M.A.; Dey, D.C.; Johnson, P.S.; Graney, D.L. Competitive capacity of Quercus rubra L. planted in Arkansas’ Boston Mountains. For. Sci. 2002, 48, 504–517. [Google Scholar]
- Oak, S.W.; Spetich, M.A.; Morin, R.S. Chapter 3: Oak decline in central hardwood forests: Frequency, spatial extent, and scale. In Natural Disturbances and Historic Range of Variation: Type, Frequency, Severity, and Post-Disturbance Structure in Central Hardwood Forests USA; Book Series Managing Forest Ecosystems; Greenberg, C.H., Collins, B.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 32, 400p. [Google Scholar]
- White, P.S.; White, R.D. Old-growth oak and oak-hickory forests. In Eastern Old-Growth Forests: Prospects for Rediscovery and Recovery; Davis, M.B., Ed.; Island Press: Washington, DC, USA, 1996; pp. 383, 178–198. [Google Scholar]
- Hawthorne, S.; Miniat, C.F. Topography may mitigate drought effects on vegetation along a hillslope gradient. Ecohydrology 2017, 11, e1825. [Google Scholar] [CrossRef]
- Sander, I.L.; Clark, F.B. Reproduction of Upland Hardwood Forests in the Central States; Handbook 405; USDA Foreign Agricultural Service: Washington, DC, USA, 1971; p. 25.
- Johnson, P.S.; Shifley, S.R.; Rogers, R.; Dey, D.C.; Kabrick, J.M. The Ecology and Silviculture of Oaks, 3rd ed.; CABI Publishing: Wallingford Oxon, UK, 2019; p. 628. [Google Scholar]
- Abrams, M.D. Fire and the development of oak forests. Bioscience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Keyser, T.L. Resprouting by seedlings of four North American deciduous broadleaved tree species following experimental burning. Oecologia 2019, 190, 207–218. [Google Scholar] [CrossRef]
- Van Lear, D.H. Fire and oak regeneration in the southern Appalachians. In Fire and the Environment: Ecological and Cultural Perspectives, Proceedings of the Internatioal Symposium, Knoxville, Tennese, 20–24 March 1990; General Technical Report SE-69; USDA Forest Service, Southern Forest Experiment Station: Asheville, NC, USA, 1991; pp. 15–21. [Google Scholar]
- Larson, D.R.; Johnson, P.S. Linking the ecology of natural oak regeneration to silviculture. For. Ecol. Manag. 1998, 106, 1–7. [Google Scholar] [CrossRef]
- Oliver, C.D. The development of northern red oak in mixed stands in central New England. Yale Univ. Sch. For. Environ. Stud. Bull. 1978, 91, 63. [Google Scholar]
- Nowacki, G.J.; Adams, M.D.; Lorimer, C.G. Composition, structure, and historical development of northern red oak stands along an edaphic gradient in north-central Wisconsin. For. Sci. 1990, 36, 276–292. [Google Scholar] [CrossRef]
- Lafon, C.W.; Naito, A.T.; Grissino-Mayer, H.D.; Horn, S.P.; Waldrop, T.A. Fire history of the Appalachian Region: A Review and Synthesis; General Technical Report SRS-219; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2017; p. 97.
- Guyette, R.P.; Dey, D.C.; Stambaugh, M.C.; Muzika, R.-M. Fire scars reveal variability and dynamics of eastern fire regimes. In Proceedings of the Fire in Eastern Oak Forests: Delivering Science to Land Managers, Columbus, OH, USA, 15–17 November 2005; Matthew, B.D., Ed.; General Technical Report NRS-P-1. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2006; pp. 20–39. [Google Scholar]
- Iffrig, G.F.; Trammel, C.E.; Cunningham, T. Pioneer Forest: A case study in sustainable forest management. In Toward Sustainability for Missouri Forests; Flader, S.L., Ed.; General Technical Report NC-239; Department of Agriculture, Forest Service, North Central Research Station: St. Paul, MN, USA, 2004; pp. 193–204. [Google Scholar]
- Woods, F.W.; Shanks, R.E. Replacement of chestnut in the Great Smoky Mountains of Tennessee and North Carolina. J. For. 1957, 55, 847. [Google Scholar]
- Blackburn, L.M.; Hajek, A.E. Gypsy Moth Larval Necropsy Guide; General Technical Report NRS-179; Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2018; p. 30. [CrossRef]
- Clinton, B.D.; Boring, L.R.; Swank, W.T. Canopy gap characteristics and drought influences in oak forests of the Coweeta Basin. Ecology 1993, 74, 1551–1558. [Google Scholar] [CrossRef]
- Jenkins, M.A.; Pallardy, S.G. The influence of drought on red oak group species growth and mortality in the Missouri Ozarks. Can. J. For. Res. 1995, 25, 1119–1127. [Google Scholar] [CrossRef]
- Fan, Z.; Fan, X.; Crosby, M.K.; Moser, W.K.; He, H.; Spetich, M.A.; Shifley, S.R. Spatio-temporal trends of oak decline and mortality under periodic regional drought in the Ozark Highlands of Arkansas and Missouri. Forests 2012, 3, 614–631. [Google Scholar] [CrossRef]
- Spetich, M.A.; Fan, Z.; Sui, Z.; Crosby, M.K.; He, H.S.; Shifley, S.R.; Leininger, T.D.; Moser, W.K. Tree mortality estimates and species distribution probabilities in southeastern United States forests. In Forest Health Monitoring: National Status, Trends, and Analysis; Kevin, M.P., Barbara, L.C., Eds.; General Technical Report SRS-222; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2017; pp. 175–181. Available online: https://www.srs.fs.fed.us/pubs/54586.
- Runkle, J.R. Gap dynamics in an Ohio Acer-Fagus forest and speculations on the geography of disturbance. Can. J. For. Res. 1990, 20, 632–641. [Google Scholar] [CrossRef]
- Spetich, M.A. Survival of Quercus alba (White Oak) advance reproduction in small group and single tree openings. Forests 2020, 11, 889. [Google Scholar] [CrossRef]
- Spetich, M.A.; Parker, G.R. Distribution of Biomass in an Indiana Old-growth Forest from 1926 to 1992. Am. Midl. Nat. 1998, 139, 90–107. [Google Scholar] [CrossRef]
- Cain, S.A. Studies on Virgin Hardwood Forest: III. Warren’s Woods, A Beech-Maple Climax Forest in Berrien County, Michigan. Ecology 1935, 16, 500–513. [Google Scholar] [CrossRef]
- Caplenor, D. The vegetation of the gorges of the Fall Creek Falls State Park in Tennessee. J. Tenn. Acad. Sci. 1965, 40, 27–39. [Google Scholar]
- Fleming, C.A.; Wofford, B.E. The Vascular Flora of Fall Creek Falls State Park, Van Buren and Bledsoe Counties, Tennessee. Castanea 2004, 69, 164–184. [Google Scholar] [CrossRef]
- Dickson, J.G.; Conner, R.N.; Williamson, J.H. Relative Abundance of Breeding Birds in Forest Stands in the Southeast. South. J. Appl. For. 1980, 4, 174–179. [Google Scholar] [CrossRef]
- Webster, J.D.; Adams, D.L. Breeding bird censuses in old-growth deciduous forests. Proc. Indiana Acad. Sci. 1973, 82, 198–206. [Google Scholar]
- Bryant, W.S.; McComb, W.C.; Fralish, J.S. Oak-hickory forests (western mesophytic/oak-hickory forests). In Biodiversity of the Southeastern United States: Upland Terrestrial Communities; Martin, W.H., Boyce, S.G., Echternacht, A.C., Eds.; Wiley: New York, NY, USA, 1993; pp. 143–201. 373p. [Google Scholar]
- Pelton, M.R. The importance of old growth to carnivores in eastern deciduous forests. In Eastern Old-Growth Forests: Prospects for Rediscovery and Recovery; Davis, M.B., Ed.; Island Press: Washington, DC, USA, 1996; pp. 65–75. 383p. [Google Scholar]
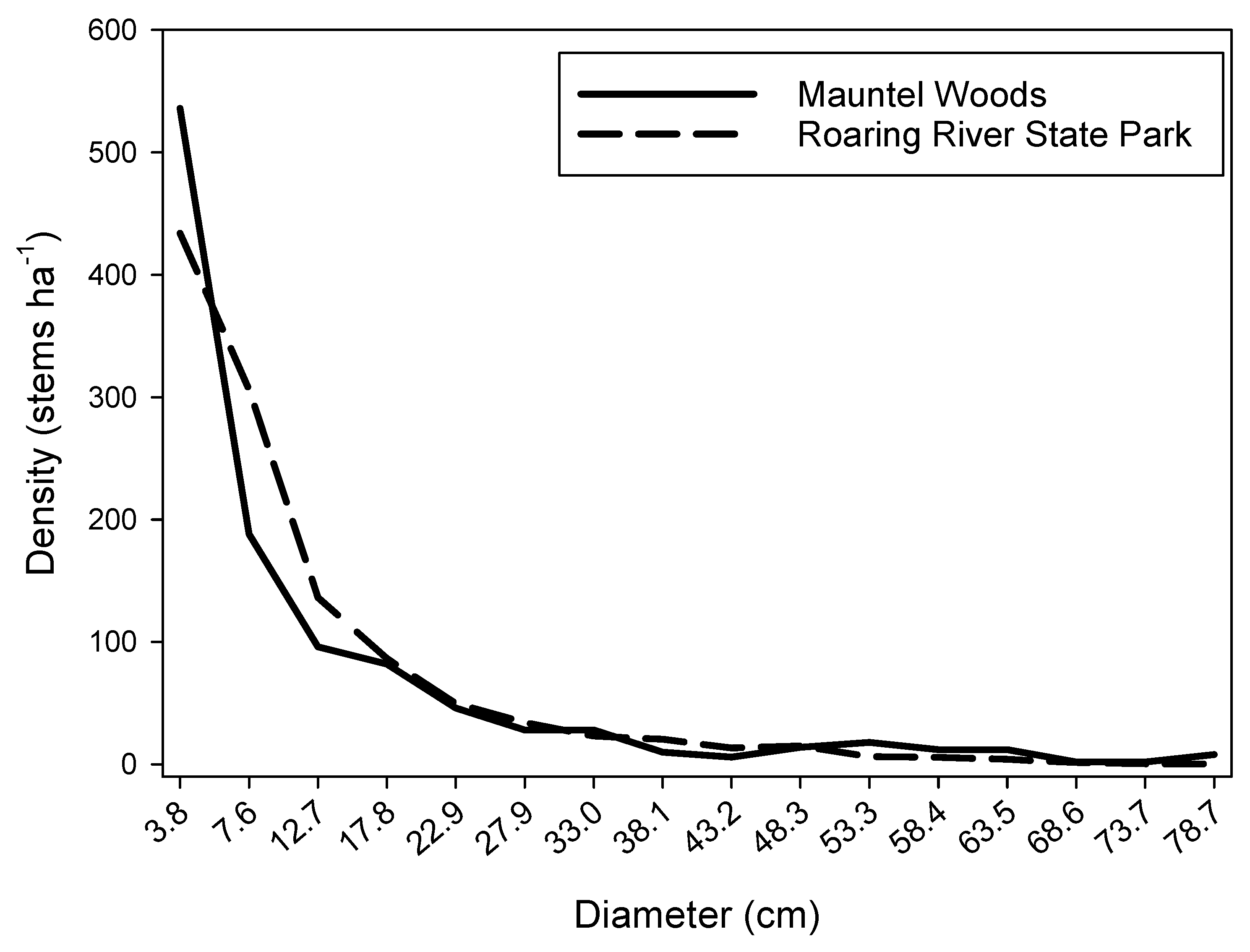
| Quantifiable Attribute | Value Range | Mean * | No. of Stands | References |
|---|---|---|---|---|
| TREES (# ha−1) | ||||
| Interior Highlands | ||||
| Springfield Plateau | ||||
| Roaring River St. Park | ||||
| N and E Slopes (≥10 cm dbh) | 0–20 | 4 | 1 | [33] (1992 Data) |
| Ridge + S & W Slopes (≥10 cm dbh) | 0–40 | 8 | 1 | |
| Salem Plateau | ||||
| Big Spring Natural Area (≥10 cm dbh) | 0–30 | 8 | 1 | [33] (1992 Data) |
| Missouri River Hills | ||||
| Englemann Woods | ||||
| Ridge + S & W slopes (≥10 cm dbh; #ha−1) | 10–20 | 13 | 1 | [33] (1992 Data) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spetich, M.A.; Jenkins, M.A.; Shifley, S.R.; Wittwer, R.F.; Graney, D.L. Characteristics of Dry-Mesic Old-Growth Oak Forests in the Eastern United States. Earth 2022, 3, 975-1009. https://doi.org/10.3390/earth3030057
Spetich MA, Jenkins MA, Shifley SR, Wittwer RF, Graney DL. Characteristics of Dry-Mesic Old-Growth Oak Forests in the Eastern United States. Earth. 2022; 3(3):975-1009. https://doi.org/10.3390/earth3030057
Chicago/Turabian StyleSpetich, Martin A., Michael A. Jenkins, Stephen R. Shifley, Robert F. Wittwer, and David L. Graney. 2022. "Characteristics of Dry-Mesic Old-Growth Oak Forests in the Eastern United States" Earth 3, no. 3: 975-1009. https://doi.org/10.3390/earth3030057
APA StyleSpetich, M. A., Jenkins, M. A., Shifley, S. R., Wittwer, R. F., & Graney, D. L. (2022). Characteristics of Dry-Mesic Old-Growth Oak Forests in the Eastern United States. Earth, 3(3), 975-1009. https://doi.org/10.3390/earth3030057